
Delelegn Woyessa
Department of Biology, Jimma University, College of Natural Sciences, P.O. Box 378, Jimma, Ethiopia
Fassil Assefa
Department of Biology, Addis Ababa University, P.O. Box 1176, Addis Ababa, Ethiopia
Research Journal of Microbiology
Year: 2011 | Volume: 6 | Issue: 4 | Page No.: 343-355







ABSTRACT
The aim of this study was to assess the effect of Plant Growth Promoting Rhizobacteria (PGPR) on growth and yield of tef. About 160 bacteria were isolated from rhizosphere of tef (Eragrostis tef Zucc. Trotter) and characterized on the basis of PGPR and biochemical features. Four best isolates (AURB15, AURB34, AURB56 and AURB65) were selected and further characterized by physiological methods and identified to species level using API kit method. The selected isolates were then evaluated by inoculation on to two tef varieties (DZ-Cr-37 and DZ-01-974) at greenhouse condition. The isolates displayed diverse morphological, physiological, biochemical and PGPR characteristics. They showed growth in wide range of temperature, pH and NaCl. The isolates were found to belong to three genera and four species. Isolate AURB15 (Pseudomonas fluorscens) was observed to significantly increase mean root dry weight (39%), root shoot ratio (42%) and yield (45%) of the tef variety DZ-Cr-37; like wise isolate AURB65 (Bacillus subtilis) better increased mean root dry weight (28%), root shoot ratio (19%) and yield (44%) of tef variety DZ-01-974. Comparable mean shoot dry weight, % nitrogen and total protein (p>0.05) was recorded by inoculation of the isolates. Where as inoculation of two isolates, AURB34 (Burkholderia cepacia) and AURB56 (Bacillus coagulans) on DZ-01-974 did not increase mean root dry weight, root shoot ratio and yield of DZ-01-974. The increase in mean root dry weight, root shoot ratio and yield of tef varieties DZ-Cr-37 and DZ-01-974 by AURB15 (Pseudomonas fluorscens) and AURB65 (Bacillus subtilis) suggests that the isolates have better potential for further field testing and application in improving yield of the tef varieties.
PDF Abstract XML References Citation
Received: November 28, 2010;
Accepted: January 15, 2011;
Published: February 26, 2011
How to cite this article
Delelegn Woyessa and Fassil Assefa, 2011. Effects of Plant Growth Promoting Rhizobaceria on Growth and Yield of Tef (Eragrostis tef Zucc. Trotter) under Greenhouse Condition. Research Journal of Microbiology, 6: 343-355.
URL: https://scialert.net/abstract/?doi=jm.2011.343.355
URL: https://scialert.net/abstract/?doi=jm.2011.343.355
INTRODUCTION
Tef (Eragrostis tef Zucc. Trotter) is an important staple food and feed crop of Ethiopia and grown on about 2 million hectares of land covering about 26% of the whole area cultivated to annual field crops (CSA, 2004). According to Ethiopian Agricultural Research Organization (EARO, 2000), the nutritional status of the crop is comparable to other major world cereals and it provides about two-thirds of the daily protein intake of most Ethiopians. Tef has been of high demand by the majority of the nation’s population because of its superior qualities and is grown predominantly for human consumption. It has also other multiple uses including as reinforcement for mud bricks. Tef grain is one of the most expensive in Ethiopia to purchase as it requires labor-intensive harvesting and processing techniques and produces especially low yield despite its coverage of the greatest land space (Belay et al., 2008).
Many of the hitherto attempts to improve tef yield and performance through breeding and/or genetic manipulation could not drastically change the low yield of tef. Though tef has been cultivated on about 30% of the cereal land in the country, it contributes only about 22% of the annual grain yield (0.9 tha-1) production compared to other cereals (Tefera et al., 2003). Furthermore, the ever-increasing cost of agricultural inputs is also becoming neither available nor affordable to most Ethiopian poor farmers that are struggling to improve food security and their living standard.
The human population increase coupled with solid consumer demand is currently responsible for a soaring price of tef making it the most expensive cereal in the country. Such a demand together with low productivity and increasing need for chemical fertilizers necessitate the need to look into additional, less expensive and environmentally friendly input to improve tef production.
Microorganisms are important components of soil that influence plant growth directly or indirectly by their beneficial or detrimental activities (Deepa et al., 2010). Among the beneficial effects of soil microorganisms are nitrogen fixation, denitrification and phytohormone production (Dobbelaere et al., 2003; Glick et al., 2007; Jalili et al., 2009; Babalola, 2010), enhanced nutrient mobility, pathogen control and stress alleviation (Glick et al., 2007; Jalili et al., 2009; Babalola, 2010) resulting in improved growth and plant yield (Babalola, 2010). The proper use of phytobeneficial soil microorganisms as agricultural inputs for improved crop production requires isolation, identification and selection of rhizosphere-competent microorganisms with plant growth-promoting attributes (Lucy et al., 2004; Hynes et al., 2008). Application of PGPR to soil has also been reported to improve soil structure with no leaving soil toxic effects in contrast to continuous use of chemical fertilizers (Babalola, 2010). On the other hand, though the use of chemical fertilizers can generally improve crop production, other drawbacks of such chemicals particularly in causing poor microbial diversity and problems in the long term sustainability have also been reported (Nakhro and Dkhor, 2010).
Isolation and identification of rhizosphere microorganisms such as Pseudomonas and Bacillus for proper utilization of their beneficial effects as plant growth promoters on maize, wheat, rye and other crops have been well reported (Lucy et al., 2004). In Ethiopia, recent studies on tef root-associated microorganisms are scarce except very few reports such as the effect of Azosprillum bacterial isolates on the growth and nitrogen content of tef by Zewdie et al. (2000). The results indicated that inoculation of Azosprillum strains resulted in increased height, shoot dry matter and mean grain yield and nitrogen content. However, there is still a dearth of information on the role of PGPR such as Bacillus and Pseudomonas on tef growth and production. Therefore, this necessitates the study of more PGPR with important traits from the roots of tef plant to serve as potential commercial inoculants to boost tef production. Therefore, the objectives of this study were: (1) To isolate and screen PGPR characteristics of tef root associated bacteria and (2) to evaluate the effect of selected isolates on various growth performance and nitrogen content of two tef varieties on pot experiment under greenhouse condition.
MATERIALS AND METHODS
Sampling sites and collection of samples: The study was conducted from August 2006 to July 2007 at Addis Ababa University, Department of Biology, in the Applied Microbiology Laboratory and Greenhouse conditions.
Soil samples were collected from two well-known tef-growing areas of agricultural fields of Alemgena and Bushoftu (about 15 km Southwest and 40 km east of Addis Ababa, respectively). Soil samples with intact tef roots were collected from tef rhizospheres in sterile plastic bags as described by Oyeyiola (2009) after removing debris of the topsoil and the top 6-10 cm soil. The soil samples were immediately transported to the Addis Ababa University, Applied Microbiology laboratory for further study.
Media preparation and isolation of culture: Soils from the rhizosphere of tef were carefully removed and bulked together. The layer particles were removed using 2 mm sieve. The roots were then thoroughly washed with sterile distilled water to remove as much PGPR as possible. A series of dilutions (10-2, 10-3, 10-4, 10-5 and 10-6) were made following standard procedures. From dilutions of 10-4 and 10-5, 0.1 mL of suspensions were transferred to nutrient agar plates (9.6-9.8 cm dm) and uniformly spread using bent glass rod. The plates were then incubated at 30±2°C for 48 h.
Representative colonies of all morphological types were randomly picked and subcultured on nutrient agar media for purification. The purified cultures were maintained on nutrient agar slant at 4°C and preserved in 80% glycerol at -22°C for subsequent experiments and all the cultures were checked for purity for each experiment.
Screening of the isolates
Screening phosphate solubilizing isolates: All the isolates were tested in vitro for their phosphate solubilization activity following the method described by Donate-Correa et al. (2005) using Pikovskaya agar medium containing the following ingredients ( g L-1 ): Glucose (5), Ca3(PO4)2 (5), (NH4)2SO4 (0.5), Yeast Extract (0.5), MgSO4.7H2O (0.2), NaCl (0.1), MnSO4 (0.002), FeSO4 (0.002) and Agar (15). The medium was autoclaved at 121°C for 15 min. About 20 mL of the molten agar medium was poured into each petridish and allowed to solidify before inoculating the isolates. A 24 h broth culture was spot inoculated on the petridishes in triplicate using sterile loop and incubated at 30±2°C for 5-7 days. Bacterial colonies that formed clear zones (haloes) were considered as phosphate solubilizers and clear zone diameters were measured.
Screening for cyanide production: Hydrogen cyanide (HCN) production from glycine was tested following the procedure of Donate-Correa et al. (2005). Tryptic Soya Agar (TSA) supplemented with glycine (4.4 g L-1) was prepared and autoclaved for 15 min at 121°C and poured on petridishes (9.2-9.8 cm diameter). The medium was maintained in a refrigerator overnight and allowed to be at room temperature for about 30 min before inoculation. A loop full of 24 h old broth culture of the isolates were inoculated to the plates. Sterile filter paper strips soaked with 0.5% (w/v) picric acid were fixed to the underside of the petridish lids. The plates were then sealed with parafilm and incubated for 5-7 days at 30°C. An uninoculated plate and a plate without picric acid impregnated papers were used as controls. A change in the color of the filter strips from yellow to brown or reddish-brown was regarded as indication of cyanogenic potential.
Screening for antimicrobial production: This test was done using a modified method of Brinkhoff et al. (2004). A reference test organism, E. coli ATCC 25922 that was kept at -22°C in tryptic soya broth supplemented with 80% glycerol was brought to Applied Microbiology laboratory, Addis Ababa University from the Ethiopian Health and Nutrition Research Institute (EHNRI) and kept at -22°C. A nutrient broth 8.0 (g L-1) and NaCl, 5.0 (g L-1) was prepared in two 250 mL flasks and autoclaved for 15 min at 121°C. One of the flasks was used for control (blank) to adjust optical density (OD) reading and the other was used for inoculation of the test organism. A sterile loopful (about 106 cfu mL-1) of the test organism was inoculated to the flask and incubated at 37°C in a Gallenkamp orbital shaker at 120 rpm until the OD600 nm reached 1.3 (the control flask was kept at 4°C and used for calibration of the spectrophotometer.
All isolates to be tested for antibiotic production were separately incubated up to their stationary phases (36-48 h) at 30°C in a sterile 5 mL nutrient broth. Plates with sterile medium containing nutrient broth in g L-1 (8.0), NaCl (5.0) and agar (15.0) were prepared and overlaid with 0.7% top agar containing 1% (50 μL test organism 5 mL-1 top agar) cell suspension of the E. coli test organism. Sterile blank antibiotic discs (Aldrich Chem. Co., USA) (diameter, 6 mm) were soaked with stationary phase culture broth of the soil isolates to be tested for antibiotic production and put on the plates at four corners and the control at the center. The control was not soaked with any culture. The plates were then incubated at 37°C for 3-5 days to check for inhibition zones.
Characterization and identification of isolates: The isolates were characterized for Gram-reaction, shape, spore production, catalase and oxidize tests as well as tolerance of different levels of temperature, pH and salt concentration. The isolates were then bio-chemically identified to species and sub species level according to Analytical Profiles Index, API 20 NE and API 50 CHB test (bio-Mérieux, Marcy, I’tole, France) for Gram-negatives and Gram-positives, respectively following the manufacturer’s instructions. The result was finally analyzed using API software package (APIWeb, Version-1.1.0).
Effect of PGPR inoculation on growth and yield of tef
Soil and seed sample preparation: A soil sample of about 150 kg was collected from local farm plots of Sebeta (25 km south west of Addis Ababa), where tef was produced for the past several years. The soil was sieved in 2 mm sieve and mixed with thoroughly washed and dried sand in a proportion of 3:1 (soil: sand). The soil and sand mixture was then filled into thirty 3 capacity plastic pots (6.9x6.5 cm) that were surface-sterilized with 95% ethanol. A hand full of two tef varieties (DZ-Cr-37 and DZ-01-974) were surface sterilized with 0.5% (w/v) HgCl2 for two minutes at room temperature and thoroughly washed with sterile distilled water (Siriskandarajah et al., 1993). Ten seeds from each variety were then sown into each of the15 pots.
Selection of isolates for greenhouse evaluation: Based on three PGPR properties (P-solubilization, antibiotic production and HCN) production during the laboratory experiment, four isolates were selected for greenhouse evaluation. Consequently, AURB15 (Psudomonas fluorscens), AURB34 (Burkholderia cepacia), AURB56 (Bacillus coagulans) and AURB65 (Bacillus subtilis) were taken for the inoculation experiment as each of these isolates showed at least two of the tested PGPR properties.
Inoculum preparation: Nutrient agar slant cultures of the selected isolates maintained at 4°C were used as stock cultures for inoculant preparation. A loop full of the stock culture was separately inoculated into nutrient broth tubes and incubated at 30±2°C for 24 h. A loop full of the broth culture was then separately streaked on to nutrient agar plates to check for purity. A single colony from each isolate was then inoculated in to nutrient broth tubes and incubated at 30±2°C for 24 h, from which a volume of 1 mL was inoculated into 250 mL Erlenmeyer flasks containing 150 mL nutrient broth. The flasks were incubated at 30°C on a Gallenkamp orbital shaker up to the cell concentration of 107-108 cfu mL-1 (OD 600 nm = 1.0) (Pillai and Swarup, 2002).
Greenhouse pot experiment: The pot experiment was carried out under greenhouse condition at the Science Faculty Campus from September to December 2006. Mean minimum and maximum temperatures inside the greenhouse during the study period were 13 and 35°C, respectively. Seeds were inoculated with one of the selected isolates at the time of sowing at a rate of 107-108 (1.0 mL) bacterial cells per seed for every treatment with a sterile 1.0 mL disposable syringe (Kucey, 1988). Inoculation was also done using the same procedure fifteen days after planting. Control pots were inoculated similarly with equal amount of sterile distilled water. The seed inoculated and control pots were laid out in a Randomized Complete Block Design (RCBD) with two tef verities and four bacterial isolates and with three replications of the following ten treatments:
| • | AURB15xDZ-Cr-37 |
| • | AURB15xDZ-01-974 |
| • | AURB56xDZ-Cr-37 |
| • | AURB56xDZ-01-974 |
| • | AURB34xDZ-Cr-37 |
| • | AURB34xDZ-01-974 |
| • | AURB65xDZ-Cr-37 |
| • | AURB65xDZ-01-974 |
| • | UninoculatedxDZ-Cr-37 |
| • | UninoculatedxDZ-01-974 |
All pots were watered regularly with sterile distilled water. The number of seedlings in each pot was thinned down to four after ten days of emergence and the plants were grown for 75 days.
Growth parameters and measurement of tef yield: After full maturity (75 days of growth), plant height and number of tillers of each plant were recorded before harvesting. Following harvest, the root mass of plants was carefully dug out from the soil and washed to remove adhering soil particles. Passing the water through 0.5 mm sieve retrieved fine root tips. The harvested plant was then sun-dried to constant weight in the greenhouse for 5 days. The root and shoot biomass (g) was then separately weighed and recorded using Mettler E2000 balance (Heiligenstadt, Germany). Grain yield per plant was also recorded after hand threshing the shoot part of each plant.
Determination of nitrogen content and crude protein of tef grains: Total tef grain nitrogen determination was done using Kjeldahl method for soil and plant analysis as described by Sertsu and Bekele (2000). Dried grain samples from each pot were milled into powder using a coffee grinder and sieved through 0.5 mm sieve. From each sample 0.3 g sub sample was weighed and subjected to Kjeldahl digestion for the determination of total nitrogen in samples. The grain protein was calculated by multiplying the total grain nitrogen by a conversion factor 6.25 as recommended by Horowitz (1975) for estimation of protein content of plant tissues.
Data analysis: The data for plant growth parameters were analyzed using SAS computer software Ver. 6.12 (SAS, 1996) after checking normality for each variable according to Gomez and Gomez (1984). Duncan’s multiple range test (DMRT) was used for mean separation.
RESULTS
Screening, characterization and identification of PGPR from tef rhizosphere.
Morphological and cultural characteristics: Of about 160 bacterial isolates from tef rhizosphere, four isolates were selected based on screening result on P-solublization, antimicrobial property and hydrogen cyanide potential. The cellular and morphological characterization also resulted in noticeable diversity (Table 1).
Screening of isolates based on PGPR and biochemical properties: Based on screening for three PGPR properties (P-solublization, antimicrobial property and HCN production, four isolates were selected as best and further characterized by different biochemical tests (Table 2).
Identification of the isolates based on API kit test: The four isolates were identified up to species levels by further screening and re-screening through several biochemical tests and using API (20NE and 50 CHB) kit method. The isolates were found to belong to two genera and four species (Table 3).
Some physiological characteristics of the isolates: The isolates displayed diverse patterns of physiological characteristics (Table 4). All the four isolates were found to grow at the temperature range of 10 to 35°C and none of the isolates were found to grow at 4°C; whereas only one isolate (Bacillus subtilus) was able to grow at 40°C and above.
| Table 1: | Some cellular and morphological characteristics of four bacterial isolates from tef |
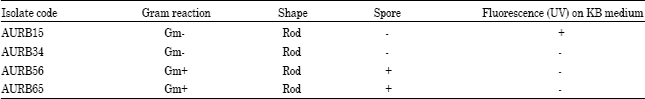 | |
| +: Positive for the test; -: Negative for the test; Gm+: Gram positive; Gm-: Gram negative | |
| Table 2: | Some PGPR and biochemical properties of selected rhizosphere isolates from tef rhizosphere |
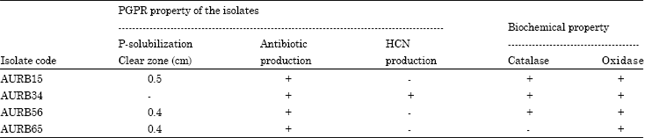 | |
| +: Positive for the test; -: Negative for the test | |
| Table 3: | Identity of the isolates based on API kits |
 | |
| Table 4: | Some physiological characteristics of four selected tef rhizosphere bacterial isolates |
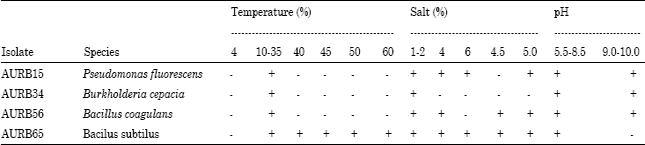 | |
| +: Growth, -: No growth | |
Growth parameters and measurement of tef yield: The pot experiment was undertaken in a 3:1 (soil: sand) culture on two tef varieties (DZ-Cr-37 and DZ-01-974) into which the four selected isolates were isolated. The tef seedlings were found to be green colored and vigorous in growth except those inoculated with AURB34 (Burkholderia cepacia), which visually looked to have relatively stunted growth. However, tef seedlings inoculated with AURB15 (Pseudomonas fluorescens) appeared to be the most vigorous. There was no significant variation in number of tillers and plant height among all the treatments.
Effect of PGPR on root dry weight, shoot dry weight and root shoot ratio of the two tef varieties: The different treatments showed variation of root dry weight (p<0.05). Inoculation of AURB15 (Pseudomonas fluorescens) on DZ-Cr-37 showed significant mean root dry weight difference from that of AURB34 (Burkholderia cepacia) and the control (Table 5). Similarly, inoculation of AURB65 (Bacillus subtilis) on DZ-01-974 showed significant increase in mean dry weight as compared to AURB34 (Burkholderia cepacia).
There was comparable variation among the different treatments on mean shoot dry weight (p>0.05). However, AURB65 (Bacillus subtilis) on DZ-Cr-37 was found to better increase the mean shoot dry weight of both varieties (Table 6).
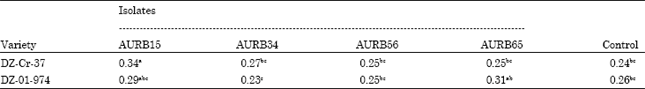
There were also significant differences in root shoot ratio by the treatments (p<0.05). Inoculation of AURB15 (Pseudomonas fluorescens) on DZ-Cr-37 showed significant variation from all the other treatments (Table 7). Likewise, AURB65 (Bacillus subtilis) on DZ-01-974 significantly increased the mean root shoot ratio as compared to AURB34 (Burkholderia cepacia).
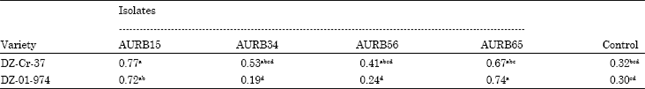
Effect of PGPR isolates on grain yield, grain nitrogen (%) and total grain protein (%) of the two tef varieties: There were significant differences in grain yield by the treatments (p<0.05). It was observed that inoculation of AURB15 (Pseudomonas fluorescens) on DZ-Cr-37 significantly increased mean grain weight as compared to the control (Table 8). Similarly, AURB65 (Bacillus subtilis) on DZ-01-974 showed variation from the uninoculated treatment as well as those inoculated with AURB34 (Burkholderia cepacia) and AURB56 (Bacillus coagulans). It was also observed that inoculation of AURB34 (Burkholderia cepacia) and AURB56 (Bacillus coagulans) on DZ-01-974 resulted in significantly lower mean grain yield as compared to AURB15 (Pseudomonas fluorescens) (p<0.05).
Inoculation of all the four bacterial isolates on tef variety DZ-Cr-37 resulted in comparable grain nitrogen content and total protein with uninoculated groups (p>0.05) (data not presented).
| Table 5: | Effect of inoculation on mean root dry weight (g) of two tef varieties per pot |
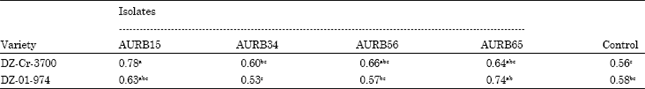 | |
| Means within a column followed by the same letter(s) are not significantly different (p=0.05). Numbers are means of three replications (4 plants per pot) | |
| Table 6: | Effect of inoculation on mean shoot dry weight (g) of two tef varieties per pot |
 | |
| Means within a column followed by the same letter(s) are not significantly different (p = 0.05). Numbers are means of three replications (4 plants per pot) | |
| Table 7: | Effect of inoculation on mean root shoot ratio of two tef varieties |
 | |
| Means within a column followed by the same letter(s) are not significantly different (p = 0.05). Numbers are means of three replications (4 plants per pot) | |
| Table 8: | Effect of PGPR isolates on mean tef grain yield (g) per plant |
 | |
| Means within a column followed by the same letter(s) are not significantly different (p = 0.05). Numbers are means of three replications (4 plants per pot) | |
DISCUSSION
All the four selected isolates were rod-shaped and the Gram reaction indicated that two of them were Gram-positive, the remaining two being Gram-negative bacteria. With regard to fluorescent pigment production on King’s B medium (King et al., 1954), only one isolate (Pseudomonas fluorescens) was found to excrete a pigment as revealed under UV radiation. Failure of fluorescent pigment production by some Fluorescent Pseudomonas sp. has been reported by Bultreys et al. (2003). Identification of our selected isolates using API kit based on carbohydrate utilization showed that the isolates were classified to three genera and four species.
As reflected from characterization of the isolates for their PGPR characteristics, three of our isolates showed phosphate solubilization and only one isolate (Burkholderia cepacia) was failed to grow on Pikovskaya agar medium. However, there was notable difference among the isolates in the degree of phosphate solubilization as indicated by the diameters of clear zone. Antimicrobial activity was confirmed from all four of our isolates during the laboratory screening. Shtark et al. (2003) also reported that rhizosphere bacteria are known to produce antagonistic factors such as antibiotics and other metabolites. On the other hand, only one isolate (Burkholderia cepacia) was positive for cyanide production. The fact that either of the isolates displays at least two of the three PGPR characteristics signals their potential for plant promotion and calls for testing them on plants. Our findings are in agreement with reports of Lucy et al. (2004) and Tilak et al. (2005) who have revealed that phosphate solubilization, antibiotic and cyanide production to be among the mechanisms by which microorganisms promote plant growth. Particularly, the phosphate solubilization by the two Bacillus species and Pseudomonas fluorescence in this investigation is further supported by the finding of Tilak et al. (2005) who reported Pseudomonas and Bacillus to be the main phosphate solubilizing bacteria. Several mechanisms phosphate solubilization by micro organisms such as production of organic acids and chealating oxoacids from sugars (Yadav et al., 2011) and lowering of pH (Gweyi-Onyango et al., 2011; Yadav et al., 2011) others have been reported.
The fact that three of our isolates to be positive for catalase enzyme indicates that they can able to decompose hydrogen peroxide into water and oxygen by the enzyme (Collins and Lyne, 1970). This is important because catalase is among one of the antioxidant enzymes (Joseph and Jini, 2011) that are essential for aerobic life. The fact that three of our isolates isolates to be oxidase positive implies that they may have cytochrome C in their electron transport system as reported by Hunt et al. (1981).
Temperature is one of the major factors that influence microbial growth. In the present study all of our isolates were found to grow in the temperature range of 10-35°C. However, Guillou and Guespin-Michel (1996) reported cessation of growth in Pseudomonas fluorescens MF0 at 35°C.
The growth of most Gram-positive isolates at higher temperatures (50-60°C) shows that they are thermotolerant as reported by Sakali and Yamanami (2006) as observed by one of our isolates (Bacilus subtilus) in the current study.
The ability to adapt to fluctuation in external osmotic pressure and development of mechanisms are fundamental to the survival of microorganisms. The tolerance of three of our isolates to a wide range of % NaCl (w/v) may probably be due to the fact that the isolates might have evolved strategies that could enable them to survive and proliferate in environments of varied salinity. Such ability of growth in different NaCl concentration is attributed to their osmoadaptation as reported by Rangarajan et al. (2002).
Soil pH is among the obvious influencing factors of microbial activity and populations. The growth of our tef rhzosphere isolates over a wide range of pH including acidic pH values suggests that there is a potential to inoculate them under different soil pH conditions if other conditions could not significantly affect their performance. The growth the tef rhizosphere isolates at acidic pH values could probably be because of their adaptation to pH of soil near roots that is usually assumed to be acidic due to CO2 and organic acid release as indicated by Curl and Truelove (1986).
Plants play an important role in selecting and enriching the types of bacteria by the constituents of their root exudates. In the present study, inoculation of AURB15 (Pseudomonas fluorescens) significantly increased root mean dry weight of tef variety DZ-Cr-37 compared to the uninoculated groups (p<0.05). It was also revealed that inoculation of AURB65 (Bacillus subtilis) significantly increased mean root dry weight of tef variety DZ-01-974 compared to mean dry weight obtained by inoculating AURB34 (Burkholderia cepacia). The minimum and maximum mean root dry weight increase in tef variety DZ-Cr-37 by inoculation was found to range from 7 to 39.3% compared with the uninoculated mean dry weght. The maximum increase in mean root dry weight (39.3%) of this tef variety was obtained by inoculating AURB15 (Pseudomonas fluorescens) and minimum increase (7%) was found by AURB34 (Burkholderia cepacia) suggesting that root growth of this tef variety better responded to Pseudomonas fluorescens. Likewise, AURB65 (Bacillus subtilis) increased the mean root dry weight of DZ-01-974 by 27.6% as compared to the uninoculated mean root dry weight. Increase in plant root and shoot dry weight is among the beneficial effects PGPR (Kokalis-Burelle et al., 2002; Lucy et al., 2004). For instance Reddy and Rahe (1989) have reported Bacillus sp. inoculation on different plants to be implicated to increase root dry weight by 13-100% and shoot dry weight by about 12-49%. The increase in mean root dry weight of the tef varieties has also been reported by Zewdie et al. (2000) by inoculation of two Azosprillum bacterial isolates although the % dry weight increase and the tef varieties varied from the present study. The higher mean dry weight by Pseudomonas fluorescens and Bacillus subtilis may be attributed to their antibiosis and phosphate solubilization property as revealed during the laboratory screening of this study.
Comparing the root shoot ratios of inoculated and control groups can also evidence inoculation effect on root development. From the present study of our result, it was revealed that inoculation of AURB15 (Pseudomonas fluoresecns) on DZ-Cr-37 significantly increased mean root shoot ratio (p<0.05). Similarly, AURB65 (Bacillus subtilis) significantly increased mean root shoot ratio of DZ-01-974 as compared to AURB34 (Burkholderia cepacia). The maximum percentage increase of root shoot ratio on dry weight base was 41.7% for tef variety DZ-Cr-37 and 19.23% for DZ-01-974 over the respective control groups. A high root shoot ratio is a clear indication of better root development. In another study, Zewdie et al. (2000) investigated maximum increase in tef root shoot ratio to be 19.6% by inoculation of Azosprillum species.
In this experiment, the treatments showed the grain yield was significantly different (p< 0.05). Accordingly, the tef grain yield of tef variety DZ-Cr-37 obtained by inoculating AURB15 (Pseudomonas fluoresecns) was found to be the highest indicator that the isolate is the best candidate growth promoter of this particular tef variety. It was also investigated that the grain yield DZ-01-974 obtained by inoculating Pseudomonas fluoresecns was significantly higher than the grain yield obtained by inoculation of AURB56 (Bacillus coagulans) and AURB34 (Burkholderia cepacia) as well as that of the control. Likewise AURB65 (Bacillus subtilis) was observed to result in the highest grain yield of DZ-01-974 over the control groups and AURB56 (Bacillus coagulans) and AURB34 (Burkholderia cepacia). The increase in grain yield of tef varieties DZ-Cr-37 and DZ-01-974 by 45 and 42% by inoculating P. fluorescens strongly suggests that the isolate may result in a promising yield of both varieties and calls for evaluation under field condition. It was also revealed that AURB65 (Bacillus subtilis) increased the grain yield of DZ-01-974 by 44% over the control. From the preliminary laboratory result, isolates of AURB15 (Pseudomonas fluorescens) and AURB65 (Bacillus subtilis) were found to be phosphate solubilizers as revealed from clear zone and showed antimicrobial activity against the test organism. Hence, it is possible to suggest that the ability of these bacteria to solubilize phosphate and having other PGPR properties could improve the growth of the respective tef plant. Similar yield increments in varying degree have been reported by inoculation with different PGPR species. Hoflich et al. (1995) indicated that the fluorescent Pseudomonads was found to stimulate development of maize, wheat, oil radish and mustard. Zewdie et al. (2000) reported a yield increase ranging from 4.41 to 10.7% upon inoculation of two tef varieties with Azosprillum sp. Dobbelaere et al. (2001) tested effect of Azosprillum brasilense on sorghum, wheat and barley under field condition and reported a consistent increase of yield up to 26%. Gupta et al. (2002) observed seed bacterization with fluorescent pseudomonads to be a potential method for increasing seedling emergence and suppression of plant pathogenic fungi and enhancing yield in peanut. However, previous investigation by Jagadeesh et al. (2006) revealed that inoculation of two Bacillus sp. designated as Bacillus DHBL and Bacillus DHBS were found to inhibit length of tomato radicle by 37 and 48%, respectively.
CONCLUSION
PGPR application on agriculture is currently attracting many attentions of the scientific community. Tef rhizosphere bacteria were evaluated from variations in their morphology, cellular, physiological, biochemical and PGPR properties. Four isolates, AURB15 (Pseudomonas fluorscens), AURB34 (Bacillus cepacia), AURB56 (Bacillus coagulans) and AURB65 (Bacillus subtilis) were selected based on their PGPR property and evaluated on growth performance and yield of two tef varieties (DZ-Cr-37 and DZ-01-974) at greenhouse condition. Isolate AURB15 (Pseudomonas fluorscens) also resulted in the highest mean dry weight, mean root shoot ratio and grain yield of tef variety DZ-Cr-37. Where as isolate AURB65 (Bacillus subtilis) was found to better increase mean dry weight, mean root shoot ratio and grain yield of tef variety DZ-01-974 suggesting that the isolates are better candidates for the respective tef varieties.
ACKNOWLEDGMENTS
The authors would like to extend their gratitude to the Department of Biology, Addis Ababa University for funding this research.
REFERENCES
- Belay, G., H. Tefera, A. Getachew, K. Assefa and G. Metaferia, 2008. Highly client-oriented breeding with farmer participation in the Ethiopian cereal tef [Eragrostis tef (Zuuc.) Trotter]. Afr. J. Agric. Res., 3: 22-28.
Direct Link - Deepa, C.K., S.G. Dastager and A. Pandy, 2010. Isolation and characterization of plant growth promoting bacteria from non-rhizospheric soil and their effect on cowpea (Vigna unguiculata (L.) Walp.) seedling growth. World J. Microbiol. Technol., 26: 1233-1240.
CrossRefDirect Link - Dobbelaere, S., J. Vanderleyden and Y. Okon, 2003. Plant growthpromoting effects of diazotrophs in the rhizosphere. Crit. Rev. Plant Sci., 22: 107-149.
CrossRef - Glick, B.R., Z. Cheng, J. Czarny and J. Duan, 2007. Promotion of plant growth by ACC deaminase-producing soil bacteria. Eur. J. Plant Pathol., 119: 329-339.
CrossRef - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons Inc., Hoboken, New Jersey, ISBN: 978-0-471-87092-0, Pages: 704.
Direct Link - Gupta, C.P., R.C. Dubey and D.K. Maheshwari, 2002. Plant growth enhancement and suppression of Macrophomina phaseolina causing charcoal rot of peanut by fluorescent Pseudomonas. Biol. Fert. Soil, 35: 399-405.
CrossRefDirect Link - Hoflich, G., W. Wiehe and C. Hecht-Buchholz, 1995. Rhizosphere colonization of different crops with growth promoting Pseudomonas and Rhizobium bacteria. Microbiol. Res., 150: 139-147.
Direct Link - Hunt, L., T.L. Overman and R. Otero, 1981. Role of pH in oxidase variability of Aeromonas hydrophila. J. Clin. Microbiol., 13: 1054-1059.
Direct Link - Hynes, R.K., G.C. Leung, D.L. Hirkala and L.M. Nelson, 2008. Isolation, selection and characterization of beneficial rhizobacteria from pea, lentil and chickpea grown in Western Canada. Can. J. Microbiol., 54: 248-258.
CrossRefPubMedDirect Link - Jalili, F., K. Khavazi, E. Pazira, A. Nejati, H.A. Rahmani, H.R. Sadaghiani and M. Miransari, 2009. Isolation and characterization of ACC deaminase producing fluorescent pseudomonads, to alleviate salinity stress on canola (Brassica napus L.) growth. J. Plant Physiol., 166: 667-674.
Direct Link - King, E.O., M.K. Ward and D.E. Raney, 1954. Two simple media for the demonstration of pyocyanin and fluorescin. Transl. Res., 44: 301-307.
PubMedDirect Link - Kokalis-Burelle N., E.N. Vavrina, E.N. Rosskopf and R.A. Shelby, 2002. Field evaluation of plant growth-promoting rhizobacteria amended transplant mixes and soil solarization for tomato and pepper production in Florida. Plant Soil, 238: 257-266.
Direct Link - Kucey, R.M.N., 1988. Plant growth-altering effects of Azospirillum brasilense and Bacillus C-11-25 on two wheat cultivars. J. Applied Bacteriol., 64: 187-196.
CrossRefDirect Link - Lucy, M., E. Reed and B.R. Glick, 2004. Applications of free-living plant growth-promoting rhizobacteria. Antonie Van Leeuwenhoek, 86: 1-25.
PubMed - Rangarajan, S., L.M. Saleena and S. Nair, 2002. Diversity of Pseudomonas spp. isolated from rice rhizosphere populations grown along salinity gradient. Micob. Ecol., 43: 280-290.
CrossRef - Reddy, M.S. and J.E. Rahe, 1989. Growth effects associated with seed bacterization not correlated with populations of Bacillus subtilis inoculant in onion seedling rhizospheres. Soil Biol. Biochem., 21: 373-378.
CrossRef - Tefera, H., K. Assefa, F. Hundera, T. Kefyalew and T. Teferra, 2003. Heritability and genetic advance in recombinant inbred lines of tef (Eragrostis tef). Euphytica, 131: 91-96.
CrossRef - Sakali, K. and T. Yamanami, 2006. Thermotolerant Bacillus licheniformis TY7 produces optically active L-lactic acid from kitchen refuse under open condition. J. Biosci. Bioeng., 102: 132-134.
PubMed - Shtark, O.Y. and A.I. Shaposhnikov and L.V. Kravchenko, 2003. The production of antifungal metabolites by Pseudomonas chlororaphis grown on different nutrient soureces. Microbiology, 72: 645-650.
PubMed - Yadav, J., J.P. Verma and K.N. Tiwari, 2011. Plant growth promoting activities of fungi and their effect on chickpea plant growth. Asian J. Biol. Sci., 4: 291-299.
CrossRefDirect Link - Oyeyiola, G.P., 2009. Rhizosphere mycoflora of okro (Hibiscus esculentus). Res. J. Soil Biol., 1: 31-36.
CrossRefDirect Link - Nakhro, N. and M.S. Dkhar, 2010. Impact of organic and inorganic fertilizers on microbial populations and biomass carbon in paddy field soil. J. Agron., 9: 102-110.
CrossRefDirect Link - Joseph, B. and D. Jini, 2011. Development of salt stress-tolerant plants by gene manipulation of antioxidant enzymes. Asian J. Agric. Res., 5: 17-27.
CrossRefDirect Link - Gweyi-Onyango, J.P., T. Tesfamariam and G. Neumann, 2011. Contrasting responses to phosphorus status by Arachis pintoi (Krapov and W.C. Gregory): A lesson for selecting vegetables for cultivation in kenyan ecozones. Asian J. Agric. Res., 5: 45-55.
CrossRef - Guillou, C. and A.F. Guespin-Michel, 1996. Evidence for two domains of growth temperature for the psychrotrophic bacterium Pseudomonas fluorescens MF0. Applied Environ. Microbiol., 62: 3319-3324.
Direct Link - Donate-Correa, J., M. Leon-Barrios and R. Perez-Galdona, 2005. Screening for plant growth-promoting rhizobacteria in Chamaecytisus proliferus (tagasaste), a forage tree-shrub legume endemic to the Canary Islands. Plant Soil, 266: 261-272.
CrossRefDirect Link - Dobbelaere, S., A. Croonenboghs, A. Thys, D. Ptacek and J. Vanderleyden et al., 2001. Responses of agronomically important crops to inoculation with Azospirillum. Aust. J. Plant Physiol., 28: 871-879.
CrossRefDirect Link