
Mohammed E. Elnageeb
Department of Physiology, Faculty of Veterinary Medicine, University of Khartoum, Sudan
Abdalla M. Abdelatif
Department of Physiology, Faculty of Veterinary Medicine, University of Khartoum, Sudan
Sharaf Eldin A. Makawi
Department of Obstetrics and Gynaecology, Faculty of Veterinary Medicine, University of Khartoum, Sudan
Pakistan Journal of Biological Sciences
Year: 2008 | Volume: 11 | Issue: 18 | Page No.: 2209-2216







ABSTRACT
Twenty sexually mature Desert ewes were used to study the effects of concentrate supplementation on thermoregulation and reproductive traits. The ewes were divided into two equal groups, control and supplemented; the supplemented group received daily 500 g of concentrate mixture (crushed sorghum grain and cotton seed cake). In both groups, oestrus was synchronized with hormonal method and the ewes were naturally inseminated. Pregnancy was diagnosed by the ultra-sonography technique. Dietary supplementation resulted in a shorter time of oestrus onset, higher rates of conception, lambing and a significant decrease in gestation length. The fertility and fecundity of ewes were also improved. Supplementations also increased significantly the pre-and post lambing weight of ewes and the birth and weaning weights of the lambs. The body weight, rectal temperature and respiratory rate increased significantly with the advance of pregnancy in both groups of ewes. These indices decreased significantly with the progress of lactation. The supplemented ewes maintained higher values of these indices during pregnancy and lactation.
PDF Abstract XML References Citation
How to cite this article
Mohammed E. Elnageeb, Abdalla M. Abdelatif and Sharaf Eldin A. Makawi, 2008. Thermoregulation and Reproductive Performance of Grazing Desert Ewes (Ovis aries) as Influenced by Concentrate Supplementation. Pakistan Journal of Biological Sciences, 11: 2209-2216.
DOI: 10.3923/pjbs.2008.2209.2216
URL: https://scialert.net/abstract/?doi=pjbs.2008.2209.2216
DOI: 10.3923/pjbs.2008.2209.2216
URL: https://scialert.net/abstract/?doi=pjbs.2008.2209.2216
INTRODUCTION
Under tropical conditions where the bulk of sheep population is maintained, nutrition constitutes a major factor that influences thermoregulation and reproductive performance of ewes. Drought conditions, especially in areas that have been overgrazed, can have deleterious effects on the reproductive cycle of sexually mature animals because of malnutrition.
Pregnancy and lactation may influence the thermoregulation responses of mammals. Also the level of food intake affects metabolic heat production and thermoregulation in ruminants (Abdelatif and Ahmed, 1992; Brosh et al., 1998). Moreover, a close relationship between the level of food intake and the rate of metabolic heat production has been established in Merino sheep (Olthoff et al., 1989). Freetly and Ferrell (1997) reported an increase in energy requirements of ewes during pregnancy; approximately 60% of the increase in heat production was associated with the gravid uterus. Laburn et al. (1992) showed that the ewe`s body temperature declined before and at parturition. Variations in the body temperature of mother and foetus in animals are presumably associated with heat dissipation capacity of the utero-placental circulation and heat generation by the foetus (Faurie et al., 2001).
The reproductive performance of the ewe which is considered as the major component of sheep productivity is usually determined by traits as age at first lambing, conception rate, lambing rate, gestation length, lamb birth weight and lamb weaning weight. The reproductive capacity of ewes can be promoted by the supra-ovulation and increase of conception rate. Follicle populations are very sensitive to nutritional input and folliculogenesis and ovulation rate can be increased by nutritional manipulation (Scaramuzzi et al., 2006). Ovulation rate is dependent on short-term improvement of nutrition during the breeding season (flushing) in ewes which are within the intermediate range of body condition (Gunn, 1983). Flushing may also increase the lambing rate in ewes (Sulieman et al., 1990; El-Hag et al., 1998). Moreover, copious supplementary feeding of ewes during gestation and lactation may reduce considerably the morbidity and mortality rates of lambs (Charring et al., 1992; Binns et al., 2002). In tropical areas, animal owners have limited knowledge regarding the energy and protein requirements of the various classes of livestock and their role in expression of genetic potentials. In addition, the formulation of livestock diets is based on information derived from studies conducted in temperate regions. These values may not necessarily apply to indigenous breeds of animals and the prevailing climatic conditions.
Therefore, studies on the nutritional requirements of sheep are needed so that feeding systems which satisfy the actual requirements can be established.
Sheep in Sudan play an important economic and social role and constitute valuable resource for local and export purposes. They provide a popular source of meat for local consumption and an important source of foreign currency. During dry summer conditions they may be exposed to shortage of food which influences their energy budget and reproductive performance. The production characteristics of Sudan desert sheep may reflect the seasonal nutritional status and husbandry system (El-Hag et al., 2001). Accordingly the objective of this study was to evaluate the effect of long term supplementation with a concentrate mixture rich in energy and protein on thermoregulation and some reproductive traits of desert ewes.
MATERIALS AND METHODS
Animals and management: Twenty sexually mature multiparous Desert ewes were used in this study during January 2006-January 2007. The ewes were 2½-3 years old with an average body weight of 35.2 3.6 kg at the beginning of the study. They were selected from the breeding stock of the Sheep Research Unit at Khartoum University Farm (latitude 15°40` north, longitude 32°32` East and 380 m above mean sea level). The animals were apparently healthy. However, the ewes were subjected to quarantine measures for one month and were thoroughly examined to eliminate sick animals and to ascertain that they were fertile and free from reproductive abnormalities. The ewes were periodically dewormed with anthelmintic (Ivomec, Anupco-England: l mL/50 kg BW) and they were given prophylactic antimicrobial treatment (Sulphadiamadin pyrimidine, Richter-pharma, Austria: 1 mL 10 kg BW). The ewes were identified individually using ear-tags and were randomly assigned to two experimental groups. Group A served as control and was kept on grazing only while group B was supplemented with a concentrate mixture. The ewes were kept in open shed with adequate ventilation to facilitate the dissipation of sensible heat and disposal of water vapour. The housing system was provided with appropriate facilities for feeding and watering.
Feeding: Normal nutrition of experimental animals (groups A and B) was provided by grazing harvested residues before being irrigated for regrowth or ploughed for the next sowing season. All animals were put on pasture and allowed to graze daily on the residues of sorghum (Sorghum bicolor) and grasses (Cyndon dactylon) for 6 h (from 8:00 am to 2:00 pm). The ewes in group B were offered a supplemental concentrate mixture composed of a high energy source and a source rich in protein. Each animal in group B received daily at 7:00 am. A mixture of 250 g of crushed sorghum grain and 250 g of cotton seed cake. Salt blocks (macro-minerals, micro-minerals and vitamin D) were also provided to the supplemented group in the animal pens. Tap water was available all the time in the shed. Table 1 shows the chemical composition of the components of the concentrate diet.
| Table 1: | The chemical composition of the components of concentrate diet (g kg-1 DM) |
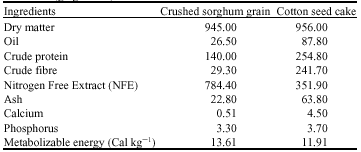 | |
Body Weight (BW), rectal temperature (Tr) and Respiration Rate (RR): During the experimental period, the ewes and their lambs were weighed to the nearest±0.05 kg using a spring balance (Salter-England). The measurements of rectal temperature (Tr) of experimental animals were made to the nearest±0.1°C using a certified mercury-in-glass clinical thermometer (Wilson-Supreme, Japan). The thermometer was inserted into the rectum to a depth of approximately 8 cm in ewes for 2 min. before the reading was obtained. The Respiration Rate (RR) of ewes was measured visually by counting the flank movements with the aid of a stopwatch. The values were taken for one minute of regular breathing with the animal standing quietly.
Synchronization of oestrus, insemination and pregnancy diagnosis: The synchronization of oestrus in ewes was performed by the hormonal method as described by Greyling et al. (1997). The intravaginal progesterone release devices (CDIR, Inter Ag-UK: 300 mg/ewe) were inserted in the ewes for 2 weeks. After withdrawal, ewes were injected with pregnant mare serum gonadotrophin (PMSG, Intervet Ltd. UK: 500 IU/ewe).
During the breeding period, subgroups of the ewes were accommodated in 4 separate pens for 5 days to facilitate oestrus detection and insemination. Each pen accommodated 5 ewes. During this period, oestrus was detected visually by monitoring the signs of heat 3 times daily. Four fertile Hamari breed rams were introduced (at the time of withdrawal of the device), each for a subgroup of ewes. The ewe which showed signs of oestrus (mucus discharge and male seeking behaviour) was allowed to be mated.
The diagnosis of pregnancy was performed one month following natural insemination in order to determine the number of ewes that conceived. Pregnancy was diagnosed by the ultrasonography technique using a standard device (550 Trioga PIE Medica-Netherlands).
Determination of reproductive traits: The reproductive traits were evaluated according to the method of Charring et al. (1992). The conception rate (the percentage of ewes which conceived and got pregnant out of the total number of ewes exposed to the ram) based on non-return rate following the double insemination, was determined 60 days after natural insemination. The lambing rate was determined by calculating the percentage of ewes that gave birth to lambs out of those which conceived. The fertility rate (number of ewes lambing out of those inseminated) was calculated by the following formula: (lambing ewes/ewes inseminated)x100. The fecundity rate (number of lambs born out of number of ewes inseminated) was calculated by the following formula: (lambs born alive/ewes inseminated)x100.
Statistical analysis: The student (t) test was used to compare the reproductive traits of control and supplemented groups of ewes. For the effects of physiological state of each group of ewes on rectal temperature (Tr) Respiration Rate (RR) and Body Weights (BW) the data were subjected to appropriate General Linear Model (GLM) procedure of statistical analysis using the SAS package (1988). The SAS was used to perform Analysis of Variance (ANOVA) to evaluate the effects of physiological state on these parameters. The data considered were at the end of each of the experimental periods: initial (IN), mating (MT), Early Pregnancy (EP), Mid-Pregnancy (MP), Late Pregnancy (LP), parturition (PR) and at the end of the first, second and third month of lactation (L1, L2 and L3, respectively). Also the student (t) test was used to compare the control and supplemented group at each of the physiological states mentioned.
RESULTS
The climatic data (Table 2) indicate that during the experimental period the highest mean value of ambient temperature was measured in May 2006, during dry summer, while the minimum mean value was recorded during January in 2006 and 2007, during winter. Also the minimum mean value of RH (%) was measured in April 2006 during dry summer, whereas the highest mean value was recorded in July and August 2006, during the rainy season.
| Table 2: | The ambient temperature (Ta) and Relative Humidity (RH) during the period January 2006-January 2007 at the experimentation location |
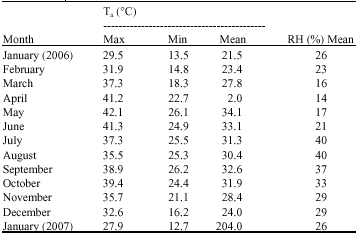 | |
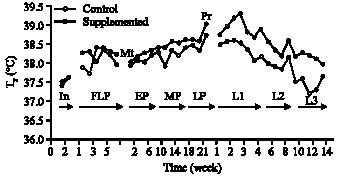 | |
| Fig. 1: | Effect of dietary supplementation during different physiological states on rectal temperature (Tr) of ewes |
Rectal temperature (Tr): There was no significant difference e in the initial values of Tr of the ewes before supplementation of group B animals (Fig. 1). During flushing period, the values of Tr fluctuated in both groups of ewes. There was a significant (p<0.05) increase in Tr values in both groups of ewes during pregnancy to reach maximum values at parturition. During the first and second month of lactation, there was a significant (p<0.05) decrease in Tr of the control ewes, but there was a significant (p<0.05) increase by the end of the third month. The supplemented group showed a progressive significant (p<0.50) decrease of Tr throughout three months of lactation. During the first and second month of lactation, the supplemented ewes showed significantly (p<0.01) higher Tr compared to the control. However, the difference in Tr between the 2 groups during the 3rd month of lactation was not significant.
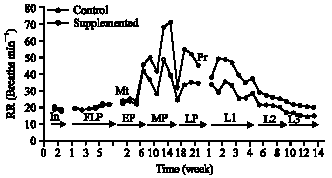 | |
| Fig. 2: | Effect of dietary supplementation during different physiological states on Respiration Rate (RR) of ewes |
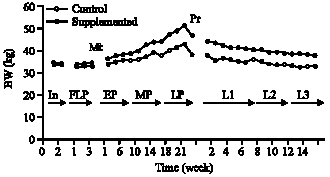 | |
| Fig. 3: | Effect of dietary supplementation during different physiological states on Body Weight (BW) of ewes |
Respiration Rate (RR): There was no significant effect of supplementation on the initial values of RR of the ewes (Fig. 2). Starting from beginning of supplementation, both groups of ewes showed a progressive significant (p<0.05) increase in RR to reach peak values in mid gestation. Thereafter, RR showed a significant (p<0.05) decrease during late gestation and at parturition. Both groups showed progressive significant (p<0.05) decrease in RR during lactation period. The supplemented ewes showed significantly higher RR values during mid gestation (p<0.001) and at parturition and lactation (p<0.01).
Body Weight (BW) of ewes: Figure 3 shows the effects of dietary supplementation at different physiological states on the mean BW of the ewes. There was no significant difference between the mean BW of the groups before supplementation of group B. In both groups of ewes, there was a significant (p<0.05) increase in the mean BW after mating and during different stages of pregnancy. After parturition, the mean BW of both groups of ewes decreased significantly (p<0.05) to reach lower values by the third month of lactation. Also it was noted that the mean BW of supplemented ewes was significantly (p<0.05) higher compared to control ewes during different stages of pregnancy and lactation. The pre-lambing and post-lambing weights of ewes were significantly (p<0.001) higher in the supplemented ewes compared to the control.
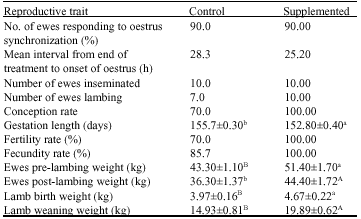
| Table 3: | Effects of dietary supplementation on reproductive performance of Desert ewes |
 | |
| Paired means within the same row bearing different superscripts (small) are significantly different at p< 0.05, Paired means within the same row bearing different superscripts (capital) are significantly different at p<0.001 | |
Reproductive performance: Table 3 indicates that the initial numbers of animals used in the studies were 10 ewes for control (kept on grazing only) and 10 ewes maintained on similar grazing conditions and supplemented with concentrate mixture before breeding and throughout the periods of gestation and lactation.
There was no significant difference between the two groups of ewes in the oestrus response to the synchronization, but the time of oestrus onset was shorter in supplemented ewes compared to the value measured for the control group. The pregnancy diagnosis test indicated that the conception rate was higher in the supplemented group compared to the control (7 ewes for control vs 10 ewes for supplemented). Dietary supplementation decreased the gestation period significantly (p<0.05); the mean values for the control and supplemented group were 155.7±0.3 and 152.8±0.4 days, respectively. The supplementation also increased the percentage of ewes lambing, which was 70% for control and 100% for the supplemented group. The percentages of fertility rate and fecundity were higher in supplemented ewes.
Table 3 also indicates that the birth weight was significantly (p<0.001) higher for lambs born to supplemented ewes compared to those born to the control group. The lambs born to the supplemented ewes also had significantly (p<0.001) higher weaning weight with a mean value of 19.89±0.6 kg compared to a mean weaning weight of 14.93±0.8 kg for lambs born to the control ewes.
DISCUSSION
Thermoregulation in ewes was influenced by pregnancy, lactation and dietary supplementation. Figure 1 and 2 indicate that there was a significant increase in rectal temperature (Tr) and Respiration Rate (RR) during pregnancy in both groups of ewes. The increases in Tr and RR are attributed to increase in metabolic heat production. The results are in line with previous studies that have reported an increase in metabolic heat production during pregnancy in ewes (Gilbert et al., 1985; Freetly and Ferrell, 1997). Furthermore, Rattray et al. (1974) reported that the extra heat production that occurs in pregnant ewes is primarily foetal in origin. The foetus loses its heat via the foetal circulation and uterine wall (Schroeder et al., 1988), foetal heat production represents an extra thermal load on mother heat production. The increase in RR during gestation could be related to the fact that pregnancy limits the excursion of the diaphragm during inspiration and when expansion of the lungs is restricted, adequate ventilation is maintained by increased frequency.
The results showed a significant increase in Tr of ewes at parturition. This is presumably attributed to the process of labour (i.e., hormonal effects) and muscle contraction during parturition. Marx and Loew (1975) reported that the stress of labour and myometrial contraction may produce sufficient heat in the active tissues that will increase body temperature. The present results support the findings of Laburn et al. (1994), who reported that the body temperature of ewes rose at 0.7°C h-1 in the final stage of labour. The decrease in Tr (Fig. 1) and RR (Fig. 2) after lambing and during lactation may be attributed to decrease in uterine blood flow and elimination of heat dissipation from the foetus as a result of delivery. Foetal lambs lose about 85% of their body heat via transfer to the placenta and 15% through the uterine wall (Shroeder et al., 1988). The progressive decrease in Tr and RR during lactation is related to metabolic adjustment and hormonal changes associated with change in milk yield. The relatively higher values of Tr and RR during lactation in the supplemented group is a reflection of the higher plane of nutrition and concomitant rise in milk yield. Gunn and Gluckman (1983) and Laburn et al. (1992) reported that the daily peak in Tr in ewes was influenced by the heat increment of feeding; both feeding and postural changes can affect Tr in ewes.
The results demonstrated that pregnancy had marked effects on the BW of the ewes (Fig. 3). The mean BW increased significantly with the advance of pregnancy in both groups of ewes. The increase in BW after mating and during early gestation is related to the development of conceptus, which was associated with increase in metabolism and body weight gain and to an increase in food intake. An increase in voluntary food intake has been reported to be associated with increase in BW of RambouilletexColumbia ewes during early gestation (Lodge and Heaney, 1973). However, in the present study, the increase in BW could be partly accounted for by changes in the weight of the uterus and foetus. Lodge and Heaney (1973) reported that at parturition, the weight of the contents of conceptus exceeded live weight gain of ewes indicating a loss in BW of ewes during the last quarter of gestation as a direct result of pregnancy, which is also associated with BW loss during mid- and late stages of pregnancy (Robinson et al., 1978). The higher BW of supplemented group of ewes reported in the present study during pregnancy is clearly attributed to the dietary factors. The results are in agreement with the findings of Quirke (1979), who reported that in pregnant ewes, there was an increase in BW with increase in the level of energy and protein intake. The rapid development of the conceptus during mid- and late gestation results in a marked loss of the maternal tissue and consequently in BW losses (Lodge and Heaney, 1973). The significant decrease in BW of both groups of ewes during lactation (Fig. 3) is associated with involution of the uterus and increase in tissue catabolism as a consequence of biosynthesis of colostrum and milk. The current result is in conformity with John et al. (2000), who indicated that during early lactation the ewe is generally unable to consume enough feed to meet the energy demands of high milk production and she loses body weight mostly due to metabolism of fat stores. The higher BW of the supplemented group of ewes during lactation indicates that the concentrate provided sufficient nutrients to reduce the rate of body weight loss. Since the nutrient demands for biosynthesis of milk decreases with the advance of lactation, the rate of BW loss decreased progressively. Hassan (1996) reported that the increase in body weight gain in lactating desert ewes was associated with increase in intake of metabolizable energy.
The current results indicate that some of the reproductive traits of the Desert ewes were improved by dietary supplementation. The data projected in Table 3 indicate that there was no significant difference between the 2 groups in oestrus response to synchronization. According to the appearance of oestrus signs, however, the interval from the injection of PMSG to the onset of oestrus was shorter in supplemented ewes (25.2 h) compared to the time recorded for the control (28. 3 h). The decrease in the time of onset of oestrus is presumably attributed to the nutrition-induced hormonal changes (gonadotropins, oestrogens). It has been established that oestrogen, which is secreted from the mature follicles in the ovary, is responsible for the signs of oestrus (Jainudeen and Hafez, 1993). Haresign (1981) and Downing and Scaramuzzi (1991) reported that nutritional factors (e.g., protein) might influence the development of growing follicles. Increased nutritional level may result in increasing rate of degradation of steroids and reduction in the inhibition of hypothalamic-pituitary axis and consequently resulting in increased gonadotrophin secretion (Thomas et al., 1987). Furthermore, high dietary protein level may increase the responsiveness of the pituitary to Gonadotrophins Releasing Hormone (GRH), resulting in high plasma level of luteinizing hormone (Jordan and Swenson, 1979). In addition, the plane of nutrition may also influence the ovarian response to circulating gonadotropins (Harrison and Randel, 1986; Cox et al., 1987). However, the change in nutrition can influence ovarian activity by changes in circulating concentrations of metabolic hormones, including insulin, IGF-1, growth hormone and Leptin (Webb et al., 2004).
The rates of conception and lambing were higher in supplemented ewes compared to the control (Table 3). This is attributable to the effect of improved nutritional status of the ewes and hence increases in ovulation and embryo survival. These results are in line with previous studies which reported that increasing the level of nutrition increased the rate of ovulation in Border LeicesterxMerino ewes (Rizzoli et al., 1976) and in Desert ewes (Sulieman et al., 1990). Furthermore, Kenney et al. (1980) supplemented Border Leicester x Merino ewes with lupin grains for two weeks prior to and during mating period and they reported an increase in the rates of ovulation and lambing. Since the fertility and fecundity of mammals are usually associated with conception rate, lambing rate and litter size (Gunn, 1983), the present findings suggest that dietary supplementation increased the rates of fertility and fecundity of Desert ewes.
Earlier studies have shown that variation in gestation length in indigenous breeds of sheep is negligible (Khalafalla and Sulieman, 1992). However, the present study showed that dietary supplementation significantly decreased the gestation length in Desert ewes (Table 3). This is attributed to the rapid development and growth of the foetus associated with maturity of the Hypothalamic-Pituitary-Adrenal (HPA) axis and improvement of the live weight of the ewes. The foetal HPA axis plays a primary role in determining the time of parturition by secreting increased amount of corticosteroids (Thorburn and Challis, 1979; McLaren et al., 1996). Hawkins et al. (1999) pointed out that modest restriction of maternal nutrition (15%) during early pregnancy depresses HPA axis function in the foetus in late gestation. Wallace et al. (1997) reported that rapid growth rate of adolescent ewes was associated with decrease in gestation length.
In the present study (Table 3), the birth weight of lambs born to supplemented ewes (4.67 kg) was higher compared to the birth weight of lambs born to ewes kept on grazing only (3.97 kg). This is clearly related to the high nutritional status and the improved body condition of supplemented ewes during pregnancy. Ocak et al. (2005) pointed out that maternal nutrition during pregnancy is a major extrinsic factor that influences the birth weight in domestic livestock. In sheep, the birth weight of lambs increased with increase of gestation live weight of dams (Mukasa-Mugerwa et al., 1994).
The data indicate than the mean body weight of the lambs born to supplemented ewes at weaning was significantly higher compared to the lambs born to control ewes (Table 3). This could be attributed to the influence of ewes` BW. Earlier studies indicated that there was a positive correlation between birth weight and weaning weight of lambs and the ewe BW (Farid and Makarechina, 1978; Ploumi and Emmanouilidis, 1999). The milk yield of dams was not measured in the present study. However, it could be assumed that the higher plane of nutrition of supplemented ewes was associated with higher milk yield. During the early weeks of life, there is high correlation between growth rate of lambs and milk yield of dams as lambs consume very little solid feed during this period. Previous studies indicated that the growth of lambs was closely related to the milk yield of ewes (Cowan et al., 1980; Snowder and Glimp, 1991). The growth rate of lambs is primarily determined by energy intake relative to the live weight (Hassan, 1996).
CONCLUSION
The studies indicate that pregnancy and lactation influence thermoregulation and body weight of Desert ewes. Dietary supplementation could influence thermoregulation and improve reproductive performance of ewes, particularly when it is maintained throughout gestation period. Further research is needed to investigate the impact of summer thermal load on endocrine responses and reproductive performance of Desert ewes.
ACKNOWLEDGMENTS
We would like to thank the laboratory staff of the Department of Physiology, Faculty of Veterinary Medicine for technical assistance. Also we acknowledge gratefully the help rendered by Dr. K.A. Abdoun in presentation of the manuscript.
REFERENCES
- Binns, S.H., I.J. Cox, S. Rizvi and L.E. Green, 2002. Risk factors for lamb mortality on UK sheep farms. Prev. Vet. Med., 52: 287-303.
Direct Link - Brosh, A., Y. Aharoni, A.A. Degen, D. Wright and B.A. Young, 1998. Effects of solar radiation, dietary energy and time of feeding on thermoregulatory responses and energy balance in cattle in a hot environment. J. Anim. Sci., 76: 2671-2677.
PubMedDirect Link - Cowan, R.T., J.J. Robinson, I. McDonald and R. Smart, 1980. Effects of body fatness at lambing and diet in lactation on body tissue loss, feed intake and milk yield of ewes in early lactation. J. Agric. Sci., 95: 497-514.
CrossRef - Cox, N.M., M.J. Stuart, T.G. Althen, W.A. Bennett and H.W. Miller, 1987. Enhancement of ovulation rate in gilts by increasing dietary energy and administering insulin during follicular growth. J. Anim. Sci., 64: 507-516.
PubMed - Downing, J.A. and R.J. Scaramuzzi, 1991. Nutrient effects on ovulation rate, ovarian function and the secretion of gonadotrophic and metabolic hormones in sheep. J. Reprod. Fert. Suppl., 43: 209-227.
PubMed - El-Hag, F.M., B. Fadlalla and H.K. Mukhtar, 2001. Some production characteristics of Sudan desert sheep under range conditions in North Kordofan, Sudan. Trop. Anim. Health Prod., 33: 229-239.
CrossRefDirect Link - Faurie, A.S., B. Mitchell and H.P. Laburn, 2001. Foet-maternal relationships in goats during heat and cold exposure. Exp. Physiol., 86: 199-204.
Direct Link - Farid, A. and M. Makarechina, 1978. The relationship between pre-and post-weaning growth traits in lambs of Iranian fat-tailed sheep and their crosses with Corriedale and Targhee rams. Anim. Prod., 26: 185-192.
CrossRef - Freetly, H.C. and C.L. Ferrell, 1997. Oxygen consumption by and blood flow across the portal drained viscera and liver of pregnant ewes. J. Anim. Sci., 75: 1950-1955.
PubMed - Gilbert, R.D., H. Schroder, T. Kawamura, P.S. Dale and G.G. Power, 1985. Heat transfer pathways between foetal lamb and ewe. J. Applied Physiol., 59: 634-638.
PubMed - Greyling, J.P.C., J.A. Erasmus, G.J. Taylor and S. van der Merwe, 1997. Synchronization of estrus in sheep using progestagen and inseminating with chilled semen during the breeding season. Small Rumin. Res., 26: 137-143.
CrossRefDirect Link - Gunn, T.R. and P.D. Gluckman, 1983. Development of temperature regulation in the foetal sheep. J. Dev. Physiol., 5: 167-179.
PubMed - Haresign, W., 1981. The influence of nutrition on reproduction in the ewe. 2 Effects of undernutrition on pituitary responsiveness to luteinizing hormone releasing hormone stimulation. Anim. Prod., 32: 257-265.
CrossRef - Harrison, L.M. and R.D. Randel, 1986. Influence of insulin and energy intake on ovulation rate, luteinizing hormone and progesterone in beef heifers. J. Anim. Sci., 63: 1228-1235.
Direct Link - Hawkins, P., C. Steyn, H.H. McGarrigle, T. Saito and T. Ozaki et al., 1999. Effect of maternal nutrient restriction in early gestation on development of the hypothalamic-pituitary-adrenal axis in fetal sheep at 0.8-0.9 of gestation. Endocrinology, 163: 553-561.
CrossRefPubMedDirect Link - John, C., C. Wand, H. Aitken and S. Cieslar, 2000. Dairy sheep nutrition. Proceedings of the 7th Great Lake Dairy Sheep Symposium. November 2-4, 2000, Guelph, Ontario, Canada, pp: 1-6.
Direct Link - Kenney, P.A., J.L. Reeve, R.W. Baxter and I.A. Cumming, 1980. Effect of different levels of supplementation of lupin grain, lucerne, wheat, wheat with urea and sulphur fed during mating in February to Border Leicester X Merino ewes in North East Victoria. Aust. J. Exp. Agric. Anim. Husb., 20: 15-19.
- Laburn, H.P., D. Mitchell and K. Goelst, 1992. Fetal and maternal body temperatures measured by radiotelemetry in near-term sheep during thermal stress. J. Appl. Physiol., 72: 894-900.
PubMed - Lodge, G.A. and D.P. Heaney, 1973. Energy cost of pregnancy in single-and twin-bearing ewes. Can. J. Anim. Sci., 53: 479-489.
CrossRefDirect Link - Marx, G.F. and D.A.Y. Loew, 1975. Tympanic temperature during labour and parturition. Br. J. Anaesth., 47: 600-602.
PubMed - Mukasa-Mugerwa, E., A.N. Said, A. Lahlou-Kassi, T. Sherington and E.R. Mutiga, 1994. Birth weight as a risk factor for perinatal lamb mortality, and the effects of stage of pregnant ewe supplementation and gestation weight gain in Ethiopian Menz sheep. J. Prev. Vet. Med., 19: 45-56.
CrossRef - Ocak., N., M.A. Cam and M. Kuran, 2005. The effect of high dietary protein levels during late gestation on colostrums yield and lamb survival rate in singleton-bearing ewes. Small Rumin. Res., (1/3)56: 89-94.
Direct Link - Olthoff, J.C., G.E. Dickerson and J.A. Nienaber, 1989. Energy utilization in mature ewes from seven breeds with diverse production potentials. J. Anim. Sci., 67: 2550-2564.
PubMed - Rattray, P.V., N.N. Garrett, N.E. East and N. Hinman, 1974. Efficiency of utilization of metabolizable energy during pregnancy and the energy requirements for pregnancy in sheep. J. Anim. Sci., 38: 383-393.
PubMed - Robinson, J.J., I. McDonald, I. MeHattie and R. Penmie, 1978. Studies on reproduction in prolific ewes. 4. Sequential changes in the maternal body during pregnancy. J. Agric. Sci. (Camb.), 91: 291-304.
CrossRef - Schroeder, H., R.D. Gilbert and G.G. Power, 1988. Computer model of foetal maternal heat exchange in sheep. J. Applied Physiol., 65: 460-468.
PubMed - Scaramuzzi, R.J., B.K. Campbell, J.A. Downing, N.R. Kendall, M. Khalid, M. Munoz-Gutierrez and A. Somchit, 2006. A review of the effects of supplementary nutrition in the ewe on the concentrations of reproductive and metabolic hormones and the mechanisms that regulate folliculogenesis and ovulation rate. Reprod. Nutr. Dev., 46: 339-354.
CrossRefPubMedDirect Link - Thomas, D.L., P.J. Thomford, J.G. Crickman, A.R. Cobb and P.J. Dziuk, 1987. Effects of plane of nutrition and phenobarbital during the pre-mating period on reproduction in ewes fed differentially during the summer and mated in the fall. J. Anim. Sci., 64: 1144-1152.
PubMed - Thorburn, G.D. and J.R.D. Challis, 1979. Endocrine control of parturition. Physiol. Rev., 59: 863-918.
PubMed - Wallace, J.M., P. Da Silva, R.P. Aitken and M.A. Cruickshank, 1997. Maternal endocrine status in relation to pregnancy outcome in rapidly growing adolescent sheep. J. Endocrinol., 155: 359-368.
PubMed - Webb, R., P.C. Garnsworthy, J.G. Gong and D.G. Armstrong, 2004. Control of follicular growth: Local interactions and nutritional influences. J. Anim. Sci., 82: E63-E74.
Direct Link