
Mohammad Taghi Akhi
Department of Medical Microbiology,Tabriz University of Medical Sciences, Tabriz, Iran
Mohammad Reza Nahaei
Department of Medical Microbiology,Tabriz University of Medical Sciences, Tabriz, Iran
Mojtaba Nikbakht
Department of Medical Microbiology,Tabriz University of Medical Sciences, Tabriz, Iran
Mohammad Asgharzadeh
Department of Biotechnology, Faculty of Medicine, Tabriz University of Medical Sciences, Tabriz, Iran
Research Journal of Microbiology
Year: 2008 | Volume: 3 | Issue: 6 | Page No.: 436-446







ABSTRACT
The aims of present investigation were to study the nasal carriage rate of MRSA in hospital staff and in-patients, determination of antibiotic resistant patterns of nasal and clinical MRSA isolates and typing of MRSA isolates by RAPD- PCR. Two hundred and six S. aureus isolates were recovered from clinical specimens and noses of 460 staff and in-patients admitted in Imam Khomeini and Pediatrics hospitals by standard methods during 6 months (2004-2005). Disk agar diffusion (using 13 antibiotics disks) and oxacillin agar screening methods for detection of MRSA isolates were performed according to CLSI. PCR was also used to amplify a 310 bp sequence from S. aureus genome (mecA gene) for detection of MRSA isolates. RAPD-PCR was carried out for fingerprinting of MRSA isolates genome. MRSA isolates were resistant up to 11 antibiotics. All of the MRSA isolates were resistant to penicillin, but sensitive to vancomycin. Of 206 S. aureus isolates, 77 MRSA isolates were detected using disk agar diffusion and oxacillin agar screening methods. In contrast, 80 isolates were detected as MRSA by amplification of mecA sequence. In RAPD-PCR experiments, 43 different RAPD patterns were obtained from our MRSA isolates. Nasal carrier rate of S. aureus was 34.7% and MRSA isolates were high (38.8%) in our hospitals. This study revealed high rate of MRSA, that infected patients and MRSA nasal carriers (staff and in-patients) were the main source of transmission and infection, therefore effective control measures are necessary to avoid nosocomial infection outbreaks.
PDF Abstract XML References Citation
How to cite this article
Mohammad Taghi Akhi, Mohammad Reza Nahaei, Mojtaba Nikbakht and Mohammad Asgharzadeh, 2008. Molecular Fingerprinting of Methicillin-Resistant Staphylococcus aureus Isolates in Hospital Staff and Patients. Research Journal of Microbiology, 3: 436-446.
URL: https://scialert.net/abstract/?doi=jm.2008.436.446
URL: https://scialert.net/abstract/?doi=jm.2008.436.446
INTRODUCTION
Staphylococcus aureus is consistently one of the top four causes of nosocomial infections. S. aureus causes a variety of suppurative infection and toxinoses in humans (Gardam, 2000). At present approximately 40% of S. aureus are resistance to methicillin and incidence of MRSA increases year by year. MRSA infections cause substantial morbidity, mortality and serious MRSA infections must be treated with vancomycin. Thus in hospitals, the high rate of MRSA increase the use of this valuable antimicrobial agent, which in turn may increase the risk for selection of vancomycin-resistant isolates (Herwaldet, 1999). Nasal carriers in hospital wards are the most common source of MRSA infection and increase in frequency of MRSA as the causal agent of nosocomial infection and in the possibility of emergence of resistance to vancomycin demands a quick and trustworthy characterization of isolates and identification of clonal spread within hospitals (Herwaldet, 1999; Tenover et al., 2001).
The greatest challenge for hospital and clinical laboratory is to carry out an analysis that can reliably group epidemiologically related strains and discriminate them from unrelated strains. Usually, conventional methods for identification of S. aureus are colony morphology, gram staining, culture, biochemical tests, DNAase and coagulase test. Because of constant genetic variations, molecular techniques have particular importance for identification and typing of MRSA and in epidemiology researches (Stefani and Agodi, 2000; Tindade et al., 2003; Oliveria and Ramos, 2002). Compared to other molecular methods, RAPD-PCR is better than the others because it is cheap, simple, more sensitive than other molecular methods and fast (Tambic et al., 1997). In the present study we characterized the MRSA strains isolated from carriers and compared them with MRSA strains isolated from different clinical specimens obtained from patients.
MATERIALS AND METHODS
Sample Collection
A prospective study was performed, which during two months period in 2005, 460 in-patients and hospital staff (220 staff and 240 in-patients of clinical wards, including: Infectious, Internal, ENT, Infant, Oncology, ICU, Surgery, Operation room, Dialyse) in Imam Khomeini and Pediatric hospitals (Tabriz) were screened for S. aureus and MRSA by nasal swabs. Nasal swabs were taken from over 48 h bedridden patients and medical and nursing staff. Patients who expired in the period of study, or did not consent were excluded from the study. From Dec. 2004 until June 2005, clinical specimens submitted to the teaching hospital laboratories were also cultured for S. aureus and tested for MRSA.
Microbiological Investigations
All swabs were cultured into nutrient broth and incubated at 37°C. After 24 h, plated onto blood agar and mannitol salt agar media by sterile loops and were incubated at 37°C for 24 h (Jawetz et al., 2004). Pure cultures were obtained and then colonies were tested for Gram staining, catalase production, bound and free coagulase and DNAase activity (Bennerman, 2003). The susceptibilities of the isolates to ciprofloxacin, clindamycin, erythromycin, gentamicin, rifampin, penicillin, co-trimoxazole, methicillin, amikacin, oxacillin, vancomycin, amoxicillin clavulanic acid, lincomycin (Mast) were tested by agar disk diffusion method (Clinical and Laboratory Standards Institute, 2006; formerly NCCLS).
Resistance to methicillin was detected using 1 μg oxacillin and 5 μg methicillin disks (Duguid, 1989) and oxacillin agar screening plate (Jureen et al., 2001) and amplification of mecA gene by PCR (Araj et al., 1999). Two hundred and six isolates were recovered that composed of 160 isolates from nasal swabs and 46 isolates from clinical samples. The strains were stocked in 10% skim milk at -80°C until use.
DNA Extraction and Amplification of mecA Gene
DNA extraction was performed as described by Van Soolingen et al. (1994).
Primers were purchased from TIB MOLBIOL, Germany. PCR was performed with two primers for amplification of 310 bp piece of MRSA genome, the sequence of primers were as:
Forward: mecA1 (5`- gTAgAAATgACTgAACgTCCgATAA-3`), 25-mer
Reverse: mecA2 (5`- CCAATTCCACATTgTTTCggTCTAA-3`), 25mer
The two primers (mecA1, mecA2), dNTP and other reagents were prepared according to the manufacturer`s recommendations. The total volume of PCR mix was 25 μL, including: Sterile redistilled H2O 17.05 μL, 10X PCR buffer 2.5 μL, dNTP mix (10 mM) 0.5 μL, MgCl2 (50 mM) 0.75 μL, forward primer (25 μM) 0.5 μL, reversed primer (25 μM) 0.5 μL, Taq DNA polymerase (5 U μL-1) 0.2 μL, template DNA 3 μL. Negative controls contained all components except template DNA. PCR reactions were performed with an automated thermal cycler (Eppendorf mastercycler gradient, Germany) with the PCR cycling conditions (initial cycle at 94°C for 4 min, 30 cycles of denaturation at 94°C for 40 sec, annealing at 57°C for 40 sec and extension at 72°C for 40 sec, final cycle extension at 72°C for 7 min) (Araj et al., 1999).
RAPD-PCR
Primers were purchased from TIB MOLBIOL (Germany). The sequences of primers were as:
| MN45 (A) | 5`- AAgACgCCgT-3` |
| KAY1 (B) | 5`- AgCAGCCTgC-3` |
| EP015 (C) | 5`- ACAACTgCTC-3` |
| EP017 (D) | 5`- TACACCCgTCAACATTgAgg-3` |
| EP007 (E) | 5`- AgCACgCTgTCAATCATgTA-3` |
The above mentioned primers and other reagents were prepared according to the manufacturer`s recommendations. The DNA amplification was performed at final volume of 25 μL, including: sterile redistilled H2O 16.35 μL, 10X PCR buffer 2.5 μL, dNTP mix (10 mM) 1.5 μL, MgCl2 (50 mM) 1.25 μL, primer (25 μM) 1 μL, Taq DNA polymerase (5 U μL-1) 0.4 μL, template DNA 3 μL and negative control for each of primers contained all components except template DNA. RAPD-PCR reactions were performed for each five primers with the automated thermal cycler (introduced earlier) with the PCR cycling conditions (initial cycle at 94°C for 5 min, 40 cycles of denaturation at 94°C for 30 sec, annealing at 35°C for 40 sec and extension at 72°C for 60 sec, final cycle extension at 72°C for 7 min) (Tambic et al., 1997, 1999).
Amplification products were separated by electrophoresis in 1.2% agarose gel for 60-120 min at 75 v in 1x TBE buffer and staining of the gel was carried out in 0.5 μL mL-1 of ethidium bromide solution for 30 min. The gel was visualized on UV transilluminator and photographed using gel documentation system for the analysis of bands (Van Soolingen et al., 1994).
Statistical Analysis
Information related to in-patients and personal were collected by questionnaires. The data were analyzed by statistic software SPSS-11 and Chi-square tests. Statistical results were interpreted as significant when p-value was recorded as <0.05. For each primer the banding patterns were scored as presence (1) or absence (0) for each MRSA isolates.
RESULTS
Among the 460 subjects (220 staff and 240 in-patients), one hundred and sixty S. aureus isolates were collected from nose of staff and in-patients which 48 isolates (30%) were diagnosed as MRSA. Forty six S. aureus isolates were also collected from clinical specimens (wound, blood, tracheal fluid, pleural fluid, urine, ascit fluid, catheter, umbelical cord) of in-patients from various wards in 6 months periods. Among these 46 isolates, 32 (69.5%) were MRSA. Percentage of MRSA in clinical specimens was higher (69.5%) than that of nasal specimens (30%), showing statistically difference (p<0.001). Finally, from 206 isolates of S. aureus, we detected 80 MRSA isolates (38.8%) with mecA gene by PCR method; in contrast we identified 77 MRSA isolates (37.4%) by oxacillin agar screening and oxacillin disk diffusion methods.
Antibiotic Susceptibility Testing
Antibiotic sensitivity tests were carried out on all isolates of MRSA, by using disk diffusion technique according to CLSI recommendation. The highest numbers of antibiotics that our 77 MRSA
| Table 1: | Antibiotic resistance pattern of MRSA isolates |
 | |
| *Patterns of MRSA; M, Methicillin; OX, Oxacillin; P, Penicillin; GM, Gentamicin; AMC, Amoxicillin-Clavulanate; CC, Clindamicin; R, Rifamicin; TS, Cotrimoxazol; VA, Vancomicin; CIP, Ciprofloxacin; L, Lincomycin; AN, Amikacin; E, Erythromycin | |
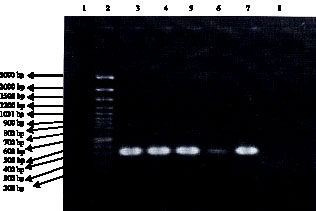 |
| Fig. 1: | 310 bp banding patterns of MRSA isolates in gel electrophoresis (1.2% agarose). Lane 1: size marker (100 bp DNA Ladder plus). Lanes 2-6: 310 bp PCR band of MRSA isolates. Lane 7: S. aureus standard strain (ATCC 29213). Lane 8: Negative control |
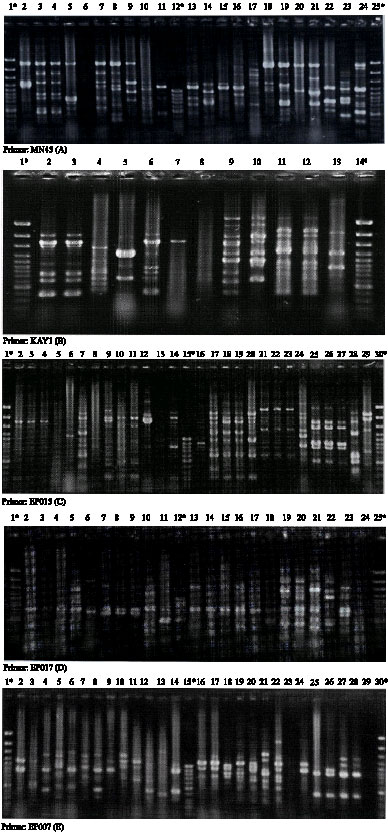 |
| Fig. 2: | RAPD patterns of some MRSA isolates with 5 primers used in this study, *, Lanes for molecular size marker (100 bp DNA Ladder plus and 100 bp DNA Ladder); numbers, MRSA isolates |
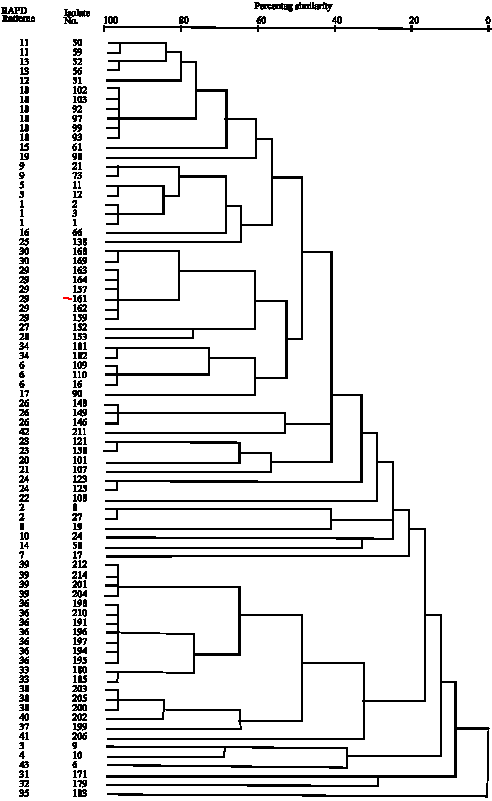 |
| Fig. 3: | Dendrogram for 80 isolates of MRSA based on RAPD-PCR data using 5 studied primers |
isolates were resistant to them were 11 (84.6%) and the least was 1 (7.7%) (Table 1). MRSA isolates showed multi-resistance to antibiotics and the highest resistance was related to penicillin 77 (100%) and co-trimoxazol 60 (78%) and the lowest resistance recorded against vancomycin 0 (0%) and rifampicin 21 (27.3%). Following resistances against test antibiotics were also obtained: ciprofloxacin (75.3%), erythromycin (70%), lincomycin (58.4%), gentamycin (44.2%), clindamycin (37.7%), amikacin (36.3%), amoxicillin-clavulanate (35%). Forty one resistance patterns were recorded for 77 MRSA isolates which were designated as R1- R41 (Table 1).
Figure 1 shows an example of mecA gene amplification and Fig. 2 are example of RAPD pattern with 5 primers (full data for all strains are available upon request). Dendrogram for 80 MRSA isolates based on RAPD-PCR data using 5 studied primers have been shown in Fig. 3.
DISCUSSION
Staphylococcus aureus is one of the top four causes of nosocomial infections, along with Escherichia coli, Enterococcus faecalis and Pseudomonas aeruginosa and is important cause of community acquired disease such as follicolitis, endocarditis, toxic shock syndrome, osteomyelitis and etc. (Gardam, 2000; Choi et al., 2006). Currently S. aureus is second only to coagulase negative staphylococci as a cause of hospital-acquired bacteremia and is the leading potentially lethal cause of these infections (Archer, 1998). The nasal cavity is the most common site to harbor S. aureus and the organism may be carried by up to 40% in healthy individuals. This nasal carriage of S. aureus appears to play a key role in the epidemiology and pathogenesis of staphylococcal infections (Davis et al., 2005). In the early 1950s, acquisition and spread of beta-lactamase-producing plasmids reduced the effectiveness of penicillin for treating S. aureus infections. In 1959, methicillin, a semisynthetic penicillin, was introduced (Gilmore et al., 2002). Therefore, rapid and reliable isolation and diagnosis of S. aureus and MRSA in clinical specimens are important for appropriate patient care and control of this microorganism in hospitals (Kluytmans et al., 2002). Conventional culture and susceptibility test procedures and PCR-based methods for detection of antimicrobial resistance have been applied to bacteria including methicillin-resistant Staphylococcus aureus (Louie et al., 2000). All strains of methicillin-resistant S. aureus produces a unique penicillin-binding (PBP2a) encoded by a chromosomal gene, mecA. The mecA gene is not present in susceptible strains. PCR has been used successfully to amplify and detect mecA gene sequences from clinical isolates and directly from clinical specimens within a few hours. Another potential use of the assay is to assist in epidemiologic studies in the setting of an outbreak. We applied both conventional and molecular methods for detection of MRSA in 206 S. aureus isolates. Eighty MRSA isolates (38.8%) were detected to have mecA gene by PCR method; in contrast we identified 77 MRSA isolates (37.4%) by oxacillin agar screening and oxacillin disk diffusion methods. This study also revealed the prevalence of the mecA gene in all nasals and clinical MRSA isolates which correlated with the phenotypic resistance in MRSA isolates. This finding suggests that the methicillin resistance mechanism in the MRSA isolates was due to production of PBP2a by mecA gene. Examination of mecA gene in 206 nasal and clinical isolates of S. aureus showed that, while there was a gross correlation between the presence of the gene and the level of bacterial resistance to oxacillin and methicillin, 3 mecA positive isolates could not be distinguished from the mecA-negative ones by the susceptibility test because of their inability to produce PBP2a. The occurrence of methicillin-resistant variants described above implied the possibility that, during chemotherapy with β-lactam antibiotics, atypical resistant subpopulation occurred in a cryptic methicillin-resistant S. aureus strain (a mecA-positive but non-PBP2a-producing strain), even when strains could not be identified as resistant by conventional susceptibility tests in clinic. For this reason, all mecA-positive isolate, must be detected precisely (Baddour et al., 2007). In our study 3 cryptic isolates were identified which 2 of them isolated from oncology ward (nasal swabs) and one isolate recovered from dialysis ward (nasal swab). All of the 3 patients were bedridden for long period of time in hospital receiving antibiotics.
The nasal carrier rate in our study was higher (34.7%) than what was earlier reported from some hospitals (Tambic et al., 1999; Choi et al., 2006) and similar results have also been reported in other studies (Lederer et al., 2007; Kuehnert et al., 2006). Results of the present study showed that MRSA encountered in 30% of nasal isolate and 69.5% of clinical isolates with average of 38.8% in 206 S. aureus isolates studied. Therefore, the rate of MRSA was higher in the S. aureus isolates from clinical infections, compared to those from carriers, which was statistically significant (p<0.001). These infected patients, when hospitalized, may constitute an important sources of cross infection to other hospitalized patients and health care workers. Since complete eradication of MRSA is not possible, control of transmission seems to be the appropriate goal.
Resistance of MRSA isolates to co-trimoxazol, ciprofloxacin, erythromycin and lincomycin was high, while rifampicin, amoxicillin-clavulanate, clindamycin and gentamicin remained active. Similar results have been obtained for erythromycin and ciprofloxacin in Malaysian hospitals (Rohani et al., 2000). Although the multi resistance ranged from 4 to 11 antibiotics, the choice of the effective chemotherapy in many instances was limited to vancomycin and rifampicin (Table 1). It has been reported that most S. aureus isolates with reduced susceptibility to vancomycin appear to have developed from preexisting methicillin -resistant S. aureus infections and many of the isolates with reduced susceptibility to glycopeptides have been associated with therapeutic failures with vancomycin (Srinivasan et al., 2002). Therefore, accurate epidemiological typing is of primary importance for the identification of MRSA clones found in a hospital and for enabling sources and routs of transmission to be identified and controlled (Tenover et al., 2001).
Bacterial strain typing has become an important clinical tool to investigate suspected outbreaks and to evaluate nosocomial transmission (Poxton, 2005). With the advent of molecular biology, strain typing focused on DNA-based methods. Among different techniques, RAPD-PCR also called arbitrarily primed PCR that use short primers with an arbitrary sequence to amplify genomic DNA, has been used successfully (Tambic et al., 1999). In this study we analyzed MRSA isolates by five primers, with good discriminatory power in other studies (Fig. 2) (Tambic et al., 1999; Hermans et al., 2000). In total, 43 different patterns were obtained from 80 MRSA isolates that analyzed at a similarity level of 95%. When the level of similarity was lowered, cluster formation decreased. For example 80 MRSA isolates located into 18 clusters at a similarity level of 50%, indicating their molecular diversity. We found that MRSA isolates with the same RAPD and antibiotic resistance patterns related to certain ward. However, in some cases, MRSA isolates with the same RAPD and antibiotic resistance pattern were isolated from different wards which suggest transfer of staff and in-patients among wards inside the hospital. Some researchers have shown that antibiotic resistance pattern may serve as reliable markers for typing of MRSA isolates (Tambic et al., 1999), but in this study in some cases antibiotic resistance pattern was unhelpful (Table 1), because some MRSA isolates with distinctive RAPD pattern fell into different antibiotic resistance pattern and vice versa. However, antibiotic resistance pattern when combined by RAPD pattern is helpful for MRSA typing (Louki et al., 2003). This fact that MRSA isolates in our hospitals with identical RAPD pattern were isolated from staff and in-patients of certain wards, shows that the principal route of MRSA transmission was via patient to patient or patient to staff and vice versa. Therefore, infected and colonized patients as well as carrier hospital staff are the main sources of MRSA in our hospitals. Louki et al. (2003) showed that antibiotic susceptibility profile combined by AP-PCR with different primers could successfully identify the two major MRSA clone that are responsible for outbreaks in the hospital of Patras as well as in other tertiary care hospital in Greece. Tambic et al. (1999) studied on 42 MRSA isolates that were detected from in-patient and staff in Zagreb hospital, their study showed that 39 of 42 MRSA isolates shared the same antibiotic sensitivity pattern, suggesting endemic spread of MRSA; however RAPD-PCR revealed four profiles, the most common involving 15 of 36 tested isolates (Tambic et al., 1999). Tambic et al. (1997) reported analysis of an outbreak of non-phage type-able MRSA by RAPD-PCR, their results revealed that the 15 of 21 isolates were cause of outbreak and located into a single cluster at a similarity level of 76% and other 6 isolates fell into 3 clusters, but 21 isolates fell into 11 clusters at a similarity level of 90%. Lee (2003) from Korea veterinary public health identified that six of the MRSA isolates from animals were identical to the patterns of certain isolates from humans and suggested possible source of human infections caused by consuming contaminated food products made from these animals. In the above mentioned study 19 MRSA isolates divided into 10 RAPD patterns. Since the hospitals studied in this research are reference hospitals in the north-west of Iran and they include various crowded wards and many patients are admitted from other healthcare settings, the various patterns of MRSA isolates in our hospitals were probably due to introduction of strains from other institution by transferring of patients. There was no obvious distinctive MRSA clone-related infection in our study; nevertheless, routine infection control surveillance is necessary for prevention of epidemic emergence.
CONCLUSION
Results of the present study demonstrated that the nasal carriage rate of Staphylococcus aureus was 34.7% in our hospitals and 30% of nasal S. aureus isolates and 69.5% of clinical isolates in the same period were resistant to oxacillin (MRSA). The mecA gene was detected in all MRSA isolates in both groups. Most of the isolates were multi resistant (up to 11 antibiotics) and 100% of MRSA isolates were resistant to penicillin and sensitive to vancomycin. Infected and colonized patients as well as hospital staff are the main source of MRSA in our hospitals and lack of standard control measures, deteriorate the situation. The various patterns and high rate of MRSA isolates in our hospital wards were probably due to the introduction of strains from other institution by transferring of patients. Thus precise diagnostic methods for detection of MRSA and consequent studies for detection of transmission sources are necessary. Strict monitoring and epidemiological investigation are essential to control MRSA and to avoid the emergence of VISA or VRSA in our hospitals.
ACKNOWLEDGMENTS
This study was funded with a research grant (No. 83-84) from Drug Applied Research Center of Tabriz University of Medical Sciences, Tabriz Iran. We would also like to thank hospital staff of Imam Khomeini and Pediatrics for their participation in this study.
REFERENCES
- Archer, G.L., 1998. Staphylococcus aureus: A well-armed pathogen. Clin. Infect. Dis., 26: 1179-1181.
PubMed - Choi, C.S., C.S. Yin, A.A. Bakar, Z. Sakewi, N.N. Naing, F. Jamal and N. Othman, 2006. Nasal carriage of Staphylococcus aureus among healthy adults. J. Microbiol. Immunol. Infect., 39: 458-464.
Direct Link - Davis, K.A., J.J. Stewart, H.K. Crouch, C.E. Flores and D.R. Hospenthal, 2004. Methicillin-Resistant Staphylococcus aureus (MRSA) nares colonization at hospital admission and its effect on subsequent MRSA infection. Clin. Infect. Dis., 39: 776-782.
CrossRefPubMedDirect Link - Gardam, M.A., 2000. Is methicillin-resistant Staphylococcus aureus an emerging community pathogen? A review of the literature. Can. J. Infect. Dis., 11: 202-211.
PubMedDirect Link - Jureen, R., K.L. Bottolfsen, H. Grewal and A. Digranes, 2001. Comparative evaluation of a commercial test for rapid identification of methicillin-resistant Staphylococcus aureus. Ovid, 109: 787-790.
Direct Link - Kuehnert, M.J., D. Kruszun-Moran, H.A. Hill, G. McQuillan and S.K. McAllister et al., 2006. Prevalence of Staphylococcus aureus nasal colonization in the United States, 2001-2002. J. Infect. Dis., 193: 172-179.
Direct Link - Lederer, S.R., G. Riedelsdorf and H. Schiffl, 2007. Nasal carriage of methicillin resistant Staphylococcus aureus: The prevalence, patients at risk and the effect of elimination on outcomes among out clinic haemodialysis patients. Eur. J. Med. Res., 12: 284-288.
Direct Link - Lee, J.H., 2003. Methicillin (oxacillin)-resistant Staphylococcus aureus strains isolated from major food animals and their potential transmission to humans. Applied Environ. Microbiol., 69: 6489-6494.
CrossRefPubMedDirect Link - Louie, M., L. Louie and A.E. Simor, 2000. The role of DNA amplification technology in the diagnosis of infectious disease. Can. Med. Assoc. J., 163: 301-309.
Direct Link - Louki, C.B., G. Dimitracopoulos and I. Spiliopoulou, 2003. Polymerase chain reaction fingerprints of methicillin-resistant Staphylococcus aureus clinical isolates in Greece are related to certain antibiotypes. J. Microbiol. Methods, 53: 417-422.
Direct Link - Oliveria, A.M. and M.C. Ramos, 2002. PCR-based ribotyping of Staphylococcus aureus. Braz. J. Med. Biol. Res., 35: 175-180.
Direct Link - Poxton, I.R., 2005. Molecular techniques in the diagnosis and management of infectious diseases: Do they have a role in bacteriology? Med. Princ. Pract., 14: 20-26.
Direct Link - Rohani, M.Y., A. Raudzah, M.G. Lau, A.A.R. Zaidatul and M.N. Salbiah et al., 2000. Susceptibility pattern of Staphylococcus aureus isolated in Malaysian hospitals. Int. J. Antimicrob. Agents, 13: 209-213.
Direct Link - Srinivasan, A., J.D. Dick and T.M. Perl, 2002. Vancomycin resistance in Staphylococci. Clin. Microbiol. Rev., 15: 430-438.
Direct Link - Stefani, S. and A. Agodi, 2000. Molecular epidemiology of antibiotic resistance. Int. J. Antimicrob. Agents, 13: 143-153.
Direct Link - Tenover, F.C., J.W. Bidelle and M.V. Lancaster, 2001. Increasing resistance to vancomycin and other glycopeptides in Staphylococcus aureus. Emerg. Infect. Dis., 7: 327-332.
PubMedDirect Link - Tindade, P.A., J.A. McCulloch, G.A. Oliveria and E.M. Mamizuka, 2003. Molecular techniques for MRSA typing: Current issues and perspectives. Braz. J. Infect. Dis., 7: 32-43.
Direct Link - Van Soolingen, D., P.E.W. De-Hass, P.W.M. Hermans and J.D.A. Van-Embden, 1994. DNA fingerprinting of Mycobacterium tuberculosis. Methods Enzymol., 235: 196-205.
Direct Link