
Ehab R. EL-Haroun
Fish Nutrition Research Laboratory, Department of Animal Production, Faculty of Agriculture, Cairo University, Cairo, 12411, Egypt
Journal of Fisheries and Aquatic Science
Year: 2007 | Volume: 2 | Issue: 5 | Page No.: 319-327







ABSTRACT
The present study was conducted to examine probiotic treatment in the diet of African catfish Clarias gariepinus. A total of 240 of African catfish (weight ranged from 81.13 to 91.35 g) were divided in the five experimental groups. The experiment was conducted for 120 days. Experimental diets were identical in all except for the variation in probiotic levels. A probiotic (Biogen7) was used at 0% (control), 0.5% (diet 2), 1.0% (diet 3), 1.5% (diet 4) and 2.0% (diet 5) inclusion rates in the experimental diets. The growth performance and nutrient utilization of African catfish including Weight Gain (WG), Specific Growth Rate (SGR), Protein Efficiency Ratio (PER), Protein Productive Value (PPV) and Energy Retention (ER) were significantly (p#0.05) higher of fish fed diets containing 0.5% Biogen7 than for fish fed control diets. In the same trend fish fed 0.5% Biogen7 recorded the best-Feed Conversion Ratio (FCR) (p#0.05) compare to the control diet. However, among the probiotic fed groups no significant differences were recorded (p$0.05). Concerning the influence of different dietary probiotic levels on chemical proximate analysis of whole carcass, protein content was significantly higher (p#0.05) at 0.5% inclusion, while, fish fed diet containing 1.0% Biogen7 recorded the highest ash content without significant (p$0.05) difference with the other treatments. Differences were observed in carcass lipid and gross energy content with significantly lowest (p#0.05) values recorded in fish fed 2.0% Biogen7. The production performance and subsequent cost-benefit analyses clearly indicated that the diets containing 0.5% probiotic Biogen7 recorded the highest net return compared to the control diet. Based on the result of this study, it is concluded that Biogen7 positively influenced growth performance and feed utilization of African catfish as well as the economic efficiency.
PDF Abstract XML References Citation
How to cite this article
Ehab R. EL-Haroun, 2007. Improved Growth Rate and Feed Utilization in Farmed African Catfish Clarias gariepinus (Burchell 1822) Through a Growth Promoter Biogen7 Supplementation. Journal of Fisheries and Aquatic Science, 2: 319-327.
DOI: 10.3923/jfas.2007.319.327
URL: https://scialert.net/abstract/?doi=jfas.2007.319.327
DOI: 10.3923/jfas.2007.319.327
URL: https://scialert.net/abstract/?doi=jfas.2007.319.327
INTRODUCTION
The African catfish, Clarias gariepinus (Burchell, 1822), is one of the most highly valued species in Africa (Egypt, Ethiopia, Ghana, Mali and Nigeria) and Asia (China, Indonesia, Malaysia, Philippines and Thailand). Recently, Clarias gariepinus has been introduced in some European and Latin American countries and its culture has increased in scale (FAO, 1997; Ali, 2001). African catfish is considered one of the best suitable alternatives to tilapia for subsistence fish farming in Africa. Yields of catfish from ponds could be higher than those of tilapia while using low grade feed composed of local agricultural by-products (Hogendoorn, 1983; Goda et al., 2007).
Catfish is cultured for its high growth rate, disease resistance and it’s tolerant of a wide range of temperature, as well as oxygen and high salinity levels (Huisman and Richter, 1987; Haylor, 1993; Bovendeur et al., 1987). The superior performance of Clarias gariepinus compared to other Clarias species in terms of growth rate has probably contributed to the fact that C. gariepinus has been widely introduced to areas outside its natural range (Verreth et al., 1993).
Antibiotics had been frequently used to enhance growth and/or resistance to diseases in aquaculture systems. However, the excess use of antibiotics to prevent bacterial diseases in aquaculture, have led to an increase in antibiotic-resistant bacteria (Teuber, 2001). In recent years, there have been increasing the beneficial effects of the probiotic in the aquaculture of many economically important marine and freshwater fish species as an alternative to the use of antibiotics in feeds. Evidence of the beneficial effects of probiotics increased the demand of probiotic in aquaculture, which are defined by Gatesoupe (1999) as “microbial cells that are administered in such a way as to enter the gastrointestinal tract and to be kept alive, with the aim of improving health”.
Recently, many kinds of probiotics have been commercially produced and used as feed additives in farmed fish such as turbot, Nile tilapia, salmon, trout, eel, sole and gilthead seabream (Robertson et al., 2000; Lara-Flores et al., 2003; Nikoskelainen et al., 2003; Hjelm et al., 2004; Lategan et al., 2004; Chabrillon et al., 2005, 2006; Picchietti et al., 2007).
Probiotic Biogen® consists of Bacillus species bacteria. The advantage of these bacteria is that they are able to survive the pelletization process. After transit through the stomach, they germinate in the intestine and use a large number of sugars (carbohydrates) for their growth and produce a range of relevant digestive enzymes (amylase, protease and lipase).
The present study was conducted to determine the effect of feeding commercial probiotic Biogen® on growth performance and nutrient utilization in African catfish.
MATERIALS AND METHODS
Experimental Fish and Culture Technique
This study was performed at the Middle East Land Reclamation Company (MELARC) Farm, Tamoo, Giza, Egypt. African Catfish Clarias gariepinus was obtained from a commercial supplier (Elobor market) Cairo, Egypt. The average initial body weight for African catfish ranged from 81.13 to 91.35 g. In March-15-2003, 240 African catfish were equally divided into 15 groups and carefully stocked into 15 ponds 8 m3 each and 1.25 m depth 16 fish in each pond. The fish were adapted to experimental conditions for 2 weeks prior to the start of the experiment and were fed a commercial diet twice a day. The actual experimental period extended for 17 weeks. The ponds were supplied with freshwater. The turnover rate of water was 0.4 m3 day-1 pond-1 and fish were held under natural light. Photoperiod was maintained at a 12:12 h light: dark schedule. During the trial, water temperature and Dissolved Oxygen (DO) were measured using an YSI model 56 oxygen meter (Yellow Springs Instrument, Yellow Springs, OH, USA) and pH by using a pH meter (Orion pH meter, USA). Ammonia and alkalinity were measured at weekly intervals according to APHA, AWWA, WPCF (1985).
All dietary treatments were tested in triplicate groups where each pond was considered as an experimental unit. During the experiment, all fish were fed their respective diets at a level of 3% of body weight for 6 days a week. The daily ration was divided into three equal amounts and offered three times a day (0900, 1200 and 1500). A random sample of fish from each treatment was weighed biweekly and the amount of daily diet was adjusted accordingly.
Experimental Diets
Five isonitrogenous and isocaloric diets were formulated for the experiment (Table 1). The control diet had no Biogen® added and four other test diets included Biogen® added at 0.5, 1.0, 1.5 and 2.0% levels, respectively. The fish meal, soybean meal, wheat bran and yellow corn were purchased from the Islamic Company (APICO), Dokki-El-Giza Egypt. Biogen® was obtained from the El-Zahra Veterinary Trading Company, an exclusive agent of the manufacturer, China Way Corporation Taiwan, Roc). The Biogen® is reported to contain: Allicin (not less than 0.247 M mole g-1), Bacillus subtilis not less than 6x107 g-1) and High Unit Hydrolytic Enzyme (not less than 3690 Unit g-1).
| Table 1: | Formulation and proximate analysis of experimental diets (% dry matter) |
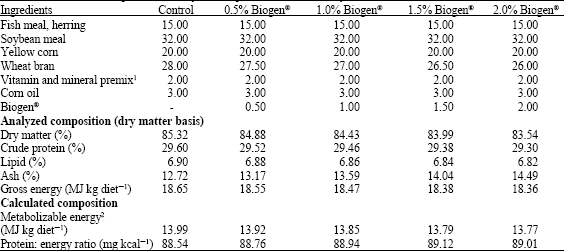 | |
| 1Vitamin and mineral mixture each 1 kg of mixture contains: retinyl acetate (vit. A), 3000 IU; cholecalciferol (vit. D), 2400 IU; all-rac-α-tocopheryl acetate (vit. E), 60 IU; menadione sodium bisulfite (vit. K), 1.2 mg; ascorbic acid monophosphate (49% ascorbic acid, vit. C), 120 mg; cyanocobalamine (vit. B12), 0.024 mg; d-biotin, 0.168 mg; choline chloride, 1200 mg; folic acid, 1.2 mg; niacin, 12 mg; d-calcium pantothenate, 26 mg; pyridoxine. HCl, 6 mg; riboflavin, 7.2 mg; thiamin. HCl, 1.2 mg; sodium chloride (NaCl, 39% Na, 61% Cl), 3077 mg; ferrous sulfate (FeSO4.7H2O, 20% Fe), 65 mg; manganese sulfate (MnSO4, 36% Mn), 89 mg; zinc sulfate (ZnSO4.7H2O, 40% Zn), 150 mg; copper sulfate (CuSO4.5H2O, 25% Cu), 28 mg; potassium iodide (KI, 24% K, 76% I), 11 mg; Celite AW521 (acid-washed diatomaceous earth silica), 1000 mg. 2ME was calculated as 75% of GE according to Hepher et al. (1983) | |
The diets were processed by blending the dry ingredients into a homogenous mixture and then passing the mixed feed through a laboratory pellet mill (California Pellet Mill Co., San Francisco, CA). The chemical composition of experimental ingredients and diets are presented in Table 1.
Sampling and Chemical Analysis
At the beginning of the experiment, a sample of 10 fish was taken for determining chemical body composition (initial carcass sample). At the end of the experiment, 5 fish from each tank were sampled. Fish were killed with an excess concentration of anesthetic (t-amyl alcohol) and then weighed. The fish were then pooled per pond (final carcass sample) for chemical analyses. The pooled fish samples were cooked in an autoclave, ground into homogeneous slurry in a food processor and oven dried, finely ground in a coffee grinder and stored at -20°C until analyzed. Diet, ingredients and carcass samples were analyzed for Dry Matter (DM) and ash according to AOAC (1995), crude protein (%Nx6.25) by Kjeldahl method using a Kjeltech autoanalyzer (Model 1030, Tecator, Höganäs, Sweden) and total lipid according to the method of Bligh and Dyer (1959). Gross energy content of carcass samples was measured using an automated bomb calorimeter (Model 1272, Parr Instruments Inc., Moline, IL USA).
Calculations Parameters
Mean fish weight of each treatment pond was determined by dividing total fish pond weight by number of fish in each pond. Weight Gain (WG), Specific Growth Rate (SGR), Feed Conversion Ratio (FCR), Protein Efficiency Ratio (PER), Protein Productive Value (PPV) and Energy Retention (ER) was calculated using the following equations:
WG = Final body weight (g) - initial body weight (g);
SGR = [(ln FBW - ln IBW) x100] period day-1,
Where,
| FBW | : | Final Body Weight (g); |
| IBW | : | Initial Body Weight (g); |
| ln | = | Natural logarithmic. |
FCR was calculated from feed intake (g)/wet weight gain (g);
PER was calculated as:
Wet weight gain (g)/protein intake (g);
PPV as (Retained protein/protein intake) x100 and
ER = (Retained energy/energy intake)x100.
Net production (kg) pond-1 = TBMH - TBMS,
where,
TBMH is the total body mass at the harvest (kg) pond-1,
TBMS is the total body mass at the start (kg) pond-1;
TBMH (kg) = (FBW/1000) *16;
TBMS (kg) = (IBW/1000) *16;
Total income (LE) pond-1 = TBMH * price (6.0 LE kg-1);
Net return (LE) pond-1 = Total income (LE) pond-1 - total cost (LE) pond-1;
Profit (%) = (net return (LE) pond-1/total cost (LE) pond-1) * 100.
Statistical Analysis
Growth performance, feed utilization efficiency and body chemical composition parameters were statistically compared using a one-way ANOVA (p≤0.05) and differences among means were identified using the Duncan Multiple Range Test. GLM procedure of SAS (SAS Inst., Inc., Cary, NC, USA, 1988). Alpha levels of 0.05 (significant) were used in all analysis and the individual tank was considered as the experimental unit.
RESULTS
No mortality was detected for African catfish after 120 days of feeding diets with and without probiotic supplementation; this may be due to the fulfillment of dietary requirement, all the water quality parameters were within the acceptable range for African catfish Clarias gariepinus. The water temperature ranged from 25 to 28°C, Dissolved Oxygen (DO) from 5.0 to 6.5 mg L-1 and pH from 6.0 to 8.0.
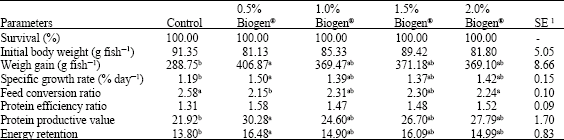
Data in Table 2 shows that African catfish fed diets containing 0.5% Biogen® had a significantly higher (p≤0.05) weight gain (406.87±8.66 g), specific growth rate (1.50±0.15% day-1), protein efficiency ratio (1.58±0.09), protein productive value (30.28±1.70) and energy retention (16.48±0.83) than fish fed control diet. However, among the probiotic fed groups no significant differences were recorded (p≥0.05).
| Table 2: | Growth performance and nutrient utilization of African catfish Clarias gariepinus after 120 days of feeding diets supplemented with Biogen®, Mean in the same row sharing the same superscript are not significantly different (p≥0.05) |
 | |
| 1Pooled standard error; Mean in the same raw sharing the same superscripts are not significant different (p>0.05) | |
| Table 3: | Chemical composition of the whole carcass of African catfish Clarias gariepinus after 120 days of feeding diets supplemented with Biogen®. Mean in the same row sharing the same superscript are not significantly different (p≥0.05) |
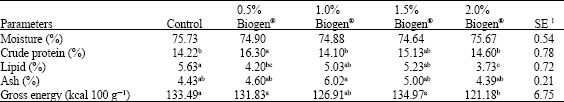 | |
| 1Pooled standard error; Mean in the same raw sharing the same superscripts are not significant different (p>0.05) | |
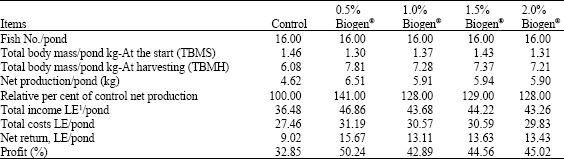
| Table 4: | Fish production and the economic efficiency of the experimental diets |
 | |
| 1One American dollar ($) = 6.0 Egyptian pond (LE) | |
Fish fed control diet without any probiotic had the numerically lowest WG, SGR, PER, PPV and ER. The best-feed conversion ratio was recorded for fish fed diet containing 0.5% Biogen® treatments (2.15±0.10); while the worst FCR was recorded for the fish fed control diet (2.58±0.10).
Concerning the influence of different dietary probiotic levels on chemical proximate analysis of whole carcass (Table 3), no statistical differences (p≥0.05) were observed in the moisture and ash content. Differences were observed in carcass protein content, with the highest significantly (p≤0.05) value recorded in fish fed with 0.5% Biogen® supplement. Carcass lipid and gross energy content was also affected by Biogen® supplemented, the lowest significantly (p≤0.05) lipid and gross energy content were recorded for the fish fed diet supplemented with 2.0% Biogen®.
The higher total net production values were 6.51, 5.91, 5.94 and 5.90 kg pond-1 for fish fed increment levels of Biogen® levels (0.5, 1.0, 1.5 and 2.0%, respectively), while the lowest value’s recorded for fish fed the control diet (4.62 kg pond-1) (Table 4). The same trend was recorded for total pond income (LE) and net pond return (LE). Referring to economic analysis (Table 4) and according to market price in 2006, the input-output analysis indicated that the percentage of 50.24, 42.89, 44.56 and 45.02 were recorded as profit (%) for fish fed diet contents Biogen® at levels of 0.5, 1.0, 1.5 and 2.0%, respectively.
DISCUSSION
All the diets supplemented with Biogen® resulted in growth performances and feed utilization better than that of the control basal diets (Table 2), suggesting that the addition of Biogen® enhance the growth performance and feed utilization of African catfish Clarias gariepinus. These results are in agreement with the findings of Wang and Zirong (2006), who found that the addition of different bacterial strain (photosynthetic bacteria (PSB) and lyophilized Bacillus sp. (B) and their mix) in common carp basal diets improved growth performances, feed utilization and digestive enzyme activities compare to the control diet. In the same trend, bacterial probiotics have been found to influence positively on growth rate in juvenile carp Cyprinus carpio L. (Noh et al., 1994; Bogut et al., 1998). Lara-Flores et al. (2003) observed that, yeast produced the best growth performance and feed efficiency in Nile tilapia Oreochromis niloticus. Kennedy et al. (1998) showed that the addition of a gram-positive probiotic bacterium to diets increased survival and growth rate of marine fish larvae (snook, red drum, spotted seatrout and stripped mullet). In contrast, Gildberg et al. (1995) and Efthimiou (1996) found that there was no effect of probiotics on growth performance was observed in Atlantic salmon fry and dentex, respectively. Also, Shariff et al. (2001) and McIntosh et al. (2000) found that treatment of P. monodon and Litopenaeus vannamei with a commercial Bacillus probiotic did not significantly increase (p≥0.05) either survival or growth. The mechanism by which Biogen® improved growth efficiency in catfish in the current study could be due to the synthesis of a range of relevant digestive enzymes (amylase, protease and lipase). These enzymes would enhance growth performance as a result of higher nutrient digestibility, which could explain the better growth and feed efficiency seen with the supplemented diets. Also, probiotics have been shown to affect on digestive processes by promoting the population of beneficial micro-organism, microbial enzyme activity, improve intestinal microbial balance, consequently improving digestibility and absorption of food and feed utilization (Bomba et al., 2002). Another possible reason for improving growth performance and feed efficiency may be due to the role of probiotics by inhibit the colonization of potential pathogens in the digestive tract by antibiosis or by competition for nutrients and space and alteration of microbial metabolism. Probiotics can also improve nutrition by detoxifying the potentially harmful compounds in feeds, by denaturing the potentially indigestible components in the diet by hydrolytic enzymes including amylases and proteases, by producing vitamins, such as biotin and vitamin B12 (Sugita et al., 1992, 1996; Hoshino et al., 1997), by producing inhibitory compounds (Spanggaard et al., 2001) and by stimulating host immunity (Fuller, 1992; Gibson et al., 1997).
With respect to survival, African catfish survival rate was 100% after 120 days of feeding diets with and without probiotic supplementation; this may be due to the fulfillment of dietary requirement, a healthy fish used in this experiment and the best experimental conditions.
The best-feed efficiency and nutrient utilization including FCR values observed with probiotic-supplemented diets suggest that addition of probiotics improved feed utilization of African catfish Clarias gariepinus. The presence of the significant differences in FCR in present study has been observed in other studies, the other authors found that probiotic improved the growth and feed efficiency of Israeli carp (Noh et al., 1994; Bogut et al., 1998), Nile tilapia (Lara-Flores et al., 2003) and common carp (Wang and Zirong, 2006). The administration of probiotic bacteria and their metabolites has found to be an effective way to promote body weight and feed conversion ratio in farm animals (Abe et al., 1995; Denli et al., 2003).
From a practical point of view, this means that probiotic treatments affect positively on fish growth and increase the feed efficiency, which causing decrease the amount of feed necessary for optimum growth, which could result in reducing production cost. Improving FCR may be due to synthesis of digestive enzymes in Biogen® treatments which led to enhanced digestion and increased absorption of food, which in turn contributed to the improved feed efficiency. In contrast, other studies have been observed there was no significant difference in feed conversion ratio (Efthimiou et al., 1994; Gildberg et al., 1995; Efthimiou, 1996; Gildberg and Mikkelsen, 1998; Hidalgo et al., 2006).
Significant differences were observed in PER, PPV and ER results indicate that supplementing diets with probiotics improves protein and energy utilization in African catfish. Better efficiency of protein uptake for the probiotic groups may be due to better digestion, absorption and assimilation of the nutrients. This results may be supported by the postulation from Lara-Flores et al. (2003) who found that the PER and ANU recorded the best values with probiotic-supplemented diets. In contrast, Hidalgo et al. (2006) found that no significant differences were observed in PER and PPV for juvenile dentex fed two probiotics (Bacillus toyoi, T and B. cereus, E) at increasing levels (0.5, 1 and 2 g kg-1 diet).
Yields of African catfish fed diets supplemented with 0.5, 1, 1.5 and 2% Biogen® resulted in 141, 128, 129 and 128% more production than the control diet, respectively (Table 4). African catfish seems to be more efficiently utilizing the nutrient when supplemented with probiotics. Meanwhile, the least net yield was recorded with the fish fed the control diet (without probiotic). Referring to the economic analysis (Table 4) and according to the market price in 2006, the total net return of fish production using probiotic at four levels from 0.5 to 2% were higher than the control. However, the cost-benefit analyses of the present study clearly indicated that the Biogen® diets are better for African catfish Clarias gariepinus than the control diet.
It could be concluded that the addition of probiotic Biogen® as a feed additive in African catfish Clarias gariepinus diets is recommended to improve growth performance and nutrient utilization as well as improve the economic efficiency.
ACKNOWLEDGMENTS
The authors are grateful to the Middle East land Reclamation Company (MELARC) for the supply of the ingredients and the fish farm used in this study. The authors wish to thank the Department of Animal Production staff and technicians at Faculty of Agriculture, Cairo University for their technical facilities providing.
REFERENCES
- Abe, F., N. Ishibashi and S. Shimamura, 1995. Effect of administration of Bifidobacteria and lactic acid bacteria to newborn calves and piglets. J. Dairy Sci., 78: 2838-2846.
CrossRefDirect Link - AOAC., 1995. Official Methods of Analysis. 16th Edn., Association of Official Analytical Chemists, Washington, DC., USA.
Direct Link - Bligh, E.G. and W.J. Dyer, 1959. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol., 37: 911-917.
CrossRefPubMedDirect Link - Bogut, I., Z. Milakovic, Z.I. Bukvic, S. Brkic and R. Zimmer, 1998. Influence of probiotic (Streptococcus faecium M74) on growth and content of intestinal microflora in carp (Cyprinus carpio). Czech J. Anim. Sci., 43: 231-235.
Direct Link - Chabrillon, M., R. Rico, M.C. Balebona and M.A. Morinigo, 2005. Adhesion to sole (Solea senegalensis) mucus of microorganisms isolated from farmed fish and their interaction with Photobacterium damselae subsp. piscicida. J. Fish Dis., 28: 229-237.
PubMedDirect Link - Denli, M., F. Okan and K. Celik, 2003. Effect of dietary probiotic, organic acid and antibiotic supplementation to diets on broiler performance and carcass yield. Pak. J. Nutr., 2: 89-91.
CrossRefDirect Link - Efthimiou, S., P. Divanach and H. Rosenthal, 1994. Growth, food conversion and agonistic behaviour in common dentex (Dentex dentex) juveniles fed on pelleted moist and dry diets. Aquat. Living Resour., 7: 267-275.
CrossRefDirect Link - Gatesoupe, F.J., 1999. The use of probiotics in aquaculture. Aquaculture, 180: 147-165.
CrossRefDirect Link - Gibson, G.R., J.M. Saavendra, S. MacFarlane and G.T. MacFarlane, 1997. Probiotics and Intestinal Infections. In: Probiotics 2, Application and Practical Aspects, Fuller, R. (Ed.). Chapman and Hall, London, UK., pp: 10-39.
Direct Link - Gildberg, A., A. Johansen and J. Bogwald, 1995. Growth and survival of Atlantic salmon (Salmo salar) fry given diets supplemented with fish protein hydrolysate and lactic acid bacteria during a challenge trial with Aeromonas salmonicida. Aquaculture, 138: 23-34.
Direct Link - Gildberg, A. and H. Mikkelsen, 1998. Effects of supplementing the feed to Atlantic cod (Gadus morhua) fry with lactic acid bacteria and immuno-stimulating peptides during a challenge trial with Vibrio anguillarum. Aquaculture, 167: 103-113.
CrossRefDirect Link - Haylor, G.S., 1993. Aspects of Biology and Culture of the African Catfish Clarias gariepinus (Burchell, 1822) with Particular Reference to Developing African Countries. In: Recent Advances in Aquaculture, Muir, J.F. and R.J. Roberts (Eds.). Vol. IV, Blackwell Scientific Publications, Oxford, pp: 235-294.
- Hidalgo, M.C., A. Skalli, E. Abellan, M. Arizcun and G. Cardenete, 2006. Dietary intake of probiotics and maslinic acid in juvenile dentex (Dentex dentex L.): Effects on growth performance, survival and liver proteolytic activities. Aquac. Nutr., 12: 256-266.
CrossRefDirect Link - Hjelm, M., O. Bergh, A. Riaza, J. Nielsen and J. Melchiorsen et al., 2004. Selection and identification of autochthonous potential probiotic bacteria from turbot larvae (Scophthalmus maximus) rearing units. Syst. Applied Microbiol., 27: 360-371.
CrossRefDirect Link - Hoshino, T., K. Ishizaki, T. Sakamoto, H. Kumeta, I. Yumoto, H. Matsuyama and S. Ohgiya, 1997. Isolation of a Pseudomonas species from fish intestine that produces a protease active at low temperature. Lett. Applied Microbiol., 25: 70-72.
Direct Link - Huisman, E.A. and C.J.J. Richter, 1987. Reproduction, growth, health control and aquacultural potential of the African catfish, Clarias gariepinus (Burchell 1822). Aquaculture, 63: 1-14.
CrossRefDirect Link - Kennedy, S.B., J.W. Tucker, C.L. Neidig, G.K. Vermeer, V.R. Cooper, J.L. Jarrell and D.G. Sennett, 1998. Bacterial management strategies for stock enhancement of warm water marine fish: A case study with common snook (Centropomus undecimalis). Bull. Mar. Sci., 62: 573-588.
Direct Link - Lara-Flores, M., M.A. Olvera-Novoa, B.E. Guzman-Mendez and W. Lopez-Madrid, 2003. Use of Streptococcus faecium and Lactobacillus acidophilus and the yeast Saccharomyces cerevisiae as growth promoters in Nile tilapia (Oreochromis niloticus). Aquaculture, 216: 193-201.
Direct Link - Lategan, M.J., F.R. Torpy and L.F. Gibson, 2004. Control of saprolegniosis in the eel Anguilla australis Richardson, by Aeromonas media strain A199. Aquaculture, 240: 19-27.
CrossRefDirect Link - McIntosh, D., T.M. Samocha, E.R. Jones, A.L. Lawrence, D.A. McKee, S. Horowitz and A. Horowitz, 2000. The effect of a commercial bacterial supplement on the high-density culturing of Litopenaeus vannamei with a low-protein diet in an outdoor tank system and no water exchange. Aquacult. Eng., 21: 215-227.
Direct Link - Nikoskelainen, S., A.C. Ouwehand, G. Bylund, S. Salminen and E.M. Lilius, 2003. Immune enhancement in rainbow trout (Oncorhynchus mykiss) by potential probiotic bacteria (Lactobacillus rhamnosus). Fish Shellfish Immunol., 15: 443-452.
CrossRefPubMedDirect Link - Picchietti, S., M. Mazzini, A.R. Taddei, R. Renna and A.M. Fausto et al., 2007. Effects of administration of probiotic strains on GALT of larval gilthead seabream: Immunohistochemical and ultrastructural studies. Fish Shellfish Immunol., 22: 57-67.
CrossRefDirect Link - Robertson, P.A.W., C. O`Dowd, C. Burrells, P. Williams and B. Austin, 2000. Use of Carnobacterium sp. as a probiotic for Atlantic salmon (Salmo salar L.) and rainbow trout (Oncorhynchus mykiss, Walbaum). Aquaculture, 185: 235-243.
Direct Link - Shariff, M., F.M. Yusoff, T.N. Devaraja and P.S.S. Rao, 2001. The effectiveness of a commercial microbial product in poorly prepared tiger shrimp, Penaeus monodon (Fabricius), ponds. Aquacult. Res., 32: 181-187.
CrossRefDirect Link - Spanggaard, B., I. Huber, J. Nielsen, E.B. Sick and C.B. Pipper et al., 2001. The probiotic potential against vibriosis of the indigenous microflora of rainbow trout. Environ. Microbiol., 3: 755-765.
CrossRefDirect Link - Sugita, H., J. Kawasaki, J. Kumazawa and Y. Deguchi, 1996. Production of amylase by the intestinal bacteria of Japanese coastal animals. Lett. Applied Microbiol., 23: 174-178.
CrossRefDirect Link - Teuber, M., 2001. Veterinary use and antibiotic resistance. Curr. Opin. Microbiol., 4: 493-499.
Direct Link - Verreth, J., E.H. Eding, G.R.M. Rao, F. Huskens and H. Segner, 1993. A review of feeding practices, growth and nutritional physiology in larvae of the catfishes Clarias gariepinus and Clarias batrachus. J. World Aquac. Soc., 24: 135-144.
CrossRef - Wang, Y. and Z. Xu, 2006. Effect of probiotics for common carp (Cyprinus carpio) based\on growth performance and digestive enzyme activities. Anim. Feed Sci. Technol., 127: 283-292.
CrossRefDirect Link
folorunso adebayo Reply
pls where can i buy probiotic biogen in nigeria because i am catfish farmer thanks.
emmanuel oghai Reply
i am atlso inetrested in where to get probiotic biogen, i am besed in Port Harcourt