
Mohammed A. Alsaif
Department of Community Health
Sciences, College of Applied Medical
Sciences, King Saud University,
P.O. Box 10219, Riyadh-11433, Saudi Arabia
Journal of Medical Sciences
Year: 2007 | Volume: 7 | Issue: 7 | Page No.: 1164-1170







ABSTRACT
The aim of the present study is to investigate the protective effects of thymoquinone on ethanol induced hepatotoxicity in Wistar rats. The rats were pretreated orally (using rats feeding needle) with different doses (5, 10 and 20 mg kg-1 body weight) of TQ for 7 days and then ethanol (7.5 g kg-1) was injected. Silymarin was used as standard hepato-protective agent for comparison. The activities of Aspartate Aminotransferase (AST), Alanine Aminotransferase (ALT), Acid Phosphatase (ACP) and Alkaline Phosphatase (ALP) and levels of proinflammatory cytokines (IL-6 and TNF-α) were estimated in serum. Malondialdehyde (MDA), Glutathione (GSH), Superoxide Dismutase (SOD), Catalse (CAT), nucleic acids (DNA and RNA) and Total Proteins (TP) levels were estimated in liver. It has been observed that pretreatment with TQ at 10 and 20 mg kg-1 of body weight respectively could able to minimize the toxicity in compare to ethanol treated group as revealed by the different enzymatic assay in serum. Serum cytokines, IL-6 and TNF-α levels also significantly decreased in TQ treated groups compared to ethanol group. SOD, CAT, GSH, DNA, RNA, TP levels were decreased and MDA was increased significantly in liver of ethanol treated rats. Pretreatment with TQ protected dose dependently the damage-induced in rats liver by the ethanol injection. In conclusion, TQ protects against the liver injury caused by ethanol administration. In view of its antioxidative nature, it may be developed as an effective therapeutic agent for alcohol-induced liver disease by its antioxidative stress and anti-inflammatory features.
PDF Abstract XML References Citation
How to cite this article
Mohammed A. Alsaif, 2007. Effect of Thymoquinone on Ethanol-Induced Hepatotoxicity in Wistar Rats. Journal of Medical Sciences, 7: 1164-1170.
DOI: 10.3923/jms.2007.1164.1170
URL: https://scialert.net/abstract/?doi=jms.2007.1164.1170
DOI: 10.3923/jms.2007.1164.1170
URL: https://scialert.net/abstract/?doi=jms.2007.1164.1170
INTRODUCTION
Alcohol abuse and alcoholism are serious current health and socioeconomic problems throughout the world. Despite great progress made in the field, the development of suitable medications for the treatment of alcoholism remains a challenging goal for alcohol research. Alcoholic Liver Disease (ALD) is the common consequence of prolong and heavy alcohol intake. The fatal changes in the liver include fatty liver, hepatitis and hepatic cirrhosis (Seitz et al., 2005). Ethanol is a fat-soluble non-electrolyte, which is readily absorbed from the gastrointestinal tract, diffuses rapidly into circulation and is distributed uniformly throughout the body (McDonough, 2003). Ethanol is almost exclusively metabolized in the body by enzyme catalyzed oxidative processes. The acetaldehyde formed further oxidized to acetate, which is then converted to carbon dioxide via the citric acid cycle (Albano, 2006). Ethanol can also cause auto-oxidation of the hepatic cells either by acting as a pro-oxidation of the hepatic cells or by reducing the anti-oxidant levels resulting in marked hepatotoxicity (Crawford and Balakenhoan, 1991). Hepatic oxidative stress of the resulting lipid peroxidation has been identified as playing a pathogenic role in alcoholic liver disease (Bunout, 1999).
Thymoquinone active constituent of Nigella sativa is pharmacologically active quinine, which possesses several properties including analgesic and anti-inflammatory actions (Abdel-Fattah et al., 2000; Houghton et al., 1995), protection against chemical induced carcinogenesis (Worthen et al., 1998) and the inhibition of eicosanoids generation (Houghton et al., 1995). Furthermore, it has been reported that TQ prevents oxidative injury in hepatocytes induced by CCl4 or tert-butyl hydroperoxide in various in vitro (Daba and Abdel-Rahman, 1998) and in vivo (Mansour et al., 2001) hepatotoxicity models, as well as acetic acid-induced colitis in rats (Mahgoub, 2003). It has been prevent the membrane lipid peroxidation in hepatocytes (Mansour et al., 2002). In a recent El Gazzar et al. (2006) showed that, TQ attenuated pulmonary inflammation in a mouse model al allergic asthma by decreasing Th2 cytokines and inflammatory cell infiltration in the lung. In another recent study, El Gazzar et al. (2007) reported that, TQ attenuates the proinflammatory response in LPS-stimulated mast cells by modulating nuclear transactivation of NF-κB and TNF-α production.
The protective action of antioxidant is usually due to the inhibition of free radical-induced chain reaction and the resultant prevention of peroxidative deterioration of structural lipids in membranous organelles. Circulating enzymic and non-enzymic antioxidants such as superoxide dismutase, catalase, cytokines and reduced glutathione play an important role in alleviating tissue damage due to the formation of free radicals (Thampi et al., 1991). The present study aims to investigate the protective effect of TQ on toxic damage induced by ethanol in Wistar male rats.
MATERIALS AND METHODS
Animals: The present study was designed and studied during May 2007. Male albino rats, weighing 160-180 g were obtained from Experimental Animal Care Center, College of Pharmacy, King Saud University, Riyadh. They were housed in a temperature controlled room (25±2°C), humidity 50-55% and with 12 h light/dark cycles for at least 1 week. After the stabilization period, rats were randomly divided into 6 groups (six rats in each group): (1) Control (vehicle), (2) Ethanol (7.5 g kg-1), (3) TQ (5 mg kg-1) + Ethanol, (4) TQ (10 mg kg-1) + Ethanol (5) TQ (20 mg kg-1) + Ethanol and Silymarin (10 mg kg-1) + Ethanol. TQ and silymarin were suspended in 0.8% (v/v) Tween 80 and administered orally by using rat feeding needle on every alternate day for 7 days period of study. Animals in groups 1 and 2 were received the same amount of vehicle. The rats in groups 2 to 6 were injected 25% ethanol (7.5 g kg-1 of body weight) intraperitoneally (ip.) 30 min after pretreatment with different doses of TQ, single dose of Silymarin (SL) and vehicle. All rats were kept on free access to standard rats pellet and drinking water.
At the end of the study time, all animals were anaesthetized using ether, blood samples were collected through cardiac puncture. Immediately liver was dissected, dipped in liquid nitrogen for a minute and kept them in deep freezer at -70°C till analysis. Blood samples were centrifuged at 3000 rpm for 10 min, the serum samples were kept in freezer at -20°C till analysis.
Enzymatic assays for serum: Aspartate transminase (AST), Alanine Transminase (ALT), Acid Phosphatase (ACP) and Alkaline Phosphatase (ALP) concentrations were estimated in serum by using commercially available diagnostic kits (Randox diagnostic reagents, Randox Laboratories, USA).
Estimation of TNF-α and interleukin-6 (IL-6) in serum: Serum TNF-α concentration was estimated by using the CYTELISA rat TNF-α obtained from CYTIMMUNE Sciences Inc., Maryland, USA. All samples were assayed in duplicate. The intra assay variation was 6.7%. To avoid inter assay variation all samples were run at one time. Optical density of each well was determined by using a microplate reader (Thermo Labsystems, Finland).
Serum IL-6 was estimated by using the ELISA Quantikine rat IL-6 immunoassay kit obtained from R and D Systems Inc., Minneapolis, MN, USA. The samples were assayed in duplicate. The intra assay variation was 5.5%. To avoid inter assay variation all samples were run at one time. Optical density of each well was determined by using a microplate reader (Thermo Labsystems, Finland).
Estimation of MDA in liver: The method described by Ohkawa et al. (1979) was used. Malondialdehyde (MDA) was measured as an indicator of lipid peroxidation. Liver tissues were homogenized in KCl solution and incubated with thiobarbituric acid. After centrifugation the pink clear layer was read at 532 nm. Malondialdehyde bis (dimethyl acetal) was used as an external standard.
Estimation of GSH in liver: Glutathione concentration was assayed using the method of Sedlak and Lindsay (1968). A cross sectional piece of liver tissues (200 mg) were dissected and homogenized in ice-cold 0.02 M ethylenediaminetetraacetic acid (EDTA). Aliquots of 0.5 mL of the tissue homogenates were mixed with 0.2 M Tris buffer, pH 8.2 and 0.1 mL of 0.01 M Ellman’s reagent, [5,5’-dithiobis-(2-nitro-benzoic acid)] (DTNB). Tubes were centrifuged at 3000 g at room temperature for 15 min. The absorbance of the clear supernatants was read in a spectrophotometer at 412 mμ in 1 cm quarts cells. The concentrations were estimated by using the standard curve.
Estimation of SOD activity in liver: Superoxide dismutase activity in liver was assayed spectrophotometerically (560 nm) by the method described by Kakkar et al. (1984) using nitroblue tetrazolium as the indicator reagent.
Assay procedure for catalase (CAT) activity in liver: The catalase activity was measured by the method of Abei (1988) using hydrogen peroxide as substrate. The disappearance of H2O2 was followed at 240 nm. The activity was expressed as μmole-1 min-1 mg-1 protein using the extension coefficient of 0.0436 mM-1 mg-1.
Estimation of total proteins and nucleic acids: Total proteins were estimated by the modified Lowry method of Schacterle and Pollack (1973). Bovine serum albumin was used as standard. The method described by Bregman (1983) was used to determine the levels of nucleic acids. Tissues were homogenized and the homogenate was suspended in ice-cold Trichloroacetic Acid (TCA). After centrifugation, the pellet was extracted with ethanol. DNA was determined by treating the nucleic acid extract with diphenylamine reagent and reading the intensity of blue color at 600 nm. For quantification of RNA, the nucleic acid extract was treated with orcinol and the green colour was read at 660 nm. Standard curves were used to determine the amounts of nucleic acids present.
Statistical analysis: All data are presented as the mean±SE The data were evaluated by a one-way ANOVA using SPSS program and the differences between the means assessed using Student’s t-test.
RESULTS
Ethanol treatment to control rats caused significant increase in AST, ALT, ALP and ACP activities compared the control group of rats. Pretreatment of rats with TQ showed significant protective activity in AST, ALT and ACP at higher doses (10 and 20 mg kg-1) and in ALP only at high (20 mg kg-1) compared to the ethanol supplemented rats. SL treatment (10 mg kg-1) also showed protection against the enzymatic activities induced by ethanol treatment (Table 1).
Both the cytokines, TNF-α and IL-6 levels in serum (pg mL-1) were found significantly higher (p<0.001) in ethanol supplemented rats compared to the experimental control rats. Such increased levels of TNF-α and IL-6 were significantly restored dose dependent manner in rats treated with TQ. Silymarin (10 mg kg-1) treatment showed significant protection against the elevated TNF-α and IL-6 levels induced by ethanol challenge (Table 2).
| Table 1: | Effect of THQ on different biochemical parameters in serum of rats |
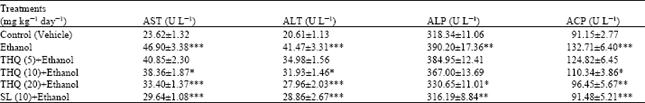 | |
| Values are expressed in Mean±SE of six animals in each group. Statistical analysis was done by ANOVA, *: p<0.05, **: p<0.01 and ***: p<0.001 Student’s t-test, Ethanol alone group was compared with control group, Drugs pretreated groups were compared with ethanol alone group | |
| Table 2: | Effect of THQ on serum TNF-alpha and IL-6 levels of rats |
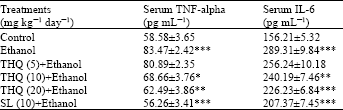 | |
| Values are expressed in Mean±SE of 6 animals in each group. Statistical analysis was done by ANOVA; *: p<0.05, **: p<0.01 and ***: p<0.001 Student’s t-test, Ethanol alone group was compared with control group. Drugs pretreated groups were compared with ethanol alone group | |
| Table 3: | Effect of THQ on hepatic GSH and MDA concentrations of rats |
 | |
| Values are expressed in Mean±SE of 6 animals in each group. Statistical analysis was done by ANOVA; *: p<0.05, **: p<0.01 and ***: p<0.001 Student’s t-test, Ethanol alone group was compared with control group. Drugs pretreated groups were compared with ethanol alone group | |
| Table 4: | Effect of THQ on hepatic SOD and CAT concentrations of rats |
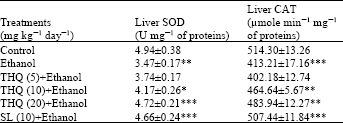 | |
| Values are expressed in Mean±SE of 6 animals in each group. Statistical analysis was done by ANOVA; *: p<0.05, **: p<0.01 and ***: p<0.001 Student’s t-test, Ethanol alone group was compared with control group. Drugs pretreated groups were compared with ethanol alone group | |
Ethanol-induced significant decrease the hepatic GSH and increase hepatic MDA concentrations as compared to experimental controls. Pretreatment with TQ protected the GSH decrease and MDA increase induced by ethanol as compared to ethanol alone group in dose dependent manner. Silymarin taken as reference drug also showed protection against the changes in induced by ethanol in hepatic GSH and MDA levels (Table 3).
Single dose of ethanol administration to rats induced significant decrease in hepatic SOD and CAT activities as compared to control group of rats. Pretreatment with TQ to ethanol administered rats showed significant increase dose dependent manner in hepatic SOD and CAT activities as compared to the rats only treated with alcohol (Table 4).
Ethanol supplementation induced significant decrease in liver RNA, DNA and protein concentrations as compared to the control group of rats. Pretreatment with TQ showed protection against the decreased levels of RNA, DNA and total proteins by alcohol administration (Table 5).
| Table 5: | Effect of THQ on hepatic RNA, DAN and total protein concentrations of rats |
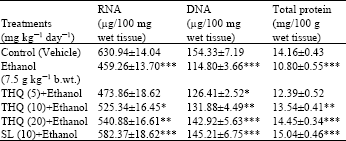 | |
| Values are expressed in Mean±SE of 6 animals in each group. Statistical analysis was done by ANOVA; *: p<0.05, **: p<0.01 and ***: p<0.001 Student’s t-test, Ethanol alone group was compared with control group. Drugs pretreated groups were compared with ethanol alone group | |
DISCUSSION
Oxidative stress is one major factor in etiology of ethanol injury, mainly by Kupffer cell derived Reactive Oxygen Species (ROS) (Vrba and Modriansky, 2002; Wheeler, 2003). Ethanol activates Kupffer cells primarily through the action of a substance called endotoxin, which is released by certain gram-negative bacteria present in the intestine (Keshavarzian et al., 2001). Kupffer cell activation generates ROS and pro-inflammatory TNF-α, it can lead to liver damage (Hoek and Pastorino, 2002; Zhou et al., 2003).
Ethanol causes injury to the liver cells and the activities of AST and ALT are the most sensitive test employed in the diagnosis of hepatic disease (Chenoweth and Hake, 1962) and were increased in ethanol treated rats. Pretreatment with TQ to ethanol treated rats decreased AST and ALT activities. Present results are in agreement with Daba and Abdel-Rahman (1998) showed TQ protection against tert-butyl hydroperoxide-induced hepatotoxicity in rats isolated hepatocytes evidenced by decreased leakage of ALT and AST. This shows TQ can preserve the structural integrity of the liver from the adverse effects of ethanol. Administration of a single dose of ethanol significantly elevated the serum ACP and ALP activities compared to the normal animals. This indicated necrosis of hepatocytes that results in the elevation of serum ACP and ALP from a possible cholistasis (Ajith et al., 2007). Pretreatment with TQ showed significant decrease in ACP and ALP levels as compared the animals administered ethanol only.
Oxidative stress has been implicated in the process of alcohol-induced TNF-α production (Zhou et al., 2003, 2004). In the present study, ethanol administration significantly raised both serum IL-6 and TNF-α levels together this confirmed that local inflammatory changes and release of pro-inflammatory cytokines did occur in our experimental model. Pretreatment with TQ showed significant decrease in ethanol-induced elevation of serum IL-6 and TNF-α level. El-Mahmoundy et al. (2005) reported that TQ administered intraperitonially in diabetic rats at 3 mg kg-1 or added to cultured macrophages at 10 μM normalized the elevated levels of the proinflammatory cytokines IL-1β and TNF-α. TQ has been shown to inhibit generation of the inflammatory mediators, thromboxane B2 and leukotriene B4, by targeting the cyclooxygenase and 5-lipoxygenase pathways of Arachidonic acid metabolism (Houghton et al., 1995).
Ethanol is capable of generating oxygen radicals, inhibiting GSH synthesis, producing GSH loss from the tissue, increasing MDA levels and impairing antioxidative defense systems (Genc et al., 1998; Pramyothin et al., 2006). It has been shown that marked decreases in the glutathione pool occur in liver after acute ethanol intoxications (Guerri and Grisolia, 1980), which is partly due to the binding of acetaldehyde with cysteine and/or glutathione (Lieber, 1994). Furthermore, severe GSH depletion is known to associate with lipid peroxidation (Videla et al., 1980). Present results are consisted by acute ethanol treatment (Speisky et al., 1985; Chen et al., 1992) and the administering TQ to alcohol treated rats helped to restore the GSH levels to near those of the experimental control rats. In the present study, MDA formation, the index of lipid peroxidation, was significantly increased after ethanol treatment and TQ supplementation was potentially effective in reducing lipid peroxidation, conform its antioxidant nature.
Natural or synthetic compounds with antioxidant properties may help to alleviate the liver damage totally of partially. Superoxide ion (O2¯) and hydroxyl radical are known to cause marked injuries to the surrounding tissues and organs (Reedy and Lokesh, 1992). Removing superoxide ion and hydroxyl radical is probably one of the most effective defense mechanisms against a variety of diseases (Sheela and Angusti, 1995). Activities of SOD and CAT help to scavenge superoxide ions and hydroxyl ions, respectively, were significantly lower in alcohol-treated rats as compared with those of control rats. Lowered activities of SOD and CAT will result in the accumulation of these highly reactive free radicals leading to deleterious effects such as loss of cell membrane integrity and membrane function (Reedy and Lokesh, 1992). This decrease in the SOD activity may be associated with the elevation of the intracellular concentration of H2O2 as a result of CAT inactivation (Antonekov and Panchenko, 1988). CAT has been reported to be responsible for the detoxification of H2O2, which is an effective inhibitor of SOD (De duve and Baudhhuin, 1996). The observed restoration of the SOD and CAT activities in the present study on treatment with TQ may be due to the direct stimulatory effect of TQ on SOD and CAT. Furthermore, the decrease in the tissue lipid peroxidation on TQ treatment can also be correlated with the elevated SOD and CAT activities.
In present study, hepatic nucleic acids levels depleted significantly in ethanol treated rats. Pretreatment with TQ and SL showed significant protection against decreased hepatic nucleic acids levels by the ethanol administrations. Several studies reported that, toxic agents such as chemotherapy drugs, carcinogens, reactive oxygen and nitrogen species cause damages in DNA and RNA molecules (Bohr, 2002; Moreira et al., 2005; Ungvari et al., 2005; Kang and Hamasaki, 2005). Wieland and Lauterburg (1995) reported that oxidation of DNA and mitochondrial proteins following ethanol administration resulted in permanent damage of the liver in alcoholic subjects. In animal studies, Abou-Seif et al. (2003) and El-Missiery et al. (2007) reported that, radiation-induced oxidative stress caused significant damage in hepatic RNA concentrations. It is well documented that green tea is an efficient radical scavenger, activates enzymes important for detoxification and prevent in vivo DNA damage and lipid peroxidation (Cheng et al., 1991). Present results indeed support such notion because TQ supplementation could effectively reduce ethanol-induced increase in DNA, RNA and total protein damage.
The results of the present study concluded that TQ significantly and dose dependently prevented the ethanol-induced acute hepatotoxicity by enhancing the hepatic antioxidant activity. However, further detailed studies are required to establish its clinical application.
REFERENCES
- Abdel-Fattah, A.F.M., K. Matsumoto and H. Watanabe, 2000. Antinociceptive effects of Nigella sativa oil and its major component, thymoquinone, in mice. Eur. J. Pharmacol., 400: 89-97.
CrossRefDirect Link - Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Abou-Seif, M.A.M., M.M. El-Naggar, M. El-Far, M. Ramadan and N. Salah, 2003. Amelioration of radiation-induced oxidative stress and biochemical alteration by SOD model compounds in pre-treated γ-irradiated rats. Clin. Chem. Acta, 337: 23-33.
Direct Link - Ajith, T.A., U. Hema and M.S. Aswathy, 2007. Zingiber officinale roscoe prevents acetaminophen-induced acute hepatotoxicity by enhancing hepatic antioxidant status. Food Chem. Toxicol., 45: 2267-2272.
Direct Link - Albano, E., 2006. Alcohol, oxidative stress and free radical damage. Proc. Nutr. Soc., 65: 278-290.
CrossRefDirect Link - Antonenkov, V.D. and L.F. Panchenko, 1988. Effect of chronic ethanol treatment under partial catalase inhibition on the activity of enzymes related to peroxide metabolism in rat liver and heart. Int. J. Biochem., 20: 823-828.
CrossRef - Bohr, V.A., 2002. DNA damage and its processing: Relation to human disease. J. Inherit. Metab. Dis., 25: 215-222.
Direct Link - Bunout, D., 1999. Nutritional and metabolic effects of alcoholism: Their relationship with alcoholic liver disease. Nutrition, 15: 583-589.
CrossRefDirect Link - Chen, L.H., N. Hu and T.L. Huang, 1992. Effects of acute alcohol intoxication on liver antioxidant defense systems in rats. Biochem. Arch., 8: 95-100.
Direct Link - Crawford, D.W. and D.H. Balakenhoan, 1991. Arterial wall oxygenation oxy radicals and atherosclerosis. Atherosclerosis, 89: 97-108.
Direct Link - Daba, M.H. and M.S. Abdel-Rahman, 1998. Hepatoprotective activity of thymoquinone in isolated rat hepatocytes. Toxicol. Lett., 95: 23-29.
CrossRefDirect Link - De Duve, C. and P. Baudhuin, 1996. Peroxisomes (microbodies and related particles). Physiol. Rev., 46: 323-357.
PubMedDirect Link - El Gazzar, M., R. El Mezayen, J.C. Marecki, M.R. Nicolls, A. Canastar and S.C. Dreskin, 2006. Anti-inflammatory effect of thymoquinone in a mouse model of allergic lung inflammation. Int. Immunopharmacol., 6: 1135-1142.
CrossRefDirect Link - El-Gazzar, M., R. El-Mezayen, J.C. Marecki, M.R. Nicolls and S.C. Dreskin, 2007. Thymoquinone attenuates proinflammatory responses in lipopolysaccharide-activated mast cells by modulating NF-kappaB nuclear transactivation. Biochim. Biophys. Acta, 1770: 556-564.
Direct Link - El-Mahmoundy, A., Y. Shimizu, T. Shiina, H. Matsuyama, M. El-Sayed and T. Takewaki, 2005. Successful abrogation by thymoquinone against induction of diabetes mellitus with streptozotocin via nitric oxide inhibitory mechanism. Int. Immunopharmacol., 5: 195-207.
CrossRefPubMedDirect Link - El-Missiry, M.A., T.A. Fayed, M.R. El-Sawy and A.A. El-Sayed, 2007. Ameliorative effect of melatonin against gamma-irradiation-induced oxidative stress and tissue injury. Ecotoxicol. Environ. Saf., 66: 278-286.
CrossRefDirect Link - Genc, S., F. Gurdol, Y. Oner-Iyidogan and I. Onaran, 1998. The effect of melatonin administration on ethanol-induced lipid peroxidation in rats. Pharmacol. Res., 37: 37-40.
Direct Link - Guerri, C. and S. Grisolia, 1980. Changes in glutathione in acute and chronic alcohol intoxication. Pharmacol. Biochem. Behavior., 13: 53-61.
CrossRefDirect Link - Hoek, J.B. and J.G. Pastorino, 2002. Ethanol, oxidative stress and cytokine-induced liver cell injury. Alcohol, 27: 63-68.
Direct Link - Houghton, P.J., R. Zarka, B. de las Heras and J.R.S. Hoult, 1995. Fixed oil of Nigella sativa and derived thymoquinone inhibit eicosanoid generation in leukocytes and membrane lipid peroxidation. Planta Med., 61: 33-36.
CrossRefPubMedDirect Link - Kakkar, P., B. Das and P.N. Viswanathan, 1984. A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem. Biophys., 21: 130-132.
PubMedDirect Link - Kang, K. and N. Hamasaki, 2005. Alterations of mitochondrial DNA in common diseases and disease states: Neurodegeneration, heart failure, diabetes and cancer. Curr. Med. Chem., 12: 429-441.
Direct Link - Keshavarzian, A., S. Choudhary, E.W. Holmes, S. Young, A. Banan and S. Jakate, 2001. Preventing gut leakiness by oats supplementation ameliorates alcohol-induced liver damage in rats. J. Pharmacol. Exp. Ther., 299: 442-448.
Direct Link - Lieber, C.S., 1994. Mechanisms of ethanol-drug-nutrition interactions. Clin. Toxicol., 32: 631-681.
Direct Link - Mahgoub, A.A., 2003. Thymoquinone protects against experimental colitis in rats. Toxicol. Lett., 143: 133-143.
PubMed - Mansour, M.A., O.T. Ginawi, T. El-Hadiyah, A.S. El-Khatib, O.A. Al-Shabanah and H.A. Al-Sawaf, 2001. Effects of volatile oil constituents of Nigella sativa on carbon tetrachloride-induced hypatotoxicity in mice: Evidence for antioxidant effects of thymoquinone. Res. Commun. Mol. Pathol. Pharmacol., 110: 239-251.
Direct Link - Mansour, M.A., M.N. Nagi, A.S. El-Khatib and A.M. Al-Bekairi, 2002. Effects of thymoquinone on antioxidant enzyme activities, lipid peroxidation and DT-diaphorase in different tissues of mice: A possible mechanism of action. Cell Biochem. Funct., 20: 143-151.
CrossRefPubMedDirect Link - McDonough, K.H., 2003. Antioxidant nutrients and alcohol. Toxicology, 189: 89-97.
CrossRefDirect Link - Moreira, P.I., M.A. Smith, X. Shu, A. Nunomura, R.J. Castellani and G. Perry, 2005. Oxidative stress and neurodegeneration. Ann. Acad. Sci. N.Y., 1043: 545-552.
Direct Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Pramyothin, P., P. Samosorn, S. Poungshompoo and C. Chaichantipyuth, 2006. The protective effects of Phyllanthus emblica Linn. extract on ethanol induced rat hepatic injury. J. Ethnopharmacol., 107: 361-364.
Direct Link - Reedy, A.C. and B.R. Lokesh, 1992. Studies on apice principle as an antioxidant in the inhibition of lipid peroxidation of rat liver microsomes. Mol. Cell Biochem., 111: 117-124.
Direct Link - Schacterle, G.R. and R.L. Pollack, 1973. A simplified method for quantitative assay for small amount of proteins in biological materials. Anal. Biochem., 51: 654-655.
PubMed - Sedlak, J. and R.H. Lindsay, 1968. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman's reagent. Anal. Biochem., 25: 192-205.
CrossRefPubMedDirect Link - Seitz, H.K., C.S. Lieber, F. Stickel, M. Salaspuro, H.P. Schlemmer and Y. Horie, 2005. Alcoholic liver disease: From pathophysiology to therapy. Alcohol. Clin. Exp. Res., 29: 1276-1281.
CrossRefDirect Link - Sheela, C.G. and K.T. Angusti, 1995. Antiperoxide effects of S-allyl cystein sulphoxide isolated from Allium sativum Linn. and gugulipid in cholesterol diet fed rats. Indian J. Exp. Biol., 33: 337-341.
Direct Link - Speisky, H., A. MacDonald, G. Giles, H. Orrego and Y. Israel, 1985. Increased loss and decreased synthesis of hepatic glutathione after acute ethanol administration. Biochem. J., 225: 565-572.
PubMed - Thampi, H.B.S., G. Manoj, S. Leelamma and V.P. Menon, 1991. Dietary fiber and lipid peroxidation. Effects of dietary fiber on levels of lipids and lipid peroxides in high fat diet. Indian J. Exp. Biol., 29: 563-567.
Direct Link - Ungvari, S., S.A. Gupte, R.A. Recchia, S. Batkai and P. Pacher, 2005. Role of oxidative-nitrosative stress and downstream pathways in various forms of cardiomyopathy and heart failure. Curr. Vasc. Pharmacol., 3: 221-229.
Direct Link - Vrba, J. and M. Modriansky, 2002. Oxidative burst of Kupffer cells: Target for liver injury treatment. Biomed. Pap., 146: 15-20.
PubMedDirect Link - Wheeler, M.D., 2003. Endotoxin and Kupffer cell activation in alcoholic liver disease. Alcohol Res. Health, 27: 300-306.
Direct Link - Wieland, P. and B.H. Lauterburg, 1995. Oxidation of mitochondrial proteins and DNA following administration of ethanol. Biochem. Biophys. Res. Commun., 213: 815-819.
Direct Link - Worthen, D.R., O.A. Ghosheh and P.A. Crooks, 1998. The in vitro anti-tumor activity of some crude and purified components of blackseed, Nigella sativa L. Anticancer Res., 18: 1527-1532.
PubMedDirect Link - Zhou, Z., L. Wang, Z. Song, J.C. Lambert, C.J. McClain and Y.J. Kang, 2003. A critical involvement of oxidative stress in acute alcohol-induced hepatic TNF-α production. Am. J. Pathol., 136: 1137-1144.
Direct Link