
A. Haghighi
Department of Parasitology and Mycology, School of Medicine, Shahid Beheshti University (Medical Campus), Tehran, Iran
S. Rasti
Department of Parasitology and Mycology, Kashan University of Medical Sciences and Health Services, Kashan, Iran
E. Nazemalhosseini Mojarad
Research Center for Gastroenterology and Liver Diseases, Shahid Beheshti University, MC, Tehran, Iran
B. Kazemi
Department of Parasitology and Mycology, School of Medicine, Shahid Beheshti University (Medical Campus), Tehran, Iran
M. Bandehpour
Cellular and Molecular Biology Research Centers, Shahid Beheshti University MC, Iran
Z. Nochi
Research Center for Gastroenterology and Liver Diseases, Shahid Beheshti University, MC, Tehran, Iran
H. Hooshyar
Department of Parasitology and Mycology, Kashan University of Medical Sciences and Health Services, Kashan, Iran
M. Rezaian
Department of Parasitology and Mycology, School of Public Health, Tehran University of Medical Sciences and Health Services, Tehran, Iran
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 23 | Page No.: 2613-2618







ABSTRACT
The nucleotide sequences of Serine-Rich Entamoeba histolytica Protein (SREHP) gene have already exhibited stable and significant polymorphism in the gene studies. Serine-rich protein is also present and polymorphic in Entamoeba dispar which called SREDP. The polymorphism of the Serine-Rich Entamoeba dispar Protein (SREDP) gene among 8 isolates obtained from Iranian cyst carriers were analyzed by a nested PCR-RFLP followed by sequencing of the PCR products. From those isolates, six distinct DNA patterns were observed after PCR-RFLP of the nested PCR, whereas sequencing showed 8 different patterns among the isolates. The results demonstrate an extensive genetic variability among Iranian E. dispar isolates. The repeat-containing region of the SREDP was found extensively polymorphic in size, number and order of repeat units. Genetic diversity of Iranian E. dispar isolates based on the SREDP was more polymorphic in comparison of Serine-Rich Entamoeba histolytica Protein (SREHP) of the E. histolytica isolates as well as were different from a few known SREDP genes.
PDF Abstract XML References Citation
How to cite this article
A. Haghighi, S. Rasti, E. Nazemalhosseini Mojarad, B. Kazemi, M. Bandehpour, Z. Nochi, H. Hooshyar and M. Rezaian, 2008. Entamoeba dispar: Genetic Diversity of Iranian Isolates Based on Serine-Rich Entamoeba dispar Protein Gene. Pakistan Journal of Biological Sciences, 11: 2613-2618.
DOI: 10.3923/pjbs.2008.2613.2618
URL: https://scialert.net/abstract/?doi=pjbs.2008.2613.2618
DOI: 10.3923/pjbs.2008.2613.2618
URL: https://scialert.net/abstract/?doi=pjbs.2008.2613.2618
INTRODUCTION
Based on morphology and structure, Entamoeba dispar is very similar to Entamoeba histolytica (WHO/PAHO/UNESCO, 1997). Although, there is some evidence that E. dispar is able to produce intestinal lesions in animals (Espinosa Cantellano et al., 1997) and epithelial cell monolayer in vitro (Espinosa- Cantellano et al., 1998), the general agreements say that E. dispar is a non-pathogenic parasite. These realities suggest that the non-pathogenicity of some strains of this species is doubtful and more investigations are required (Pinheiro et al., 2005).
The first nucleotide sequences have been shown the stable and significant polymorphism in the Strain Specific Gene (SSG) and the Serine-Rich E. histolytica Protein (SREHP) gene. SSG is absent from E. dispar and all other species as far as is known (Clark, 2006) but SREHP is also present and polymorphic in E. dispar and called SREDP (Mai and Samuelson, 1998). Ayeh-Kumi et al. (2001) studied clinical samples from Bangladesh; they used a nested PCR of the SREHP gene coupled with restriction digestion. Twenty five genotypes have been found among 42 intestinal isolates and 9 genotypes among the isolates from 12 ALA samples. In Clark and Diamond (1993) study, 16 different genotypes were observed among 18 isolates of E. histolytica from diverse geographical locations using the combined results of restriction digestion of SREHP and amplification of the SSG locus. Ghosh reported 5 different SREDP sequencing patterns from 5 isolates and showed high genetic variation among the E. dispar in compare with E. histolytica (Ghosh et al., 2000). Haghighi et al. (2003) investigated 79 isolates of E. histolytica, mostly from Japan and Thailand by sequencing of four loci (two tRNA-linked STR loci, Chitinase and SREHP); however, it was failed to find an association between the parasite genotype and the outcome of infection. Cloning and characterization of SREHP gene from an Iranian E. histolytica isolate was also studied (Rasti et al., 2006), but no genetic variation was reported.
In the present study, the genetic variability of E. dispar strains based on the SREDP gene locus was analyzed using RFLP-nested PCR and sequencing of the obtained genes.
MATERIALS AND METHODS
Stool specimens: Eight E. dispar strains were previously isolated from three different climatic regions including central, northern and southern regions of Iran (Hooshyar et al., 2004; Kobayashi et al., 2005), which were analyzed in this study from June 2005 to May 2006. The Entamoeba strains were originally detected microscopically and differentiated from E. histolytica by PCR. The cultivated isolates in Robinson medium were transferred to Japan and maintained in liquid nitrogen in National Institute of Infection Diseases (NIID), Tokyo, Japan.
Extraction of genomic DNA and PCR identification: The DNA was extracted for PCR amplification in this study after recovery and re-cultivation of the frozen isolates in Robinson medium (Robinson, 1968). Total genomic DNA from trophozoites was purified using QIAamp DNA stool mini-kit in NIID (QIAGEN, Tokyo, Japan) according to the manufacturer’s directions. The purified DNA was transferred to Iran and maintained in -20°C until used.
To confirm the presence of E. dispar and for exclusion of E. histolytica from study, PCR with two sets of species-specific primers from locus D-A gene was performed (Hsp1 and Hsp2 for E. histolytica and Dsp1 and Dsp2 for E. dispar) (Zaki and Clark, 2001; Zaki et al., 2002; Ali et al., 2005). PCR amplification consisted of 35 cycles of 45 sec at 93°C, 30 sec at the primer-dependent annealing temperature and 60 sec at 72°C with a final extension of 5 min at 72°C. The PCR products were electrophoresed in 1.2% agarose gel (Fermentas, No. R0491) containing ethidium bromide (Sambrook and Russell, 2001), was photographed under UV light (UVI doc Deluxe GAS 9000, England).
Nested SREDP gene PCR amplification: A set of primers [SREHP-BamH1 (S) (5` GAGGATCCATGTTC GCA TTTTTATTGT 3`) and SRD-Bam (AS) (5`GAGGATCCTT AGAAGACAATTGCCA3`)] which were designed based on SREDP gene (Ghosh et al., 2000) were used for initial PCR, followed by a second set of primers [Bam-SRED (S) (5`TTTATTGGATCCACTACT GCAACTA3`) and Bam-SRED (AS) (5`AAAGCAGGATCCA TAATGAATGG AC3`)].
External PCR was carried out in a 50 μL reaction mixture containing 200 ng of DNA, 2 μM of each external primer, 1.5 mM MgCl2, 0.2 mM of dNTP and 0.2 μL of Taq DNA polymerase (5 units μL-1) with the following cycling parameters: (1) pre-denaturation at 94°C for 5 min (2) 35 cycles of denaturing at 93°C for 45 sec, annealing at 60°C for 30 sec and extension at 72°C for 45 sec and (3) final extension at 72°C for 10 min.
Internal PCR carried out in a 50 μL reaction mixture containing 1 μL of the first PCR product, 2 μM of each internal primer, 1.5 mM MgCl2, 0.2 mM of dNTP and 0.3 μL of Taq DNA polymerase (5 units μL-1) in the same external cycling parameters with the exception of annealing at 64°C for 30 sec. The amplified fragment of SREDP gene was almost 680 bp. PCR product was purified by Q-Biogen Gene clean II Kit (BIO101, Lajolla, Calif).
AluI digestion of the nested SREDP PCR products: The PCR products were additionally analyzed by digestion with the restriction endonuclease AluI (Gibco BRL, Life Technologies). Seven microliter of SREDP PCR product was digested for 2 h at 37°C according to the manufacturer’s recommended procedure. Digested PCR products were electrophoresed in 2% agarose gel, in TBE buffer, ethidium bromide solution 10 mg mL-1, at 100 V for 1 h. The gel was visualized under UV light and photographed (Sambrook and Russell, 2001).
Sequence analysis: SREDP PCR products were directly sequenced using an Applied Biosystems (ABI) Terminator Cycle Sequencing Ready Reaction kit (BigDye ® Terminator V3.1 Cycle Sequencing Kit) on an ABI Analyzer. The obtained sequences were manually edited and aligned using Lasergene software (version 6.00).
RESULTS AND DISCUSSION
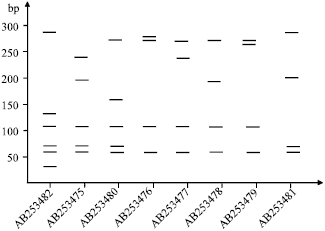
Eight E. dispar isolates were analyzed in this study. The polymerase chain reaction using Dsp1+Dsp2 reconfirmed that all the samples were E. dispar. The nested SREDP PCR fragments from the 8 Iranian strains showed slightly polymorphism of size among the isolates (Fig. 1). In the other hand to better understand the nature of the polymorphism observed by gel electrophoresis, the resultant nucleotide sequences of SREDP from all 8 isolates were used to investigate genetic diversity of E. dispar isolates. Sequencing of SREDP, showed 8 new genotypes with a highly polymorphic repeat-containing region of 20 to 24 distinct repeat units (Fig. 2). Sequencing revealed the appearance of the characteristic 648-720 bp band in 8 isolates of E. dispar, in which 684 bp fragments were found in 4 (50%) of the samples. The repeat-containing region of SREDP was found to be extensively polymorphic in size, species, number and order to repeat units among the isolates. Figure 2 shows sequencing analysis of the SREDP gene from HHR 1 IR strain (accession No. AB253476) and schematic diagram for genetic diversity of the gene repeat units. Nucleotide sequences of the E. dispar isolates were submitted to the GenBank/DDBJ/EMBL and are available for public access under the accession No. AB253475-AB253482. We also verified the AluI RFLP analysis of SREDP from all 8 isolates. Six distinct DNA patterns were observed after AluI digestion of nested PCR products. We conducted a computational RFLP analysis by AluI digestion of the SREDP locus based on the nucleotide sequences of the SREDP loci (Fig. 3). Although, all 8 genotypes showed distinct computational RFLP pattern, but the isolates with database accession number AB253476 and AB253479 as well as AB253478 and AB253481 showed very close patterns not possible to differentiate in agarose gel electrophoresis.
 | |
| Fig. 1: | Agarose gel electrophoresis of locus SREDP from Iranian E. dispar isolates. Lane 1 to 8: PCR fragment of the 8 isolates. Number of the lanes is in order to size of the sequencing results in Fig. 2, Lane DW: Negative control with distilled water. Lane M: 100 bp DNA ladder |
Studies show that E. dispar is perhaps 10 times more prevalent than E. histolytica worldwide, but local prevalence may vary meaningfully (Ali et al., 2005). Serine-Rich E. histolytica Protein (SREHP) gene is a surface protein also present and polymorphic in E. dispar and called SREDP (Mai and Samuelson, 1998). Although in serine-rich E. histolytica protein gene, polymorphism in lengths, types and numbers of internal repeat units were previously reported by Clark and Diamond (1993), Ghosh et al. (2000) and Haghighi et al. (2002, 2003), but genetic variation was studied just in 5 isolates in the SREDP gene (Ghosh et al., 2000). The present study is an initial experience of characterizing and sequencing of SREDP gene among Iranian E. dispar isolates in Iran. SREDP gene sequences contain conserved regions and internal tandem dodecapeptides and octapeptides repeat units like SREHP gene sequences (Haghighi et al., 2002). However, the patterns and sizes of amplified fragments corresponding to SREDP in E. dispar showed highly variable completely different from SREHP in E. histolytica strains (Rasti et al., 2006; Haghighi et al., 2002, 2003). A significant polymorphism among E. histolytica isolates collected from a wide geographic range was shown in previous study (Ghosh et al., 2000; Clark and Diamond, 1993).
PCR-RFLP of SREDP gene indicated 6 different patterns of 8 E. dispar strains which are emphasized by nucleotide sequencing of 8 different patterns. Recently in Bangladesh and Georgia, an extensive genetic diversity of E. histolytica was also observed by analysis of the SREHP gene by PCR amplification and AluI digestion (Simonishvili et al., 2005; Ayeh-Kumi et al., 2001). The serine-rich gene of E. histolytica was investigated by nested PCR amplification and restriction enzyme fragment length polymorphism in 26 clinical isolates of Turkey. After AluI digestion of nested PCR products, 12 different DNA patterns were observed which was demonstrated a comprehensive genetic variability of Turkish E. histolytica clinical isolates (Tanyuksel et al., 2008 ). In another study, 13 different profiles were generated from 23 E. histolytica isolates from Cameroon, Zimbabwe and South Africa, while 20 others were generated from 38 E. histolytica PCR positive stool samples from South Africa with RFLP-PCR (Samie et al., 2008).
Haghighi et al. (2003) found the most noteworthy and extensive variations among the SREHP locus. They demonstrated that E. histolytica had a highly complex genetic structure independent of geographic location, but failed to find an association between the parasite genotype and the outcome of infection.
Ayeh-Kumi et al. (2001) examined the combinations of the nested PCR results and the AluI digestion of the PCR products. That examination yielded 25 distinct DNA banding patterns among the 42 stool isolates and an additional 9 distinct patterns among the 12 liver abscess isolates (Ayeh-Kumi et al., 2001). The results here demonstrate an extensive genetic variability among SREDP gene of 8 Iranian E. dispar isolates. From those isolates six distinct DNA patterns were observed after PCR-RFLP of the nested PCR, whereas sequencing showed 8 different patterns among the isolates with a highly polymorphic repeat-containing region of 20 to 24 distinct repeat units. Genetic diversity of Iranian E. dispar isolates based on the SREDP gene showed 8 novel E. dispar genotypes which were distinct from only 5 known SREDP genes (Ghosh et al., 2000).
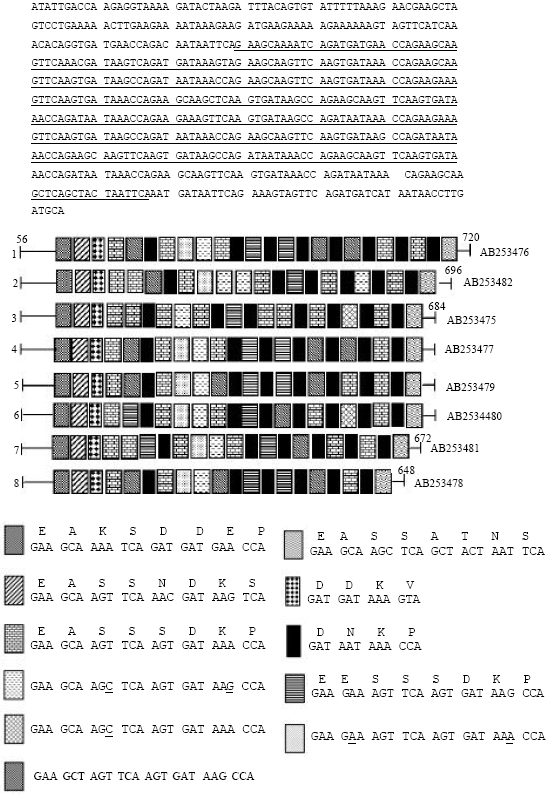 | |
| Fig. 2: | Sequence and schematic representation of the polymorphisms in novel genotypes of SREDP in 8 Iranian E. dispar isolates. Sequencing and the underline repeat-containing region of SREDP in the sequence derived from sample HHR 1 IR (Accession No. AB253476). The sequence No. 3 (accession No. AB253475) was used as a standard strain of E. dispar (Kobayashi et al ., 2005). The sequences of each of the 8 variants of the 24 and 12 nucleotide repeats are shown with a corresponding shaded block. The conserved non-repeated regions are shown as a single line. Genotypes of the 8 isolates and their related accession numbers are shown to the left and right of the schematics, respectively. The nucleotide and deduced amino acid sequences of tetra- and octapeptide repeats are shown above |
 | |
| Fig. 3: | Schematic profile of AluI digests of the 8 novel genotypes. The isolates AB253476 and AB253479 as well as AB253478 and AB25381 showed very close patterns. Note that the individual genotype is designated for each DNA fragment and the database accession number of the isolates |
ACKNOWLEDGMENTS
The authors would like to express special thanks to Shohreh Farnia from Tehran University of Medical Sciences and Health Services, School of Public Health, for her kind assistance. The research project was fully sponsored by Cellular and Molecular Biology Research Centers, Shahid Beheshti University M.C., Iran, with grant number 6371.
REFERENCES
- Ali, I.K.M., M. Zaki and C.G. Clark, 2005. Use of PCR amplification of tRNA gene-linked short tandem repeats for genotyping Entamoeba histolytica. J. Clin. Microbiol., 43: 5842-5847.
CrossRefDirect Link - Ayeh-Kumi, P.F., I.M. Ali, L.A. Lockhart, C.A. Gilchrist, W.A. Petri and R. Haque, 2001. Entamoeba histolytica: Genetic diversity of clinical isolates from Bangladesh as demonstrated by polymorphisms in the serine-rich gene. Exp. Parasitol., 99: 80-88.
CrossRef - Clark, C.G., 2006. Methods for the investigation of diversity in Entamoeba histolytica. Arch. Med. Res., 37: 258-261.
CrossRef - Espinosa-Cantellano, M., G. Castanon Gutierrez and A. Marti�nez-Palomo, 1997. In vivo patogenesis of Entamoeba dispar. Arch. Med. Res., 28: 204-206.
PubMedDirect Link - Espinosa-Cantellano, M., A. Gonzales-Robles, B. Chavez, G. Castanon, C. Arguello A. Lazaro-Haller and A. Mart�nez-Palomo, 1998. Entamoeba dispar: Ultra structure, surface properties and cytopathic effect. J. Eukaryot. Microbiol., 45: 265-272.
PubMedDirect Link - Ghosh, S., M. Frisardi, L. Ramirez-Avila, S. Descoteaux and K. Sturm-Ramirez et al., 2000. Molecular epidemiology of Entamoeba spp: Evidence of a bottleneck (Demographic sweep) and transcontinental spread of diploid parasites. J. Clin. Microbiol., 38: 3815-3821.
Direct Link - Haghighi, A., S. Kobayashi, T. Takeuchi, G. Masud and T. Nozaki, 2002. Remarkable genetic polymorphism among Entamoeba histolytica isolates from a limited geographic area. J. Clin. Microbiol., 40: 4081-4090.
CrossRef - Haghighi, A., S. Kobayashi, T. Takeuchi, G.N. Thammapalerd and T. Nozaki, 2003. Geographic diversity among genotypes of Entamoeba histolytica field isolates. J. Clin. Microbiol., 41: 3748-3756.
CrossRef - Hooshyar, H., M. Rezaeian, B. Kazemi, M. Jeddi-Tehrani and S. Solaymani-Mohammadi, 2004. The distribution of Entamoeba histolytica and Entamoeba dispar in Northern, Central and Southern Iran. Parasitol. Res., 94: 96-100.
CrossRef - Kobayashi, S., E. Imai, A. Haghighi, S. A. Khalifa, H. Tachibana and T. Takeuchi, 2005. Axenic cultivation of Entamoeba dispar in newly designed yeast extract-iron-gluconic acid-dihydroxyacetone-serum medium. J. Parasitol., 91: 1-4.
Direct Link - Pinheiro, S.M., R.F. Maciel, M.A. Morais Jr., I.S. Aca, L.B. Carvalho Jr. and M.R. Coimbra, 2005. Genetic characterization of Entamoeba dispar isolates in Northeast Brazil. Acta Trop., 94: 35-40.
CrossRef - Rasti, S., A. Haghighi, B. Kazemi and M. Rezaeian, 2006. Cloning and Characterization of Entamoeba histolytica Protein Gene from an Iranian Entamoeba histolytica isolate. Pak. J. Biol. Sci., 9: 654-658.
Direct Link - Sambrook, J. and D.W. Russell, 2001. Molecular Cloning: A Laboratory Manual. 3rd Edn., Cold Spring Harbor Laboratory Press, New York, USA., ISBN-13: 9780879695774, Pages: 2344.
Direct Link - Samie, A., C.L. Obi, P.O. Bessong, E. Houpt and S. Stroup et al., 2008. Entamoeba histolytica: Genetic diversity of African strains based on the polymorphism of the serine-rich protein gene. Exp. Parasitol., 118: 354-361.
CrossRefPubMedDirect Link - Simonishvili, S., S. Tsanava, K. Sanadze, R. Chilikadze and A. Miskalishvili et al., 2005. Entamoeba histolytica: The serine-rich gene polymorphism-based genetic variability of clinical isolates from Georgia. Exp. Parasitol., 110: 313-317.
CrossRef - Tanyuksel, M., M. Ulukanligil, H. Yilmaz, Z. Gucliu, R.E. Araz, G. Mert, O. Koru and W.A. Petri, 2008. Genetic variability of the serine-rich gene of Entamoeba histolytica in clinical isolates from Turkey. Turk. J. Med. Sci., 38: 239-244.
Direct Link - Zaki, M. and C.G. Clark, 2001. Isolation and characterization of polymorphic DNA from Entamoeba histolytica. J. Clin. Microbiol., 39: 897-905.
CrossRef - Zaki, M., P. Meelu, W. Sun and C.G. Clark, 2002. Simultaneous differentiation and typing of Entamoeba histolytica and entamoeba dispar. J. Clin. Microbiol., 40: 1271-1276.
CrossRefDirect Link