
Amany H.A.Abo Ellil
Botany Department, Faculty of Science, Cairo University, Giza, Egypt.
Neveen S.I. Geweely
Botany Department, Faculty of Science, Cairo University, Giza, Egypt.
Pakistan Journal of Biological Sciences
Year: 1999 | Volume: 2 | Issue: 2 | Page No.: 290-295







ABSTRACT
High alkalinity inhibited sugar utilization by the alkalosensitive fungus (P. albicans) and increased the polysaccharide content in its fungal mats. The total amino-N content decreased in the culture of P. albicans, whereas its quantity increased in the biomass with progressive alkalinity. Alkalinity inhibited nitrogen uptake and protein synthesis leading to a corresponding decline in growth, and hence apparent alkalosensitivity. High alkalinity, caused enhancement of sugar uptake by the facultative alkolophile V. lateritium associated with higher reducing sugar contents and decreased polysaccharide accumulation in mycelial mats. The amino-N content increased in the culture medium of V. lateritium and decreased in the biomass with pH elevation. The utilization of soluble nitrogen increased, accompanied by increased protein synthesis, leading to maximum growth at higher pHs, and hence apparent alkalophilicity. The sodium ion content in the medium of the alkalosensitive fungus decreased at high pHs associated with respective increase in the biomasses. Oppositely, in the alkalophilic fungus, the sodium ion content increased in culture and decreased in biomasses with increased alkalinity, indicating an important role played by this ion. No clear relation could be obtained between phosphorus content in cultures or biomasses of the two tested fungi and alkalophilicity.
PDF Abstract XML References Citation
How to cite this article
Amany H.A.Abo Ellil and Neveen S.I. Geweely, 1999. Comparative Biochemical Studies on Penicillium albicans (Alkalosensitive) and Verticillium lateritium (Facultative Alkalophile). Pakistan Journal of Biological Sciences, 2: 290-295.
DOI: 10.3923/pjbs.1999.290.295
URL: https://scialert.net/abstract/?doi=pjbs.1999.290.295
DOI: 10.3923/pjbs.1999.290.295
URL: https://scialert.net/abstract/?doi=pjbs.1999.290.295
INTRODUCTION
The mechanism that allows the organism to adapt to extreme environments, such as highly alkaline pH values, are one of the most interesting subjects for microbiologists. Studies on alkalophily have been primarily focused on bacteria such as Bacillus circulaus (Chislett and Kushner, 1961), B. alcalophiles (Takahara and Tanabe, 1961). Later on, alkaline tolerant fungi and yeasts were isolated from soils and poultry excrements (Matsushima et al., 1980; Goto et al., 1981; Horikoshi and Akiba, 1982). Miyashita et al. (1984) isolated alkalophilic actinomycete strains from Japanese soils being capable of growing in synthetic and organic media between pH 7 and 11 but not at pH 6.5. With respect to alkalophily among fungi, different degrees of pH effect on mycelial growth and conidial germination in Fusarium spp., Penicillium spp., Aspergillus spp. and Verticillium spp. are recorded (Thompson et al., 1993; Wheeler et al., 1991; Nowak and Hurle, 1990; Dai et al., 1991). Different explanations have been proposed interpreting alkalophily mostly in bacteria (Guffanti et al., 1980, Krulwich et al., 1982) and less in fungi (Verticillium lacanii by Sato et al. (1983).
In the present investigation, some physiological and biochemical aspects were studied in the alkalosensitive fungus Penicillium albicans and the facultative alkalophilic fungus Verticillium lateritium after growth at various pHs aiming to clarifying differences in behaviour of such fungi which may explain alkalosensitivity or alkalophilicity. Searching for such physiological factors, the fungal dry biomasses and the growth media were analyzed for their carbon, nitrogen, phosphorus and sodium contents at different pHs.
MATERIALS AND METHODS
Test organisms: Verticillium lateritium, isolated and identified by Neveen (1997) from alkaline Egyptian soil as facultative alkalophile, since its biomass gain increased progressively with pH elevation to reach maximum value at 11.5. It grew minimally at pH 6.5.
Penicillium albicans, isolated and identified (Neveen, 1997) from alkaline Egyptian soil as an alkalosensitive fungus since optimum growth occurred around pH 7.5 after which a decrease in biomass was recorded with increased alkalinity and the growth stopped completely at pH 11.5.
Media: The organisms were cultivated and maintained on a medium prepared according to Horikoshi and Akiba (1982) contained (g/L): soluble starch 10.0, peptone 5.0, yeast extract 5.0, KH2PO4 1.0, MgSO4. 7H20 0.20; Na2CO3 10.0, agar (if needed) 20.0, distilled water 1 L, Na2CO3 must be sterilized separately. The pH of that medium ranged from 10.3 to 10.5. Unless otherwise stated the liquid medium used for growing the organisms was the same but without agar.
Adjustment of the growth medium to different pH values was carried out using two buffer systems, Na2CO3 - Na2HCO3 and NaH2PO4 - NaOH (Okada et al., 1993). The buffers were separately sterilized by filtration before addition in suitable aliquots to the basal sterile medium to give pHs 6.5, 7.5, 8.5, 9.5, 10.5 and 11.5 (Khodair et al., 1991).
Biochemical analysis of fungal mats and culture filtrates: All fungal mats as well as culture media were analyzed for their nitrogen and sugar contents. The fungal mats were further analyzed for their polysaccharide and insoluble nitrogen contents.
Carbohydrate analysis: Extraction of mats was carried out according to the procedure recommended by Naguib (1963), (ii) nitrogen analysis and soluble phosphorus was carried out according to Naguib (1969).
Methods of Analysis: All nitrogen, phosphorus and carbohydrate components were estimated spectrophotometrically. Triplicate samples were used in each estimation. (I) Carbohydrate analysis was carried out according to Clark (1964). (ii) Nitrogen analysis was carried out by the Berthelot reaction (Fawcett and Scott, 1960; Chaney and Marbach, 1962). (iii) Total amino-N analysis. was carried out according to the method of Russell (1944). (iv) phosphorus estimation by the sulfite metal method adopted by Burton and Riley (1954). (v) Sodium estimation by the method of Macdonald and Sirichanya (1969).
Statistical Analysis: The data generated in these studies were suitable for the least significant difference (L.S.D.).
RESULTS
Analysis of culture media and fungal mats of the tested fungi after 7 day growth on media adjusted at different pH values were carried out.
Analysis of culture media: In V. lateritium, the DRV increased with the increase in pH up to 11.5 (Table 1). The direct reducing sugars by the end of the growth period of P. albicans (7 day) revealed its highest value (479 μg/mL) at pH 10.5 followed by the medium adjusted to pH 11.5. The least DRV (376 μg/mL) was achieved in the medium with pH 6.5.
The uptake of reducing sugar (%) showed an opposite relationship with pH increase in the growth medium of P. albicans (Table 1). The highest uptake (59%) and the lowest (43%) were achieved at pHs 6.5 and 11.5, respectively. The three lower experimental pHs (6.5, 7.5, 8.5) in the culture media of V. lateritum were accompanied with less sugar uptake than the three higher pHs (9.5, 10.5, 11.5). The least sugar uptake (53%) and the highest one (96%) were recorded at pH 6.5 and 11.5, respectively. Table 1 shows the effect of pH variation on the amino-N contents in the growth medium of the 2 test fungi after 7 day growth on starch containing Horikoshi medium, initially included with peptone as a N-source. Two opposite trends were exhibited by the tested fungi. The first began with a maximum amino-N content followed by a gradual drop with the increase in alkalinity to reach a minimum value at the highest pH (11.5) as in case of P. albicans. The second trend was exhibited by V. lateritium where the amino-N content generally increased significantly in the media as the pH increased up to 10.5.
Table 1 also shows that the total soluble nitrogen (TSN) content in the culture media of P. albicans increased with pH elevation to reach a maximum value (430 μg/ml) at pH 11.5. The minimum value (110 μg/ml) was recorded at pH 6.5. V. lateritium revealed a decrease in TSN with the increase in pH of media up to pH 11.5 at which a minimum value (220 μg/ml) was measured. At pH 6.5 and 7.5 the total soluble nitrogen recorded the highest quantities.
Table 1 presents the effect of pH variation on the total phosphorus ions in the culture media of the two fungi. KH2PO4 (1 g/L) was initially included in the medium as a source of phosphorus. In case of P. albicans, the maximum total phosphorus was obtained in media adjusted to pHs 6.5 and 9.5 (81 and 80 μg/ml, respectively). The other pHs were accompanied with less contents of phosphorus. The total phosphorus in the growth media of V. lateritium showed the only significant change at the highest pH (11.5) where 35.2 μg/ml were measured. No statistically detectable changes were recorded at the other experimental pHs.
Table 1 includes the effect of pH variation on the amount of sodium ions in the culture of the two fungi. Na2CO3 was the initial source of sodium in the medium. The amount of Na+ in the growth media of P. albicans showed nonsignificant changes with pH increase up to 10.5 where the highest Na+ accumulation (940 μg/ml) was recorded. At pH 11.5 a highly significant drop in Na+ content was observed (110 μg/ml medium). V. lateritium behaved in an opposite trend, where Na+ accumulation increased with elevation of pH to reach a maximum value at pH 10.5 (980 μg/ml medium). The least accumulation of Na+ was at 6.5 (100 μg/ml).
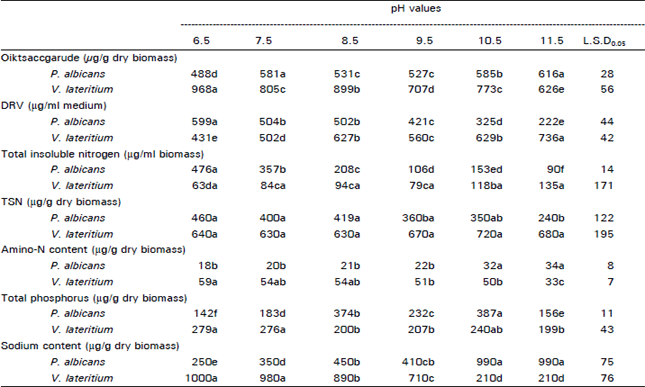
Analysis of mycelium: The dry mycelial mats of the two tested fungi were analyzed for the direct reducing sugars (DRV), polysaccharides, amino nitrogen, total soluble nitrogen (TSN), total insoluble nitrogen, phosphorus and sodium contents.
In P. albicans, the low pHs (6.5, 7.5 and 8.5) were associated with high mycelial contents of DRV with a maximum value (599 μg/g) at pH 6.5 (Table 2). As the alkalinity increased the DRV decreased with a minimum value at pH 11.5 (222 μg/g). The DRV in mats of V. latentium increased with increase in pH of medium to reach a maximum value (736 μg/g) at pH 11.5.
At high alkalinity levels, significant polysaccharide contents were detected in mats of P. albicans which decreased at lower pHs (Table 2). Oppositely, the dry mycelial mats of V. lateritium contained significantly high polysaccharide contents which decreased with increased alkalinity of the growth media. Maximum polysaccharide contents (968 μg/g) were attained at pH 11.5.
The amino nitrogen contents in the mycelial mats of P. albicans increased concomitantly as the alkalinity of the growth media being increased (Table 2). Maximum mycelial amino-N (34 μg/g) was recorded at pH 11.5 while minimum value (18 μg/g) was obtained at pH 6.5. In case of V. lateritium, an opposite trend to the aforementioned fungus was exhibited. The mycelial amino-N content decreased with the increase in alkalinity to reach a minimum value (33 μg/g) at pH 11.5. Highest amino-N contents in dry mats of the fungus (59 μg/g) were obtained at pH 6.5.
The total soluble nitrogen contents decreased in the mycelial mats of P. albicans associating increased pH of growth media (Table 2). The highest value (460 μg/g) was recorded at pH 6.5, while the least TSN (240 μg/g) was found at pH 11.5. In case of V. lateritium, the total soluble nitrogen fraction remained statistically unaltered with pH variation at all experimental values. On the other hand and in P. albicans, the increase in alkalinity led to decrease in mat content of the insoluble nitrogen fraction (protein). Hence, maximum value (476 μg/g) was detected at 6.5 while minimum value (90 μg/g) was obtained at pH 11.5. Oppositely, the total insoluble nitrogen fraction in V. lateritium increased concomitantly with the increase in pH of media to reach its highest quantity (135 μg/g) at pH 65.
The total phosphorus contents in the dry mycelial mats of the test fungi revealed a maximum value in the dry biomass of P. albicans raised at pH 10.5 (387 μg/g) (Table 2). Higher or lower alkalinity levels than 10.5 were accompanied with statistically significant decreases in phosphorus contents in the dry biomass in V. lateritium. With respect to sodium contents in the dry biomass of the test fungi (Table 2), two opposite behaviours being exhibited. In P. albicans the sodium content recorded significant increases with pH increase in the media to reach a maximum value (990 μg/g) at pH 11.5. In case of V. lateritium the sodium content in the biomass decreased with increased alkalinity of growth media to reach a minimum value (210 μg/g) at 11.5. The highest accumulation of sodium ions in the mats of this fungus (1000 μg/g) was detected at pH 6.5 (Table 2).
DISCUSSION
The present investigation revealed significant differences between P. albicans and V. lateritium. High alkalinity inhibited sugar utilization by the alkalosensitive fungus, P. albicans, resulting in growth retardation. In case of alkalophilic V. lateritium, the increase in alkalinity was associated with an increase in reducing sugar in the growth medium, together with marked enhancement of sugar uptake. It could thus be concluded that the increase in media content of DRV at high pHs in case of the latter fungus is not due to low sugar utilization, as it was coupled with maximum growth but due to increased activity of amylase system which has pH optimum at the alkaline side. The DRV content in the biomass of the alkalosensitive fungus decreased and the polysaccharide increased as the alkalinity being increased such decline in soluble sugar content of mycelium may be due to either decreased sugar uptake, increased trend towards sugar polymerization into polysaccharide and/or possible decreased respiration rates. All these factors, each alone or all together, are expected to lead to growth suppression of such alkalosensitive fungus. In the alkalophilic fungus, the reverse trend was obtained,
where high alkalinity was coupled with an increase in reducing sugar content associating parallel decrease in polysaccharide accumulation. This may be due to lowerer rate of sugar polymerization into polysaccharides and activation of polysaccharide hydrolyzing enzymes to ME the increased rate of respiration. In this connection Ohta et al. (1975) reported that respiration by the cells alkalophilic Bacillus sp No. 8-1 increased with increase external pH and was maximum at pH 9.0 in presence glucose or succinate. Kimura and Horikoshi (1988) found that bacterial strains grown on Horikoshi medium at p = 10.0 indicated oxidative metabolism.
The total amino-N either in growth media or in biomasses the two tested fungi revealed that in the alkalosensitive albicans, the increase in media pHs was coupled with decrease in the amount of amino acids in culture filtrate while its quantities increased in the mycelial mats. The observation when coupled with the recorded least grow parameters under such conditions led to the postulation. that I) high alkalinity inhibits protein synthesis, ii) the amino acid content in the growth media may be responsible for alkalophilicity more than mycelial amino acid content, while despite of its increased quantity at high pHs, the fungi growth declined. In case of the alkalophilic V. lateritium, the increase in media pHs led to an increase in its content amino-N. This was accompanied with parallel decrease in biomass contents of free amino-N, a phenomenon which denotes an expected increase in protein synthesis associating optimum growth and confirms the assumption, that high levels of amino acids in culture media play protective role in alkalosensitivity. An evidence in favour this postulation is shown by the amino acid content in the culture of V. lateritium (alkalophilic) which remained at high level up to pH 11.5. Determination of the total soluble nitrogen in culture filtrates and fungal biomasses reveal that in P. albicans, increased pHs led to a concomitant inhibition in nitrogen absorption and metabolism leading minimum growth and apparent alkalosensitivity. In case the alkalophilic V. lateritium, a reverse trend was exhibit. The utilization of soluble nitrogen from the media increase with pH increase. It kept the level of total soluble nitrogen in mycelial mats almost constant at all tested pHs (6.1-11.5) in spite of the increased synthesis of insoluluble nitrogen compounds.
This directs the attention to the important role posses played by soluble nitrogen compounds in alkalophilicity. In this field, Koyama et al. (1976) reported that synthesis protein by alkalophilic bacteria becomes more active in alkaline environment. Horikoshi and Akiba (1982) stated that the comparative studies of protein synthesized systems of alkalophilic bacilli and the neutrophilic Bacil subtilis indicated that the ribosomes of alkalophilic bacill contributed to the higher activity of protein synthesis alkaline pH.
Phosphorus determinations in the culture filtrates and biomasses of the tested fungi revealed no definite trend clear relation between this fraction and increased alkalinity of growth media.
| Table 1: | Effect of pH variation on the uptake of reducing sugars (%) from culture media and direct reducing sugar values (DRV, as glucose), total soluble nitrogen (TNS), amino-N, total phosphorus and sodium contents in the culture media of P. albicans and V. lateritium after 7 d. Growth on starch-containing Horikoshi medium |
 | |
| Means followed by the same letters are statistically non-significant | |
| Table 2: | Polysaccharide contents and direct reducing sugar value (DRV, as glucose), total insoluble nitrogen, total soluble nitrogen (TSN), amino-N contents, total phosphorus and sodium content in the mycelial mats of P. albicans and V. lateritium after 7 d. growth on starch-containing Hoikoshi medium adjusted to different pH values |
 | |
| Means followed by the same letters are statistically non-significant | |
It appears, therefore, that phosphorus has no role in differentiating alkalosensitive or alkalophilic fungi. Guffanti and Hicks (1991) found that the phosphorylation potential of Bacillus firmus was not changed at pH 7.5-10.5 and was in conventional range for bacteria, Krulwich (1982) reported that fine structural studies on the alkalophiles do not indicate the presence of intracytoplasmic organelles that it could be true sites of ATP synthesis.
Sodium ion determination in culture filtrates and fungal biomasses provided an essential difference between alkalosensitive and alkalophilic fungal species. In the alkalosensitive P. albicans the sodium ion content in the media decreased associating a consequent increase in the biomasses. The opposite trend was exhibited by the alkalophilic V. lateritum where sodium ion content increased in the media accompanied by a corresponding decrease in the respective biomasses. This led to a conclusion that at high alkalinity the mycelial mats of the alkalosensitive fungi accumulate high quantity of Na which may suppress growth. Oppositely, the alkalophilic fungus contains low mycelial Na content which may be due to the formation of Na/H antiporter to acidify the cytoplasm under high alkalinity conditions. In V. lateritum, the maximum growth at pH 11.5 was associated with high Na content without any significant drop from its quantity at pH 10.5. This finding indicates the intimate relation of Na and alkalophilicity. In this connection Schuldiner and Fishkes (1978) reported that Na/H antiporter catalyses the inward movement of protons in exchange of Na and that the following gradients are established in alkalophilic cells: Na out > Na in >H out. Non-alkalphiles lack the antiporter and cannot maintain a relatively acidified cytoplasm and hence have lost the ability to grow above pH 9.0. Similar observations and conclusions where also reached by Krulwich et al. (1979), Mandel et al. (1980), Horikoshi and Akiba (1982) and Koyama (1989). They all stated that internal pH of alkalophilic bacteria remains relatively constant over a wide range of external pH values. This indicates a mechanism for the return of the protons into the cell by using antiporter systems for the removal of cations from internal spaces or by exchanging the cations for protons depending on the presence of Na through the operation of an Na/H antiporter. Kitada and Horikoshi (1979) demonstrated Na-dependent uptake of glutamate by whole cells of alkalophilic Bacillus No. 8-1 and Na-dependent transport of 13 amino acids in membrane vesicles from Bacillus alkalophilus. Oshima et al. (1987) found that the growth rate of alkalophilic Bacillus ASSC-2 at alkaline pH was accelerated by addition of NaCI. Horikoshi and Akiba (1982) reported that Na stimulates the uptake of nutrients in alkalophilic Bacillus strains.
REFERENCES
- Chaney, A.L. and E.P. Marbach, 1962. Modified reagents for determination of urea and ammonia. Clin. Chem., 8: 130-132.
PubMedDirect Link - Chislett, M.E. and D.J. Kushner, 1961. A strain of Bacillus circulans capable of growing under highly alkaline conditions. Microbiology, 24: 187-190.
CrossRefDirect Link - Fawcett, J.K. and J.E. Scott, 1960. A rapid and precise method for the determination of urea. J. Clin. Pathol., 13: 156-159.
CrossRefPubMedDirect Link - Goto, S., R. Aono, J. Sugiyama and K. Horikoshi, 1981. Exophiala alcalophila, a new black, yeast-like hyphomycete with an accompanying Phaeococcomyces alcoalophilus morph and its physiological characteristics. Trans. Mycol. Soc. Jap., 22: 429-439.
Direct Link - Guffanti, A.A., R. Blanco, R.A. Benenson and T.A. Krulwich, 1980. Bioenergetic properties of alkaline-tolerant and alkalophilic strains of Bacillus firmus. Microbiology, 119: 79-86.
CrossRefDirect Link - Guffanti, A.A. and D.B. Hicks, 1991. Molar growth yields and bioenergetic parameters of extremely alkaliphilic Bacillus species in batch cultures and growth in a chemostat at pH 10.5. Microbiology, 137: 2375-2379.
CrossRefDirect Link - Kimura, T. and K. Horikoshi, 1988. Isolation of bacteria which can grow at both high pH and low temperature. Applied Environ. Microbiol., 54: 1066-1067.
Direct Link - Kitada, M. and K. Horikoshi, 1979. Relation of glutamate transport to growth of alkalophilic Bacillus species. Agric. Biol. Chem., 43: 2273-2277.
CrossRefDirect Link - Koyama, N., 1989. Na+-dependence of the stability under alkaline conditions of the membrane of an alkalophilic Bacillus. FEMS Microbiol. Lett., 61: 7-10.
CrossRefDirect Link - Koyama, N., A. Kiyomiya and Y. Nosoh, 1976. Na+-dependent uptake of amino acids by an alkalophilic Bacillus. FEBS Lett., 72: 77-78.
CrossRefDirect Link - Krulwich, T.A., K.G. Mandel, R.F. Bornstein and A.A. Guffanti, 1979. A non-alkalophilic mutant of Bacillus alcalophilus lacks the Na+ H+ antiporter. Biochem. Biophys. Res. Commun., 91: 58-62.
CrossRefDirect Link - Macdonald, A.M.G. and P. Sirichanya, 1969. The determination of metals in organic compounds by oxygen-flask combustion or wet combustion. Microchem. J., 14: 199-206.
CrossRefDirect Link - Mandel, K.G., A.A. Guffanti and T.A. Krulwich, 1980. Monovalent cation/proton antiporters in membrane vesicles from Bacillus alcalophilus. J. Biol. Chem., 255: 7391-7396.
Direct Link - Matsushima, K., I. Mori, N. Ito and K. Shimada, 1980. The feature of the proteolytic enzyme system of acid tolerant and alkaline-tolerant fungi: The ecological study on the proteolytic enzymes of microorganisms part II. J. Agric. Chem. Soc. Jap., 54: 875-876.
CrossRefDirect Link - Miyashita, K., Y. Mikami and T. Arai, 1984. Alkalophilic actinomycete, Nocardiopsis dassonvillei subsp. prasina subsp. nov., isolated from soil. Int. J. Syst. Evol. Microbiol., 34: 405-409.
CrossRefDirect Link - Nowak, A. and K. Hurle, 1990. Dependence of the effect of the herbicide afalon on Verticillium sp. on concentration and pH of the growth medium. J. Plant Dis. Prot., 12: 367-371.
Direct Link - Ohta, K., A. Kiyomiya, N. Koyama and Y. Nosoh, 1975. The basis of the alkalophilic property of a species of Bacillus. Microbiology, 86: 259-266.
CrossRefDirect Link - Okada, G., Y. Niimura, T. Sakata, T. Uchimura, N. Ohara, H. Suzuki and M. Kozaki, 1993. Acremonium alcalophilum, a new alkalophilic cellulytic hyphomycete. Trans. Mycol. Soc. Jap., 34: 171-185.
Direct Link - Russell, J.A., 1944. Note on the colorimetric determination of amino nitrogen. J. Biol. Chem., 156: 467-468.
Direct Link - Sato, M., T. Beppu and K. Arima, 1983. Studies on antibiotics produced at high alkaline pH. Agric. Biol. Chem., 47: 2019-2027.
CrossRefDirect Link - Schuldiner, S. and H. Fishkes, 1978. Sodium-proton antiport in isolated membrane vesicles of Escherichia coli. Biochemistry, 17: 706-711.
CrossRefDirect Link - Thompson, D.P., L. Metevia and T. Vessel, 1993. Influence of pH alone and in combination with phenolic antioxidants on growth and germination of mycotoxigenic species of Fusarium and Penicillium. J. Food Protect., 56: 134-138.
CrossRefDirect Link - Wheeler, K.A., B.F. Hurdman and J.I. Pitt, 1991. Influence of pH on the growth of some toxigenic species of Aspergillus, Penicillium and Fusarium. Int. J. Food Microbiol., 12: 141-149.
CrossRefDirect Link