
Ahmad Qasaimeh
Department of Civil Engineering, Jadara University, Jordan
Mohammad Raji Abdallah/Qasaimeh
Department of Chemical Engineering, Al-Huson University College, Al-Balqa� Applied University, Salt, Jordan
Departments of Civil and Environmental Engineering, College of Engineering at Al Lith, Umm Al-Qura University, Saudi Arabia
Falah Bani Hani
Department of Chemical Engineering, Al-Huson University College, Al-Balqa� Applied University, Salt, Jordan
Journal of Environmental Science and Technology
Year: 2016 | Volume: 9 | Issue: 1 | Page No.: 1-25







ABSTRACT
Biogas in landfill is being captured by natural and engineered processes. The natural processes are represented by biological activities such as bacterial methane oxidation and plant uptake for carbon dioxide at topsoil layer. Landfill gas is transported through soil layers in landfill top or in nearby areas before being released to the atmosphere. Whilst transported in the soil layers the biogas is mixed with atmospheric air and the methane may hence be oxidized by the methanotrophic bacteria in the soil using oxygen from atmosphere. Methane oxidation is affected by different environmental factors such as; temperature, water content, nutrients, substrate and oxygen concentrations. One of the ways to decrease greenhouse emissions in the future is to plant fast growing woody crops thereby sequestering carbon and displacing fossil fuels by harvesting woody biomass for bio-energy, or by storing carbon in long-lived woody products. Plant uptake for carbon dioxide is affected by some parameters such as; CO2 concentration, nitrogen concentration, water content and temperature. The engineered processes are represented by various physical biogas extractions; gas is collected using network of collection pipes and wells. The gas collection efficiency in landfills is between 40-90%. Landfill gas can be collected by either a passive or an active collection system. Passive gas collection systems use existing variations in landfill pressure and gas concentrations to vent landfill gas into the atmosphere or a control system. Active gas collection is considered a good means of landfill gas collection. An active collection system composed of extraction wells connected to header pipe to a pump that delivers gas for energy recovery.
PDF Abstract XML References Citation
Received: September 21, 2015;
Accepted: November 03, 2015;
Published: November 19, 2015
How to cite this article
Ahmad Qasaimeh, Mohammad Raji Abdallah/Qasaimeh and Falah Bani Hani, 2016. A Review on Biogas Interception Processes in Municipal Landfill. Journal of Environmental Science and Technology, 9: 1-25.
DOI: 10.3923/jest.2016.1.25
URL: https://scialert.net/abstract/?doi=jest.2016.1.25
DOI: 10.3923/jest.2016.1.25
URL: https://scialert.net/abstract/?doi=jest.2016.1.25
INTRODUCTION
Solid waste at landfills is a free source of energy, biogas is generated naturally by bacterial activity and thus municipal solid waste is recently manipulated via several approaches that entail sustainable management systems, techniques, models, policies and strategies (Liamsanguan and Gheewala, 2008; Rada et al., 2009; Ionescu et al., 2013; Vaccari et al., 2012).
The biodegradable municipal solid waste material is made up of vegetation, domestic matter, paper, garden waste, wood and textile material. The organic carbon is the common element between biodegradable materials that allows the development of energy and methane production. On previously mentioned bases, municipal waste is being investigated by many studies (Trulli et al., 2013; Chanakya et al., 2009; Rada et al., 2013).
In land filled waste, the biochemical reactions go through several phases: hydrolysis phase, an acidogenesis phase and the methanogenesis, which is the main source of methane contained in the final biogas mixture. Biological processes under anaerobic conditions drive the production of biogas, a gaseous mixture composed almost of methane (CH4) and carbon dioxide (CO2). Anaerobic bacteria is responsible of the decomposition of organic material (Davis and Cornwell, 2008).
Landfill gas production rate is influenced by functional parameters such as abiotic factors and landfill operation procedures (Christensen et al., 1996). The abiotic factors of concern can be summarized as: pH, nutrients, inhibitors, temperature and water content, while the landfill operation procedures of concern can be summarized as; waste composition, sewage sludge addition, shredding, compaction, soil cover, recirculation of leachate and pre-composting.
The methane producing bacteria is called methanogens which operate within 6-8 pH in methanogenesis process. Anaerobic ecosystem requires much less nitrogen (N2) and phosphorous (P) than the aerobic system does. The optimal ratio for organic matter (as COD), N2 and P content, is 100:0.44:0.08 (Christensen et al., 1996). Nutrients and metal supplementation have a positive effect on biogas production rate (Isci and Demirer, 2007). An adequate ratio of nitrogen (N) and phosphorus (P) is also required by the anaerobic ecosystem (Zhang et al., 2014).
The temperature for mesophilic bacteria is in the range of 30-35°C, while 45-65°C is for the thermophilic bacteria. The active temperature for methanogens is in the range of 30-50°C. The optimum temperature range of gas generation is between 30-45°C during the main landfill gas generation phase (Williams, 2005). The change of temperature will have an impact on the growth and the activity of the microorganisms (Mora-Naranjo et al., 2004). The range of moisture content in a typical landfill is 15-40% with a typical average of 30% (Williams, 2005). The moisture content controls methane production, since it stimulates microbial activity by providing closer contact between soluble and insoluble substrates and bacteria (Machadoa et al., 2009).
There are two main ways to capture biogas emission from landfills. One, like those in Northern Europe where there are many small and old landfills with low gas generation that is biologically uptaken (Jones and Nedwell, 1993; Boeckx et al., 1996; Borjesson and Svensson, 1997), where biological biogas uptake at landfills is apparently executed by biological methane oxidation by methanotrophs and carbon dioxide uptake by plants. Other option is gas collection and utilization, which could be very effective with a high gas generation. In this case, biogas can be collected by means of vertical and horizontal drain pipes and is employed to produce heat and energy. There are different technologies being studied to find the best handling of biogas collection (Andreottola and Cossu, 1988; EMCON Associates, 1980; El-Fadel, 1991).
BIOLOGICAL METHANE OXIDATION
Methane has Global Warming Potential (GWP) 21 times more than carbon dioxide. Atmospheric methane concentration has doubled during the last 100 years and is an explosive gas in concentrations between 5 and 15% in air, a 10% of global anthropogenic source of methane is from MSW landfills (Chalvatzaki and Lazaridis, 2009; Zhang et al., 2008).
While soils have not been considered as significant sinks for methane until recently, methane consumption has been reported in agricultural soils, forest soils, tundra and bogs (Topp and Hanson, 1991). Biological oxidation of CH4 by bacterial methanotrophs has attracted much attention from the research community as an inexpensive waste gas treatment mechanism. Methane oxidizing activity, with a decrease in soil oxygen and an increase in microbial biomass, has been demonstrated in soils around leaks in natural gas pipes (Adams and Ellis, 1960) and in landfill covers (Kightley et al., 1995; Whalen et al., 1990; Bogner et al., 1995). Methane oxidizing activity in soils is an event that could have a strong effect on CH4 emissions control from sources such as municipal landfills and the optimization of this process may give out an inexpensive strategy for controlling and utilizing emissions of this potent greenhouse gas.
Microbial oxidation in well-drained soils is the only identified biological sink for atmospheric CH4 and accounts for 3-9% of total annual atmospheric CH4 destruction (Prather et al., 1996). This is similar in magnitude to the current atmospheric increase (Houghton et al., 1996). Accordingly, alterations of the soil sink strength are a significant determinant of the rate of change in the atmospheric CH4 concentration (Prather et al., 1996) and absence of this sink will cause the atmospheric CH4 concentration to increase at 1.5 times the current rate (Duxbury, 1994). The control of aerobic methane oxidation is obviously related to the requirement for oxygen and methane. As a result, maximum oxidation rates are found where diffusion of oxygen from above and of methane from below is optimal for methanotrophs (King, 1992; Sundh et al., 1995a, b).
Xin et al. (2004) showed that methanotrophs oxidizes methane to carbon dioxide through sequential reactions catalyzed by a series of enzymes including methane monooxygenase, methanol dehydrogenase, formaldehyde dehydrogenase and formate dehydrogenase. Methanotrophic bacteria cultivate aerobically on methane as a sole source of carbon and energy. The first two enzymes involved in methane oxidation are methane monooxygenase (MMO) and methanol dehydrogenase (MDH) (Anthony, 1986). The MMO oxidizes methane to methanol and MDH catalyzes the oxidation of methanol to formaldehyde. Methylosinus trichosporium OB3b is a methanotrophic bacterium and contains two forms of MMOs: a soluble (sMMO) and a membrane-bound particulate (pMMO) whose syntheses depend on growth conditions (Murrell et al., 2000; Nielsen et al., 1997; Takeguchi and Okura, 2000).
Methane emission rates were strongly dependent on the oxidation coefficient utilized, which varies with the cover material and microclimate conditions of the site (Moreira et al., 2015). The maximum methane oxidation rates values for compost cover layer is higher than regular soil cover (Abichou et al., 2009). Some experiments in the literature such as, Im et al. (2009), Abichou et al. (2009), Scheutz et al. (2008, 2009) and Scheutz and Kjeldsen (2004) show that methane oxidation rate in landfill covers obtained in field and laboratory measurements usually formatted as parameters follow Michaelis-Menten kinetics equation and this present very different rates of methane oxidation under different site and microclimatic conditions. Methane oxidation rates depend upon different variables such as microclimate conditions, temperature, atmospheric pressure, moisture, soil conditions, oxygen concentration, methane concentration and amount of methanotrophs (Spokas and Bogner, 2011; Chanton et al., 2011).
Landfill gas is transported through soil layers in landfill top covers or in nearby areas before being released to the atmosphere. Whilst transported in the soil layers the biogas is mixed with atmospheric air and the methane may hence be oxidized by the methanotrophic bacteria in the soil using oxygen from atmosphere. Methane oxidation is affected by different environmental factors such as: temperature, water content, nutrients, substrate and oxygen concentrations (Hanson and Hanson, 1996). In the following sections is the description of environmental factors effect on methane oxidation.
Effect of soil moisture: Christophersen et al. (2000) showed that methane oxidation rate is a function of soil moisture content at different temperatures for the different soils. The optimum soil moisture content was different for each soil and depended on the temperature. Some soils had the lowest optimum soil moisture content and others had the highest. At lower temperatures the difference in oxidation rates with soil moisture content was reduced.
Methane oxidation rates decreased extensively after soil samples were dried below field moisture contents, increased to an optimum value as water was added and decreased with sustaining water addition. The maximum oxidation rate occurred at moisture content of 15.4% (dry weight basis) (Stein and Hettiaratchi, 2001). The texture and structure of soil will influence its moisture content in a manner that is site specific, depending on climatic variables such as temperature, solar flux, average wind speed and the type of vegetative cover (Stein and Hettiaratchi, 2001). Methane uptake was controlled strongly by soil moisture, with reduced fluxes under conditions of very low or very high soil moisture contents. The mineral soil Q10 (Q10 is the value for how many times the oxidation rate increases when temperature is increased 10°C at temperatures below the optimum temperature) of 1.11 for CH4 uptake indicates that methane uptake is controlled primarily by physical processes (Bowden et al., 1998).
Effect of temperature: Christophersen et al. (2000) showed for all the soil investigations, the oxidation rate increased with increasing temperature. Predictably, optimum temperatures were not found in this experimentation. For all the soils the optimum temperature must be higher than 15°C, which was the highest temperature in these explorations. Most researchers found optimum temperatures around 30°C, which do seldom occur in temperate soils. Dunfield et al. (1993) found optimum temperature for the methane oxidation around 25°C. As the temperature is increased, CH4 oxidation increases exponentially to maximum and then decreases with continued temperature increase (Stein and Hettiaratchi, 2001).
At high methane concentrations the oxidation becomes saturated and the rate-limiting stage is the enzymatic action. Thus, the temperature response is something like parabolic: increasing rates with increasing temperature to a maximum and declining with continued temperature increase (Bailey and Ollis, 1986). King and Adamsen (1992) investigated soil cores where the temperature was increased between 0 and 30°C. The depth distribution of methane consumption and methane diffusion showed low sensitivity to changes in temperature. They observed methane consumption at -1°C and they suggested that methane consumption might occur at low temperatures on condition that the soil water remains liquid. Sommerfeld et al. (1993) showed that the soil microflora was active even when the soil was snow-covered and near 0°C and that methane consumption was taking place under that condition. In the 0-10°C range methane oxidation was about 13-38% of maximum activity. Prieme and Christensen (1997) observed methane oxidation to be active at low temperatures, down to 1°C in the field and -2°C in soil core experiments. Both in the field and in soil cores similar temperature responses of methane oxidation were measured. This indicated that temperature acts directly (i.e., via its effect on enzymatic processes and methane diffusion) on methane oxidation in the field. They suggested that the small temperature response of methane oxidation was partly a result of low substrate concentration.
Effect of organic matter content and methane concentration: Oxidation rates increased with increasing organic matter content. The optimum soil moisture content also increased with increasing organic matter content (Christophersen et al., 2000). Oxidation of methane in top covers of landfills has been observed on several occasions and soil exposed to elevated methane concentrations can develop a high potential for methane oxidation (Whalen et al., 1990; Kightley et al., 1995; Boeckx et al., 1996; Czepiel et al., 1996; Borjesson and Svensson, 1997; Bogner et al., 1997). Low initial methane concentrations resulted in low maximal oxidation rates (Boeckx and van Cleemput, 1996; Boeckx et al., 1996). The oxidation rates at low initial methane concentrations were much lower than oxidation rates at high initial methane concentrations and the oxidation was performed by different kinds of bacteria (Bender and Conrad, 1992). Several researchers have shown that different species of bacteria are active at low and high methane concentrations (Bender and Conrad, 1994; Kightley et al., 1995; Bogner et al., 1997).
Effect of nitrogen content: There is a worldwide increase in atmospheric nitrogen (N) deposition on terrestrial and aquatic environments (Matthews, 1994; Galloway et al., 1995). Methanotrophs are inhibited by high soil N; driving attention that the contemporary worldwide increase in atmospheric N deposition will decrease soil CH4 oxidation. Oxidation of CH4 by methanotrophic and methylotrophic bacteria occurs in aerobic soils and the magnitude and rate of oxidation are influenced by soil type, aeration, environmental parameters and Nitrogen availability (Topp and Pattey, 1997; Le Mer and Roger, 2001). Application of fertilizer has been shown to inhibit CH4 oxidation in soil (Steudler et al., 1989; Hutsch, 1998; Tlustos et al., 1998; Kravchenko et al., 2002). Nitrogen content frequently shows low atmospheric CH4 oxidation comparative to unfertilized controls (Steudler et al., 1989; King and Schnell, 1994; Sitaula et al., 1995).
In two marshes, the vertical distribution of methane oxidation in the sediment and methane oxidation inhibition by ammonium was investigated by Van Der Nat et al. (1997). In a slurry incubation experiment, he conducted tests for two sites different in their prevailing vegetation type, i.e., reed and bulrush and in their heights above sea level. Inhibition of methane oxidation by ammonium was observed in all samples and depended on methane and ammonium concentrations. Increasing ammonium concentrations resulted in more inhibition and increasing methane concentrations resulted in less inhibition. Increasing atmospheric concentrations of CO2 may increase emissions of N2O by denitrification and either increase or decrease the ability of soil to buffer atmospheric CH4 depending on fertilizer application (Baggs and Blum, 2004).
Effect of leachate recirculation, metals and minerals: Leachate recirculation is one option for inexpensive leachate disposal (Kinman et al., 1987; Cureton et al., 1991), in reducing the cost of post-closure care and long-term liability (Diamadopoulos, 1994; Westlake, 1995; Reinhart and Al-Yousfi 1996). It could participate to improve leachate quality; reduce volume of leachate to be treated and enhance gas production (Reinhart, 1996; Sulisti et al., 1996; Warith et al., 1999). Chan et al. (2002) found that leachate recirculation reduced waste stabilization time and was effective in enhancing gas production and improving leachate quality, especially in terms of Chemical Oxygen Demand (COD). The results also indicated that leachate recirculation could maximize the efficiency and waste volume reduction rate of landfill sites. Leachate recirculation gives an aqueous environment that assists the supply of nutrients and biomass within the landfill that stimulates the degradation of municipal solid waste (El-Fadel, 1999). Mobilizing nutrients and microorganisms in aqueous wastes improves mass transfer to prevent the development of inactive zones in landfill zones (Chugh et al., 1998). Leachate thus provides supply of COD, nutrients, biomass and water content with dissolved oxygen which stimulates methane oxidation.
Maurice et al. (1999) showed that larger trees arise on plots irrigated with leachate, presumably due to the positive effect of water and nutrient supply. Methane oxidation levels between 50 and 950 mol m–2 year were observed. The positive relationship between soil methane oxidation capacity and tree existence is important for reduction of methane emission by landfill vegetation type. Optimizing methane oxidation using vegetation on topsoil could reduce the amount of methane released to the atmosphere (Maurice et al., 1999). Leachate provides the soils with higher content of water, nutrient and organic matter, it also provides metals. Heavy metals affect the growth, morphology and metabolism of microorganisms of soils through functional disturbance, protein denaturation, or the destruction of the integrity of the cell membrane (Baath, 1989; Babich et al., 1980; Leita et al., 1995). In a laboratory incubation study, Mishra et al. (1999) showed that selected heavy metals in three rice soils were different in their effect on methanogenesis and methane-producing bacteria. The Cd, Cu and Pb inhibited CH4 production in all soils. The Zn stimulated CH4 production in the alluvial soil, but inhibited it in laterite and acid sulfate soils. The Cr effectively inhibited CH4 production in the alluvial soil, but stimulated it in laterite and acid sulfate soils (Mishra et al., 1999).
In a laboratory study, Mohanty et al. (2000) showed that selected heavy metals differed in their effect on CH4 oxidation in two soils for two water systems. The Cr significantly inhibited CH4 oxidation in alluvial soil at 60% moisture capacity, while Cu stimulated the process. On the other hand, Zn inhibited CH4 oxidation in both alluvial and laterite soils under saturated conditions.
The effect of inorganic redox substances (species of NO3‾, Mn4+, Fe3+ and SO4–2) on methane production and oxidation in anoxic rice soil samples has vital effects. Sulfate was the most inhibiting for methane production followed by Fe3+, NO3‾ and Mn4+, respectively. Laboratory studies showed that the addition of MnO2 and K2SO4 enhanced aerobic methane oxidation in soil samples at 60% water content. Nitrate and Fe3+ motivated methane oxidation under anaerobic conditions and delayed it under aerobic conditions. Manganese (IV) delayed methane oxidation under anaerobic conditions, but enhanced it under aerobic conditions. On the other hand, SO4–2 stimulated methane oxidation in soil equivalent medium under both aerobic and anaerobic conditions (Kumaraswamy et al., 2001).
There is substantial interest in methane monooxygenase (MMOs) of methanotrophic bacteria in soils, because these enzymes in methanotrophs oxidize methane to a potential fuel source, methanol, detoxifies trichloroethylene and uses a greenhouse gas as a reactant. It is well known that sMMO expression in methanotrophs is repressed by copper ions sMMO is expressed for concentrations lower than 0.86 mmol g–1 dry cell weight (Barta and Hanson, 1993) or, generally, when the copper concentration is lower than 1 mM (Burrows et al., 1984). Above this concentration, the particulate (membrane-bound) form of methane monooxygenase (pMMO) is produced (Nguyen et al., 1994). To overcome this natural regulation, a M. trichosporium OB3b mutant has been obtained that expresses sMMO in the presence of copper probably because of a deficiency in copper transport (Phelps et al., 1992). Transcription of soluble methane monooxygenase (sMMO) of methanotrophs is tightly regulated by low concentrations of copper ions [Cu2+ e.g., transcription is completely repressed at copper concentrations higher than 0.86 μmol g–1 dry cell weight] (Green et al., 1985). In a research study, Jahng and Wood (1996) showed sMMO inhibition by metal ions and different medium ingredients was investigated for the first time using sMMO purified from the type II methanotroph Methylosinus trichosporium OB3b. Cu(I) and Cu(II) decreased sMMO activity of Methylosinus trichosporium OB3b by inhibiting not only the reductase but the hydroxylase component as well. Ni (II) also inhibited both enzyme components and Zn (II) inhibited sMMO by lowering the activity of the hydroxylase only. The Ni (II) and Zn (II) aggregated the reductase component of sMMO and the later precipitated the hydroxylase component. Cu (II) caused the reductase to precipitate (Jahng and Wood, 1996).
Uncertain environmental factors: Field studies in temperate forests have shown a chronological illustration of CO2 and CH4 fluxes (Castro et al., 1994, 1995; Peterjohn et al., 1994) that corresponds strongly with seasonal changes in soil moisture and temperature. Temperature is considered the primary predictor of CO2 fluxes, not surprisingly; moisture also influences soil respiration rates (Groffman et al., 1992). Moisture usually exerts strong control over CH4 uptake rates, although inclusion of both moisture and temperature in models can increase predictive capabilities. Lessard et al. (1994) suggested that the strong relationship between moisture and CH4 uptake may mask relationships between temperature and uptake, thus it has been difficult to determine the relative importance of these factors. Steinkamp et al. (2001) showed significant seasonal differences in the magnitude of CH4 oxidation rates at experimental sites with high rates during summer, relative low rates during winter and intermediate rates during spring and autumn. Hellebrand and Scholz (2000) showed results that the temperature was the main reason for the seasonal change of the methane uptake. Whereas the uptake dropped near to zero during the winter period, the uptake rates reached values up to 0.6 mg CH4 m–2 day–1 (25 μg CH4 m–2 h–1) in the summer. Lessard et al. (1994) used field studies to determine the relative importance of moisture and temperature in controlling flux rates is difficult because soil temperature and moisture usually vary seasonally in temperate ecosystems. Soil temperatures are usually highest by late summer, but strong evapotranspiration potentials usually reduce soil water even if precipitation stays relatively constant. Thus, it is not a straightforward exercise to determine if maximum rates of soil respiration or CH4 uptake in late summer, for example, are due to high temperatures, lower soil moisture, or an interaction of both factors.
BIOLOGICAL PLANT UPTAKE OF CARBON DIOXIDE
Measurements of CO2 in the atmosphere, which began in Maunaloa (Hawaii) in 1958, indicated clearly that the concentration of CO2 in the atmosphere is increasing rapidly (Keeling et al., 1982). The ice core studies showed that the CO2 concentration was about 205 μmol mol–1 some 20,000 years ago. Pre-industrial value was 280 mmol mol–1 during the past 10,000 years. Whereas, before 1900, the CO2 concentration in the atmosphere was 290 mmol mol–1. Maunaloa studies also observed that from 1958-1982 there was an increase of 1.0 mmol mol–1 CO2 per year. The 1958 value of atmospheric CO2 was 316 mmol mol–1 and the present concentration were recorded as high as 370 mmol mol–1 (Kimball, 1997). Thus the concentration of CO2 in the atmosphere is likely to be doubled (600 mmol mol–1) by the middle of 21st century (Houghton et al., 1990). Recent studies suggest that boreal forests may play major role in regulating the climate of the northern hemisphere and in global carbon cycling (Bonan et al., 1992). In North America, the Boreal ecosystem atmosphere study program has investigated carbon and energy exchange in two regions of Canada (Sellers et al., 1995).
Short rotation woody crops fix carbon dioxide from the atmosphere and store carbon both above and belowground as biomass. Moreover, the harvested portions of the trees displace other products that are made from non-renewable fossil fuels (Tuskan and Walsh, 2001). One of the way to decrease greenhouse emissions in the future is to plant fast growing woody crops on unproductive land thereby sequestering carbon and displacing fossil fuels by harvesting woody biomass for bio-energy, or by storing carbon in long-lived woody products (Tuskan and Walsh, 2001). Short rotation poplar and willow crops deployed as phytoremediation buffer systems would qualify for carbon uptake and if planted at large scales would contribute greatly to atmospheric carbon dioxide gas reduction (Isebrands and Karnosky, 2001).
Examples of CO2 plant uptake: Plants in their juvenile phase can benefit more than mature ones of optimal growing conditions. Transplant production in greenhouses offers the opportunity to optimize growing factors in order to reduce production time and improve transplant quality. Carbon dioxide and light are the two driving forces of photosynthesis. Carbon dioxide concentration can be enriched in the greenhouse atmosphere, leading to heavier transplants with thicker leaves and reduced transpiration rates (Tremblay and Gosselin, 1998). The almost entirely juvenile tissues of seedlings are all expanding and could be utilizing and diluting the enhanced photosynthate production in an enriched CO2 atmosphere (Lindhout and Pet, 1990). Hence, the greatest advantage of CO2 enrichment would be realized in the vegetative growth of young plants (Kimball, 1983). As leaf tissues formed early in seedling culture begin to mature, starch accumulation begins to slow photosynthetic rates and relative growth rate (Thomas et al., 1975).
The potential benefit from CO2 augmentation of the greenhouse atmosphere has been known for a long time (Hand, 1984; Enoch, 1990) and they are particularly important for roses (Urban, 1994; Baille et al., 1996). The response of "Parfum de Rose" to CO2 augmentation is rapid: it takes only 2 months to observe a sharp yield increase.
The net primary production of maturing loblolly pine (Pinus taeda), on a site of moderate soil fertility, increased by 25% in the two years following the onset of a 200 ppm CO2 enrichment in a ‘Free Air CO2 Enrichment’ FACE experiment (DeLucia et al., 1999). A parallel longer-term study produced a larger stimulation of tree growth of up to 34% over the first three years of enrichment (Oren et al., 2001). It is particularly interesting; therefore, that carbon allocation to reproduction is strongly stimulated in loblolly pine after three years of CO2 enrichment (LaDeau and Clark, 2001). After this period, trees growing in the enriched CO2 were twice as likely to be reproductively mature and produced three times as many cones and seeds, than control trees. In contrast, flowering and seed set in grasslands, where species may have deterministic life cycles, were stimulated (Smith et al., 2000).
C4 and C3 species increased total biomass significantly in elevated CO2 by 33 and 44%, respectively and higher carbon assimilation rates were found in both C3 and C4 grass 33 and 25%, respectively (Wand et al., 1999). Carbon dioxide enrichment increased transplant leaf area, shoot and root dry weight and decreased the leaf area ratio of celery (Apium graveolens L.) transplants (Tremblay et al., 1987). Study on the characterization of CO2 responsiveness in Brassica oxycamp hybrid and its parents B. oxyrrhina and B. campestris showed that the response of B. oxycamp hybrid to elevated CO2 was significantly positive in respect to photosynthesis and growth, similar to that of its parent B. campestris. Hybrid B. oxycamp and B. campestris with greater sink potential responded significantly, whereas, B. oxyrrhina with poor sink size did not respond to CO2 enrichment (Uprety et al., 1998).
Woodrow et al. (1987) demonstrated that CO2 affects both source metabolism and partitioning to sinks (stems, roots and leaf carbohydrate) in tomato plantlets. They found that CO2 enrichment produced heavier transplants desirable for successful field establishment without elongation growth. Dry matter accumulation in shoot and root was increased as well as leaf dry weight (by 81% over control). Transpiration rates were reduced under CO2 enrichment conditions by 34%. Increased leaf dry weight accumulation and Specific Leaf Weight (SLW) under CO2 enrichment suggests that more carbohydrate may be available to the plant for future growth. Apparently, the ratio of total sugars to amino acids in the leaf is shifted in favor of sugar content. In a study with tree seedlings, Luxmoore et al. (1986) suggest that CO2 enrichment may increase sucrose translocation in roots and facilitate the mobilization of N and C compounds to new root primordia. Increased net leaf photosynthesis rate and decreased transpiration rate under CO2 enrichment are well documented (Woodrow et al., 1987). One of the most important effects of CO2 enrichment is the increase in water efficiency (Wong, 1979), which leads to drought tolerance. Actually, rising CO2 concentration reduces the transpiration of plants by 20-40% (Mortensen, 1987). Radoglou et al. (1992) reported an increase in water use efficiency of bean (Phaseolus vulgaris L.) leaves as a result of increased assimilation rate and decreased stomatal conductance at higher ambient CO2 concentrations. In cotton (Gossypium arborcum L.), stomata conductance after 40 day of CO2 enrichment took 5 d to reach normal levels in non-enriched conditions. In short, CO2 enrichment of vegetable transplants shortens the nursery period and modifies photosynthate allocation to the diverse parts, leading to sturdier, higher quality plants. This, together with the fact that CO2 enriched plants make a more efficient use of water may impact favorably on the plant’s ability to overcome transplanting stress (Sasek et al., 1985). The relative increase in net assimilation rate due to an increase in CO2 concentration from 200-1000 ppm was almost as great at the lower as at the higher light level studied. The light compensation point is lowered by increased CO2 concentration (Mortensen, 1987). Fierro et al. (1994) demonstrated interactive effects of CO2 and light enrichment on tomato and pepper transplants. If either were applied 3 weeks before transplanting tomatoes and peppers, they increased accumulation of dry matter in shoots by 50%. Fierro et al. (1994) results suggested that it is more important to achieve optimal light conditions first and then make use of CO2 enrichment.
CO2 plant uptake mechanism: Plants and other photosynthetic organisms convert carbon dioxide into energy rich organic molecules, which are the precursors of carbohydrates. These molecules are either used by the organism directly or are passed into the food chain to be utilized by non-photosynthetic organisms (Paradise and Cyr, 1995). The complete photosynthetic process may be summarized by the following equation:
nCO2+nH2O→(CH2O)n+nO2
Carbon dioxide serves as substrate to photosynthetic carbon assimilation. There is concomitant decline in photo-respiratory activity and alteration in stomatal aperture. It was reported that C3 plants (wheat, rice, oilseeds, pulses, etc.) respond to elevated CO2 since elevated CO2 reduces oxygenase activity of RuBP carboxylase oxygenase enzyme in plants. The C4 plants (sorghum, maize, sugarcane, etc.) show little or no photosynthetic response to elevated CO2 because C4 pathway is not competitively inhibited by O2 and is completely CO2 saturated. However, there is no consensus on the quantitative effects of increased CO2 in plant processes and growth due to differences in response at different stages of growth, species of crops and because of growth limiting environmental factors (Uprety et al., 2000a).
The key enzyme for CO2 fixation is rubisco. Its activity depends on the ratio of the O2 and CO2 concentration in the atmosphere. The major effect of CO2 enrichment is the shift in balance between the carboxylation and oxygenation activity of rubisco, this effect is important at low and high light levels. Kimball (1983) stated that, on average, yields of crops should increase by 33% with a doubling of CO2 concentration in the earth’s atmosphere. Although these estimates have been developed for plants over their complete life cycles, enhanced growth and dry matter accumulation are correlated with higher net photosynthetic rates in young vegetative tissues under CO2 enrichment as well.
Studies on the response of rice cultivars Basmati-1 and Pusa-677 to the elevated CO2 showed increase in growth, photosynthesis and seed yield (Uprety et al., 2000b). The increased photosynthesis and greater accumulation of sugar contributed significantly to the accelerated development of leaves and tillers in both cultivars. The response of rice cultivars to the elevated CO2 with reference to their stomatal characters showed a marked increase in the stomatal resistance, stomatal index, size of guard cells, stroma and epidermal cells (Uprety et al., 2002).
Parameters affecting CO2 plant uptake: Ecosystem responses to CO2 enrichment and climate change are expressed at different levels: biochemical and stomatal processes at the leaf level, growth processes at the plant level and water budget and carbon-nitrogen cycling at the ecosystem level. Predicted responses of net primary production and N mineralization to CO2 enrichment and climate change are, therefore, complex. Responses to individual climate factors were often modified by responses to other factors, through interaction among processes at the same and different levels of organization (Coughenour and Chen, 1997). The final outcome of ecosystem responses to CO2 enrichment and climate change is dependent on the relative importance of these different processes. The relative enhancement of net primary production by CO2 enrichment was greater at high temperature in the C3 grass species and greater under ambient temperature conditions in the C4 grass. Most previous studies revealed that a CO2-doubling generally caused greater increases in growth at higher temperatures (Cure, 1985). Although, Sionit et al. (1981) reported that the CO2 response of okra had stimulatory effects on growth at lower temperature and Coleman and Bazzaz (1992) reported that the effects of CO2 enrichment on final biomass of a C4 annual forb was positive at low temperature and negative at high temperature. Chen et al. (1994) found that the relative enhancement of CO2 enrichment on an aquatic plant was strongly temperature dependent and that the maximum relative effects of CO2 were reached at a low temperature. These different responses suggest that the effects of elevated CO2 and temperature may interact in a seasonal environment and that this interaction is species specific (Hunt et al., 1991).
The responses of N mineralization to elevated CO2 were negative or near neutral except for the C3 grass species system at high precipitation. Reduced N mineralization under elevated CO2 may be ascribed to reduced litter quality (Strain and Bazzaz, 1983). However, N mineralization in the P. smithii system was increased by doubling CO2 under wet conditions. Increased N mineralization under elevated CO2 was also observed in an open top hamper study of Populus granditenta (Zak et al., 1993). They found up to five-fold increases in N mineralization. They suggested that elevated CO2 would increase below ground carbon inputs, which would increase microbial biomass and their activity and thus decomposition and mineralization rate. Decomposition and mineralization could be greater under elevated CO2 in spite of decreased residue quality, because decomposition and mineralization, being donor-controlled processes, tend to increase with net primary production (Hunt et al., 1991; Coughenour and Chen, 1997). Interactions among biochemical, ecophysiological and ecosystem processes appear to be significant. To reasonably estimate ecological effects of CO2 enrichment and climate change, a model must couple biochemical, ecophysiological and ecosystem processes. Ecosystem models that ignore physiological and ecophysiological responses are incapable of predicting responses to elevated CO2 and climate change (Coughenour and Chen, 1997). On the other hand, ecophysiological models that do not couple soil water and nitrogen cycling would be not able to represent the feedbacks of biogeochemical processes such as N mineralization. In natural ecosystems, many plant species coexist competitively. There are hundreds of plant species representing C3, C4 and CAM photosynthetic pathways in Colorado shortgrass steppe. The CO2 enrichment and climate change differently affect many plant and ecosystem processes. It has been hypothesized that different responses of C3 and C4 species to CO2 enrichment and climate change could change the competitive balance between these species in a future CO2 enriched atmosphere (Arp et al., 1993).
Trees treated with both nitrogen fertilizer and elevated CO2 showed a three-fold increase in growth over controls. The photosynthetic rates of plant species within a grassland community also showed little positive response to CO2 enrichment (Lee et al., 2001) observed after at least three years of fumigation (Oren et al., 2001). The stimulation of carbon sequestration in the vegetation and litter of forest trees was paralleled in grassland where CO2 was enriched by 240 ppm over a six-year period (Niklaus et al., 2001). The elevated CO2 increased the rate of photosynthesis, promoted greater foliage, more number of siliqua and increased root growth. The carbon need to satisfy the demand for these newly generated sinks was met from new photosynthates produced during the high CO2 exposure. Plants accumulated increased metabolic reserves in terms of sugars and starch to significantly enhance metabolic flexibility to respond to adverse moisture stress (Uprety et al. 1995. J.Crop Sci. & Agron.[Germany] 175, 231-237.; Uprety et al. 1995. "Climate variability of South Asia & its impact on agriculture. Narosa Publ. House, London/N.Delhi. P 262-278.).(Uprety et al., 1995). The main constraint to growth occurs when the plant is unable to photosynthesize at the potential rate, governed by the amount of incoming solar radiation. This will normally occur when one of the substrates for photosynthesis (carbon, water, nitrogen) is limiting (Agren, 1985).
Plant uptake of CO2 in landfill: The landfill industry makes use of a potentially valuable, environmentally beneficial and sustainable utilization of restored landfill sites. The biomass produced at restored landfill sites from Short Rotation Coppice (SRC) trees, such as willow and poplar, may have a number of uses: as a fuel for electricity generation plants; for the production of charcoal; as a soil amendment for clay caps; or simply as a carbon sink for atmospheric CO2 (Cannell et al., 1987).
Present municipal solid waste landfills generate biogas that is flared on site to destroy noxious contaminants and water is extracted from leachate to be drained away. However, biogas could alternatively be a cheap fuel for winter heating and could provide horticultural greenhouses with abundant carbon dioxide to boost plant growth all year long (Jaffrin et al., 2003).
In the research of Chan et al. (1997), two landfill sites in Hong Kong (Shuen Wan and Junk Bay StageI) were chosen as the field sites and three sites (Yuen Chau Kok, Tai Po and Lok Wo Sha) were chosen as reference sites. The vegetation survey classified the plants under woody plants, herbs and grasses groups. Seventeen species of vascular plants were found at the reference sites; whereas 25 and 20 species, respectively, were found at the two-landfill sites. The gross average coverage of plants at the reference sites (57.8%) was lower than that at the two-landfill sites (70.3, 61.4%). Tree cover on the landfill sites was dominated by two species: Acacia confusa and Leucaena leucocephala, which seemed suited for growth and were abundant on landfill sites. Therefore, completed landfill sites can be good habitats, which support a variety of plants and animals, provided that the plants can phytoremediate landfill gas and leachate contamination (Chan et al., 1997).
PHYSICAL BIOGAS EXTRACTION
Biogas collection systems: In landfills, gas is collected using network of collection pipes and wells. The gas collection efficiency in landfills is between 40-90% (Augenstein and Pacey, 1991). Biogas is collected by means of some vertical and horizontal drain pipes and is then burned in flares or employed to produce heat and energy (EMCON Associates, 1980). The designs for gas abstraction systems include different types of well configurations such as vertical, horizontal, hybrid, etc (Leach, 1990). The landfill gas collection system consists of vertical extraction well, transport pipe network, blower for passive gas collection or pumping for active collection, monitoring equipment and flare stack (Park and Shin, 2001).
Landfill gas can be collected by either a passive or an active collection system. Passive gas collection systems (Fig. 1) use existing variations in landfill pressure and gas concentrations to vent landfill gas into the atmosphere or a control system. The collection wells are typically constructed of perforated or slotted plastic and are installed vertically throughout the landfill to depths ranging from 50-90% of the waste thickness. Vertical wells are typically installed after the landfill or a portion of a landfill, has been closed. A passive collection system may also include horizontal wells located below the ground surface to serve as conduits for gas movement within the landfill (ATSDR., 2001).
Active gas collection (Fig. 2) is considered a good means of landfill gas collection (EPA., 1991). An active collection system composed of extraction wells connected to header pipe to a pump that delivers gas for energy recovery (Wheless and Wiltsee, 2001).
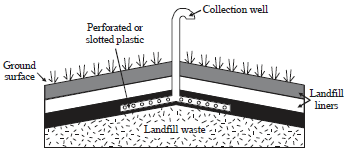 | |
| Fig. 1: | Passive gas collection system (ATSDR., 2001) |
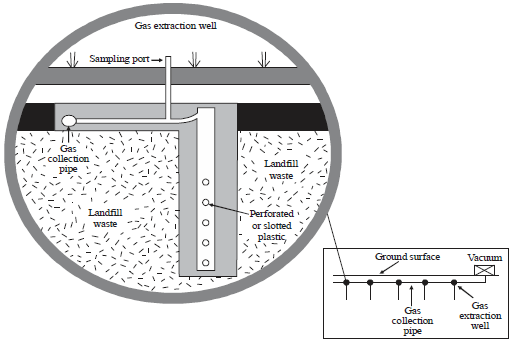 | |
| Fig. 2: | Active gas collection system (ATSDR., 2001) |
An active system consists of extraction well, header pipe, vacuum, condensate removal and burner (Bagchi, 1994, 2004). However, wells in the active system should have valves to regulate gas flow and to serve as a sampling port. Sampling allows the system operator to measure gas generation, composition and pressure (ATSDR., 2001).
The Environmental Protection Agency (EPA), state and local regulators use assumed gas collection efficiencies to calculate landfill emissions in regulating and setting policies for landfills. These assumed efficiencies are usually around 75%. A thorough review of gas system operators’ experiences and of researcher’s efficiency determination methods and their results is required to better assess actual collection efficiencies. As stated by Sanitation Districts of Los Angeles County in 2015.
Biogas control methods: There are two common methods of conducting a control to gas migration: using very low permeability materials to resist gas flow and using highly permeable materials to allow the gas to vent to the surface. Among combination of those methods the following could be summarized:
| • | Trench filled with granular backfill acts as passive system to collect gas from boundaries |
| • | Trench backfilled with gravel and an impermeable membrane installed along the back wall of the trench to trap the gas to be more effective interceptor |
| • | A gravel trench with vertical perforated pipe to collect gas within trench passive collection system |
| • | A gravel trench with horizontal and vertical perforated pipes to collect gas within the trench (McBean et al., 1995) |
Current methods of forming a gas resistant barrier usually involve the excavation of a trench and backfilling with either a low permeable material such as bentonite or the inclusion of a gas resistant membrane. Vent trenches are normally constructed using trenches backfilled with either gravel or geocomposite venting media to promote gas flow to the surface. An alternative method is to provide a series of discrete vent wells at regular spacing or using vertical permeable curtain. These methods allow the gas to exhaust directly to atmosphere without any dilution in the system (Wilson and Shuttleworth, 2002). The Vent System provide preferential pathway (route of least resistance) to atmosphere as shown in Fig. 3 (Permavoid Ltd., 2015).
Biogas collection layers: There are other processes of gas collection such as collection layers. Tire shreds have been used as landfill gas collection material at municipal solid waste landfills (Augustein et al., 1997). GeoSyntec Consultants (1998a) for the California Integrated Waste Management Board provided a summary of recommended procedures for use of tire shreds as landfill gas collection material at municipal solid waste landfills, including horizontal landfill gas collection layers, horizontal trenches and vertical boreholes. Tire shreds have a compressibility that is several orders of magnitude greater than materials typically used for landfill gas collection such as gravel, sand, or crushed rock (GeoSyntec Consultants, 1998b). Consideration should be given to using a properly selected geotextile as a separation between the tire shreds and soil materials. The geotextile would prevent soil from migrating into the relatively large voids between the tire shreds (GeoSyntec Consultants, 1998a).
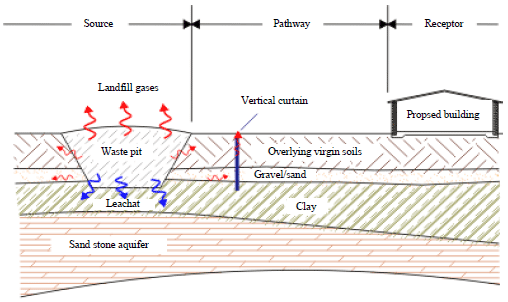 | |
| Fig. 3: | Vertical permeable venting curtain for gas control |
According to intrinsic permeability values, it is found that grain size fractions of 0.1-2, 0.6-1.2 (sand) and 0.1-56 mm (natural soil), are inappropriate for the application as gas collection layer materials, as they demonstrated lower values than a sample of solid waste (Huber and Wohnlich, 1999). As the mentioned grain size fractions of sand and natural soil belong to materials, which are commonly used for constructing the foundation layer of landfill cover systems (Fox, 1996), these layers cannot serve at the same time as gas collection layers. The coarse materials (grain size fraction of 16-32 mm of gravel, crushed lime stone, crushed granite and crushed basalt) were proved to be the most suitable for constructing gas collection layers, since it indicates high intrinsic permeability (Huber and Wohnlich, 1999).
Biogas collection covers: There are other methods of gas collection such as collection covers, which are used other than application in landfill, but could give good analogy of incorporated processes and/or materials to apply in landfill.
McGrath and Mason (2004) in their application to assess biogas production from anaerobic waste stabilization pond treating farm dairy wastewater, they used floating gas collection cover was constructed of fiber-reinforced polypropylene geomembrane fabric (J.P.S. Elastomerics, Westfield, NC, USA), supported on a 2 m by 2 m framework fabricated from 25 mm diameter and foam filled 110 mm diameter polyvinylchloride pipe and fittings.
Manure lagoons can be covered by floating polyethylene liners on the liquid surface allowing for the cost-effective construction of anaerobic digesters which use microorganisms to ferment and process waste producing methane gas. The polyethylene cover prevents vector migration out of the lagoon preventing odors, vapors and gasses from entering the atmosphere. They also prevent rainwater dilution. The most desirable effect of the polyethylene cover is to create an inflatable methane gas reservoir (Kriofske, 1998).
The XR-5 cover by ETP Inc, AL USA, is a proven material for biogas collection. It is strong, tough and it is highly resistant to biogas. Its low gas permeability makes it ideal for the collection of biogas and for use as a floating cover material. Poly-log-floats are used to buoy the cover and to form pass-ways for the collection of the biogas. Rainwater drainage is provided by the use of thru-cover drains. The cover’s unique design allows it to act as a gas storage container at times of peak production (ETP Inc., 2015).
A large employer in upper State Maine, USA, expanded their food processing capability. As part of that expansion, they needed to build an anaerobic digester that would retain heat in the cold climate of Maine and be able to collect the biogas produced. Lemna, MN, USA used LemTec™ Modular Insulated Cover System a turnkey to biogas design (Lemna Technologies Inc., 2015).
Geomembrane Technologies Inc. GTI, NB, Canada produces gas collection covers that are floating cover systems used to collect gases from wastewater treatment lagoons, sludge ponds, aeration systems, flow equalization tanks and pretreatment tanks. In the case of anaerobic digestors, GTI cover systems are used to capture biogas. The GTI cover system includes reduction of process heat loss/gain, elimination of water evaporation and prevention of sunlight penetration. Features can include insulation, automatic rainwater removal, baffles, sample ports and hatches. These durable, UV protected covers are strong enough to safely support foot traffic, light vehicles and snow loads (GTI., 2015).
The Canadian french-fry manufacturer, needed to replace an old (competitor’s) insulated gas collection cover that was prone to failure due to migrating solids (grease bergs) that ripped the gas collection membrane. The design of the old cover made it next to impossible to remove the grease bergs and to repair the damaged cover. The insulated gas collection cover by Lemna replaced the old one. The new gas collection cover has the ability to remove sections of the cover to dispel solids when required (Lemna Technologies Inc., 2015).
A dairy in Pennsylvania had a wastewater tank that was generating odors. Lemna Technologies proposed the installation of LemTec™ Gas Collection Cover System. This system provides effective odor control by completely covering the water surface with a single layer membrane. Channels are created beneath the cover for gas to flow for collection. Channels above the cover provide for rainwater removal (Lemna Technologies Inc., 2015).
Scotford and Williams (2001) investigated the effectiveness of a floating plastic cover to reduce ammonia emissions from slurry lagoon. They measured ammonia emissions from both an uncovered lagoon and covered lagoon. The ammonia emissions were measured using hoods. Each hood was fitted with an inlet pipe and outlet pipe. Air was forced into the hood using a fan that gave a superficial air velocity across the measured surface. The ammonia flux was measured into and out of the hoods. The floating plastic cover prevented nearly 100% of these emissions.
CONCLUSION
Biogas in landfill is intercepted by different means of biological and physical processes. The biological processes are natural activities such as bacterial methane oxidation and plant uptake for carbon dioxide at topsoil layer. Methane oxidation is affected by different environmental factors such as: temperature, water content, nutrients, substrate and oxygen concentrations. Plant uptake for carbon dioxide is affected by some parameters such as: CO2 concentration, nitrogen concentration, water content and temperature. The physical engineered processes are represented by various physical biogas extraction systems. Landfill gas can be collected by either a passive or an active collection system. Passive gas collection systems depend on variations of gas pressures and concentrations. An active collection system is an extraction systems depend on pumping that delivers gas for energy use.
REFERENCES
- Abichou, T., K. Mathieu, L. Yuan, J. Chanton and G. Hater, 2009. Effects of compost biocovers on gas flow and methane oxidation in a landfill cover. Waste Manage., 29: 1595-1601.
CrossRefDirect Link - Adams, R.S. and R. Ellis, 1960. Some physical and chemical changes in the soil brought about by saturation with natural gas. Soil Sci. Soc. Am. J., 24: 41-44.
CrossRefDirect Link - Anthony, C., 1986. Bacterial oxidation of methane and methanol. Adv. Microb. Physiol., 27: 113-210.
PubMedDirect Link - Arp, W.J., B.G. Drake, W.T. Pockman, P.S. Curtis and D.F. Whigham, 1993. Interactions between C3 and C4 salt marsh plant species during four years of exposure to elevated atmospheric CO2. Adv. Vegetation Sci., 14: 133-143.
CrossRefDirect Link - Baath, E., 1989. Effects of heavy metals in soil on microbial processes and populations: A review. Water Air Soil Pollut., 47: 335-379.
CrossRefDirect Link - Babich, H., G. Stotzky and H.L. Ehrlich, 1980. Environmental factors that influence the toxicity of heavy metal and gaseous pollutants to microorganisms. CRC Crit. Rev. Microbiol., 8: 99-145.
CrossRefDirect Link - Baggs, E.M. and H. Blum, 2004. CH4 oxidation and emissions of CH4 and N2O from Lolium perenne swards under elevated atmospheric CO2. Soil Biol. Biochem., 36: 713-723.
CrossRefDirect Link - Baille, M., R. Romero-Aranda and A. Baille, 1996. Gas-exchange responses of rose plants to CO2 enrichment and light. J. Hortic. Sci. Biotechnol., 71: 945-956.
Direct Link - Barta, T.M. and R.S. Hanson, 1993. Genetics of methane and methanol oxidation in gram-negative methylotrophic bacteria. Antonie Van Leeuwenhoek, 64: 109-120.
CrossRefDirect Link - Bender, M. and R. Conrad, 1992. Kinetics of CH4 oxidation in oxic soils exposed to ambient air or high CH4 mixing ratios. FEMS Microbiol. Lett., 101: 261-269.
CrossRefDirect Link - Bender, M. and R. Conrad, 1994. Microbial oxidation of methane, ammonium and carbon monoxide and turnover of nitrous oxide and nitric oxide in soils. Biogeochemistry, 27: 97-112.
CrossRefDirect Link - Boeckx, P. and O. van Cleemput, 1996. Methane oxidation in a neutral landfill cover soil: Influence of moisture content, temperature and nitrogen-turnover. J. Environ. Qual., 25: 178-183.
CrossRefDirect Link - Boeckx, P., O. van Cleemput and I. Villaralvo, 1996. Methane emission from a landfill and the methane oxidising capacity of its covering soil. Soil Biol. Biochem., 28: 1397-1405.
CrossRefDirect Link - Bogner, J.E., K.A. Spokas and E.A. Burton, 1997. Kinetics of methane oxidation in a landfill cover soil: Temporal variations, a whole-landfill oxidation experiment and modeling of net CH4 emissions. Environ. Sci. Technol., 31: 2504-2514.
CrossRefDirect Link - Bogner, J., K. Spokas, E. Burton, R. Sweeney and V. Corona, 1995. Landfills as atmospheric methane sources and sinks. Chemosphere, 31: 4119-4130.
CrossRefDirect Link - Bonan, G.B., D. Pollard and S.L. Thompson, 1992. Effects of boreal forest vegetation on global climate. Nature, 359: 716-718.
CrossRefDirect Link - Borjesson, G. and B.H. Svensson, 1997. Seasonal and diurnal methane emissions from a landfill and their regulation by methane oxidation. Waste Manage. Res., 15: 33-54.
CrossRefDirect Link - Bowden, R.D., K.M. Newkirk and G.M. Rullo, 1998. Carbon dioxide and methane fluxes by a forest soil under laboratory-controlled moisture and temperature conditions. Soil Biol. Biochem., 30: 1591-1597.
CrossRefDirect Link - Burrows, K.J., A. Cornish, D. Scott and I.J. Higgins, 1984. Substrate specificities of the soluble and particulate methane mono-oxygenases of methylosinus trichosporium OB3b. J. Gen. Microbiol., 130: 3327-3333.
CrossRefDirect Link - Cannell, M.G.R., R. Milne, L.J. Sheppard and M.H. Unsworth, 1987. Radiation interception and productivity of willow. J. Applied Ecol., 24: 261-278.
CrossRefDirect Link - Castro, M.S., J.M. Melillo, P.A. Steudler and J.W. Chapman, 1994. Soil moisture as a predictor of methane uptake by temperate forest soils. Can. J. For. Res., 24: 1805-1810.
CrossRefDirect Link - Castro, M.S., P.A. Steudler, J.M. Melillo, J.D. Aber and R.D. Bowden, 1995. Factors controlling atmospheric methane consumption by temperate forest soils. Global Biogeochem. Cycles, 9: 1-10.
CrossRefDirect Link - Chalvatzaki, E. and M. Lazaridis, 2009. Assessment of air pollutant emissions from the Akrotiri landfill site (Chania, Greece). Waste Manage. Res., 28: 778-788.
CrossRefDirect Link - Chanakya, H.N., I. Sharma and T.V. Ramachandra, 2009. Micro-scale anaerobic digestion of point source components of organic fraction of municipal solid waste. Waste Manage., 29: 1306-1312.
CrossRefDirect Link - Chan, G.Y.S., L.M. Chu and M.H. Wong, 2002. Effects of leachate recirculation on biogas production from landfill co-disposal of municipal solid waste, sewage sludge and marine sediment. Environ. Pollut., 118: 393-399.
CrossRefDirect Link - Chan, G.Y.S., L.M. Chu and M.H. Wong, 1997. Influence of landfill factors on plants and soil fauna-an ecological perspective. Environ. Pollut., 97: 39-44.
CrossRefDirect Link - Chanton, J., T. Abichou, C. Langford, K. Spokas and G. Hater et al., 2011. Observations on the methane oxidation capacity of landfill soils. Waste Manage., 31: 914-925.
CrossRefDirect Link - Chen, D.X., M.B. Coughenour, D. Eberts and J.S. Thullen, 1994. Interactive effects of CO2 enrichment and temperature on the growth of dioecious Hydrilla verticillata. Environ. Exp. Bot., 34: 345-353.
CrossRefDirect Link - Christophersen, M., L. Linderod, P.E. Jensen and P. Kjeldsen, 2000. Methane oxidation at low temperatures in soil exposed to landfill gas. J. Environ. Qual., 29: 1989-1997.
CrossRefDirect Link - Chugh, S., W. Clarke, P. Pullammanappallil and V. Rudolph, 1998. Effect of recirculated leachate volume on MSW degradation. Waste Manage. Res., 16: 564-573.
Direct Link - Coleman, J.S. and F.A. Bazzaz, 1992. The effects of CO2 and temperature on growth and resource use of co-occurring C3 and C4 annuals. Ecology, 73: 1244-1259.
CrossRefDirect Link - Coughenour, M.B. and D.X. Chen, 1997. Assessment of grassland ecosystem responses to atmospheric change using linked plant-soil process models. Ecol. Applic., 7: 802-827.
CrossRefDirect Link - Cureton, P.M., P.H. Groenevelt and R.A. McBride, 1991. Landfill leachate recirculation: Effects on vegetation vigor and clay surface cover infiltration. J. Environ. Qual., 20: 17-24.
CrossRefDirect Link - Czepiel, P.M., B. Mosher, P.M. Crill and R.C. Harris, 1996. Quantifying the effect of oxidation on landfill methane emissions. J. Geophys., 101: 16721-16729.
CrossRefDirect Link - DeLucia, E.H., J.G. Hamilton, S.L. Naidu, R.B. Thomas and J.A. Andrews et al., 1999. Net primary production of a forest ecosystem with experimental CO2 enrichment. Science, 284: 1177-1179.
CrossRefPubMedDirect Link - Diamadopoulos, E., 1994. Characterization and treatment of recirculation-stabilized leachate. Water Res., 28: 2439-2445.
CrossRefDirect Link - Dunfield, P., R. Knowles, R. Dumont and T.R. Moore, 1993. Methane production and consumption in temperate and subarctic peat soils: response to temperature and pH. Soil Biol. Biochem., 25: 321-326.
CrossRefDirect Link - Duxbury, J.M., 1994. The significance of agricultural sources of greenhouse gases. Fertilizer Res., 38: 151-163.
CrossRefDirect Link - El-Fadel, M., 1999. Leachate recirculation effects on settlement and biodegradation rates in MSW landfills. Environ. Technol., 20: 121-133.
CrossRefDirect Link - Enoch, H., 1990. Crop responses to aerial carbon dioxide-refereed paper. Acta Hortic., 268: 17-32.
CrossRefDirect Link - Fierro, A., A. Gosselin and N. Tremblay, 1994. Supplemental carbon dioxide and light improved tomato and pepper seedling growth and yield. HortScience, 29: 152-154.
Direct Link - Galloway, J.N., W.H. Schlesinger, H. Levy II, A. Michaels and J.L. Schnoor, 1995. Nitrogen fixation: Anthropogenic enhancement-environmental response. Global Biogeochem. Cycles, 9: 235-252.
CrossRefDirect Link - Green, J., S.D. Prior and H. Dalton, 1985. Copper ions as inhibitors of protein c of soluble methane monooxygenase of Methylococcus capsulatus (Bath). Eur. J. Biochem., 153: 137-144.
CrossRefDirect Link - Groffman, P.M., A.J. Gold and R.C. Simmons, 1992. Nitrate dynamics in riparian forests: Microbial studies. J. Environ. Qual., 21: 666-671.
CrossRefDirect Link - Hand, D.W., 1984. Crop responses to winter and summer CO2 enrichment. Acta Hortic., 162: 45-64.
CrossRefDirect Link - Hanson, R.S. and T.E. Hanson, 1996. Methanotrophic bacteria. Microbiol. Rev., 60: 439-471.
Direct Link - Houghton, J.T., L.G.M. Filho, B.A. Callandar, N. Harriss, A. Kattenberg and K. Maskell, 1996. Technical Summary. In: Climate Change 1995: The Science of Climate Change, Houghton, J.T., L.G.M. Filho, B.A. Callandar, N. Harriss, A. Kattenberg and K. Maskell (Eds.). Cambridge University Press, Cambridge, ISBN: 9780521564366, pp: 9-49.
- Hutsch, B.W., 1998. Methane oxidation in arable soil as inhibited by ammonium, nitrite and organic manure with respect to soil pH. Biol. Fertil. Soils, 28: 27-35.
CrossRefDirect Link - Hunt, H.W., M.J. Trlica, E.F. Redent, J.C. Moore and J.K. Detling et al., 1991. Simulation model for the effects of climate change on temperate grassland ecosystems. Ecol. Modell., 53: 205-246.
CrossRefDirect Link - Im, J., S. Moon, K. Nam, Y.J. Kim and J.Y. Kim, 2009. Estimation of mass transport parameters of gases for quantifying CH4 oxidation in landfill soil covers. Waste Manag., 29: 869-875.
CrossRefPubMedDirect Link - Ionescu, G., E.C. Rada, M. Ragazzi, C. Marculescu, A. Badea and T. Apostol, 2013. Integrated municipal solid waste scenario model using advanced pretreatment and waste to energy processes. Energy Convers. Manage., 76: 1083-1092.
CrossRefDirect Link - Isci, A. and G.N. Demirer, 2007. Biogas production potential from cotton wastes. Renewable Energy, 32: 750-757.
CrossRefDirect Link - Jaffrin, A., N. Bentounes, A.M. Joan and S. Makhlouf, 2003. Landfill biogas for heating greenhouses and providing carbon dioxide supplement for plant growth. Biosyst. Eng., 86: 113-123.
CrossRefDirect Link - Jahng, D. and T.K. Wood, 1996. Metal ions and chloramphenicol inhibition of soluble methane monooxygenase from Methylosinus trichosporium OB3b. Applied Microbiol. Biotechnol., 45: 744-749.
CrossRefDirect Link - Xin, J.Y., J.R. Cui, J.Z. Niu, S.F. Hua, C.G. Xia, S.B. Li and L.M. Zhu, 2004. Production of methanol from methane by methanotrophic bacteria. Biocatal. Biotransform., 22: 225-229.
CrossRefDirect Link - Jones, H.A. and D.B. Nedwell, 1993. Methane emission and methane oxidation in land-fill cover soil. FEMS Microbiol. Ecol., 11: 185-195.
CrossRefDirect Link - Kightley, D., D.B. Nedwell and M. Cooper, 1995. Capacity for methane oxidation in landfill cover soils measured in laboratory-scale soil microcosms. Applied Environ. Microbiol., 61: 592-601.
Direct Link - Kimball, B.A., 1983. Carbon dioxide and agricultural yield: An assemblage and analysis of 430 prior observations. Agron. J., 75: 779-788.
CrossRefDirect Link - King, M.A. and A.P.S. Adamsen, 1992. Effects of temperature on methane consumption in a forest soil and in pure cultures of the methanotroph Methylomonas rubra. Applied Environ. Microbiol., 58: 2758-2763.
Direct Link - King, G.M. and S. Schnell, 1994. Effect of increasing atmospheric methane concentration on ammonium inhibition of soil methane consumption. Nature, 370: 282-284.
CrossRefDirect Link - Kinman, R.N., D.L. Nutini, J.J. Walsh, W.G. Vogt, J. Stamm and J. Rickabaugh, 1987. Gas enhancement techniques in landfill simulators. Waste Manage. Res., 5: 13-25.
CrossRefDirect Link - Kravchenko, I., P. Boeckx, V. Galchenko and O. van Cleemput, 2002. Short- and medium-term effects of NH4+ on CH4 and N2O fluxes in arable soils with a different texture. Soil Biol. Biochem., 34: 669-678.
CrossRefDirect Link - Kumaraswamy, S., B. Ramakrishnan and N. Sethunathan, 2001. Methane production and oxidation in an anoxic rice soil as influenced by inorganic redox species. J. Environ. Qual., 30: 2195-2201.
PubMedDirect Link - LaDeau, S.L. and J.S. Clark, 2001. Rising CO2 levels and the fecundity of forest trees. Science, 292: 95-98.
CrossRefPubMedDirect Link - Le Mer, J. and P. Roger, 2001. Production, oxidation, emission and consumption of methane by soils: A review. Eur. J. Soil Biol., 37: 25-50.
Direct Link - Lee, T.D., M.G. Tjoelker, D.S. Ellsworth and P.B. Reich, 2001. Leaf gas exchange responses of 13 Prairie grassland species to elevated CO2 and increased nitrogen supply. New Phytol., 150: 405-418.
CrossRefDirect Link - Leita, L., M. De Nobili, G. Muhlbachova, C. Mondini, L. Marchiol and G. Zerbi, 1995. Bioavailability and effects of heavy metals on soil microbial biomass survival during laboratory incubation. Biol. Fertil. Soils, 19: 103-108.
CrossRefDirect Link - Lessard, R., P. Rochette, E. Topp, E. Pattey, R.L. Desjardins and G. Beaumont, 1994. Methane and carbon dioxide fluxes from poorly drained adjacent cultivated and forest sites. Can. J. Soil Sci., 74: 139-146.
CrossRefDirect Link - Liamsanguan, C. and S.H. Gheewala, 2008. LCA: A decision support tool for environmental assessment of MSW management systems. J. Environ. Manage., 87: 132-138.
CrossRefPubMedDirect Link - Lindhout, P. and G. Pet, 1990. Effects of CO2 enrichment on young plant growth of 96 genotypes of tomato (Lycopersicon esculentum). Euphytica, 51: 191-196.
Direct Link - Machadoa, S.L., M.F. Carvalho, J.P. Gourc, O.M. Vilar and J.C.F. do Nascimento, 2009. Methane generation in tropical landfills: Simplified methods and field results. Waste Manage., 29: 153-161.
CrossRefDirect Link - Matthews, E., 1994. Nitrogenous fertilizers: Global distribution of consumption and associated emissions of nitrous oxide and ammonia. Global Biogeochem. Cycles, 8: 411-439.
CrossRefDirect Link - Maurice, C., M. Ettala and A. Lagerkvist, 1999. Effects of leachate irrigation on landfill vegetation and subsequent methane emissions. Water Air Soil Pollut., 113: 203-216.
CrossRefDirect Link - McGrath, R.J. and I.G. Mason, 2004. An observational method for the assessment of biogas production from an anaerobic waste stabilisation pond treating farm dairy wastewater. Biosyst. Eng., 87: 471-478.
CrossRefDirect Link - Mishra, S.R., K. Bharati, N. Sethunathan and T.K. Adhya, 1999. Effects of heavy metals on methane production in tropical rice soils. Ecotoxicol. Environ. Saf., 44: 129-136.
CrossRefPubMedDirect Link - Mohanty, S.R., K. Bharati, N. Deepa, V.R. Rao and T.K. Adhya, 2000. Influence of heavy metals on methane oxidation in tropical rice soils. Ecotoxicol. Environ. Saf., 47: 277-284.
CrossRefDirect Link - Moreira, J.M.L., E.R. da Silva and G. Candiani, 2015. Approximate estimation of landfill emissions considering methane oxidation. Open Waste Manage. J., 8: 12-20.
Direct Link - Mortensen, L.M., 1987. Review: CO2 enrichment in greenhouses. Crop responses. Sci. Hortic., 33: 1-25.
CrossRefDirect Link - Warith, M.A., W. Zekry and N. Gawri, 1999. Effect of leachate recirculation on municipal solid waste biodegradation. Water Qual. Res. J. Can., 34: 267-280.
Direct Link - Murrell, J.C., I.R. McDonald and B. Gilbert, 2000. Regulation of expression of methane monooxygenases by copper ions. Trends Microbiol., 8: 221-225.
CrossRefDirect Link - Mora-Naranjo, N., J.A. Meima, A. Haarstrick and D.C. Hempel, 2004. Modelling and experimental investigation of environmental influences on the acetate and methane formation in solid waste. Waste Manage., 24: 763-773.
CrossRefDirect Link - Nguyen, H.H.T., A.K. Shiemke, S.J. Jacobs, B.J. Hales, M.E. Lidstrom and S.I. Chan, 1994. The nature of the copper ions in the membranes containing the particulate methane Monooxygenase from Methylococcus capsulatus (bath). J. Biol. Chem., 269: 14995-15005.
Direct Link - Nielsen, A.K., K. Gerdes and J.C. Murrell, 1997. Copper-dependent reciprocal transcriptional regulation of methane monooxygenase genes in Methylococcus capsulatus and Methylosinus trichosporium. Mol. Microbiol., 25: 399-409.
CrossRefDirect Link - Niklaus, PA., M. Wohlfender, R. Siegwolf and C. Korner, 2001. Effects of six years atmospheric CO2 enrichment on plant, soil and soil microbial C of a calcareous grassland. Plant Soil, 233: 189-202.
CrossRefDirect Link - Oren, R., D.S. Ellsworth, K.H. Johnsen, N. Phillips and B.E. Ewers et al., 2001. Soil fertility limits carbon sequestration by forest ecosystems in a CO2-enriched atmosphere. Nature, 411: 469-472.
CrossRefDirect Link - Park, J.W. and H.C. Shin, 2001. Surface emission of landfill gas from solid waste landfill. Atmos. Environ., 35: 3445-3451.
CrossRefDirect Link - Peterjohn, W.T., J.M. Melillo, P.A. Steudler, K.M. Newkirk, F.P. Bowles and J.D. Aber, 1994. Responses of trace gas fluxes and n availability to experimentally elevated soil temperatures. Ecolog. Applic., 4: 617-625.
Direct Link - Phelps, P.A., S.K. Agarwal, G.E. Speitel Jr. and G. Georgiou, 1992. Methylosinus trichosporium OB3b mutants having constitutive expression of soluble methane monooxygenase in the presence of high levels of copper. Applied Environ. Microbiol., 58: 3701-3708.
PubMedDirect Link - Prather, M., R. Derwent, D. Ehalt, P. Fraser, E. Sanhueza and X. Zhau, 1996. Other Trace Gases and Atmospheric Chemistry. In: Climate Change 1995: The Science of Climate Change, Houghton, J.T., L.G.M. Filho, B.A. Callandar, N. Harriss, A. Kattenberg and K. Maskell (Eds.)., Cambridge University Press, Cambridge, Uk, pp: 77-126.
- Prieme, A., and S. Christensen, 1997. Seasonal and spatial variation of methane oxidation in a Danish spruce forest. Soil Biol. Biochem., 29: 1165-1172.
CrossRefDirect Link - Rada, E.C., I.A. Istrate and M. Ragazzi, 2009. Trends in the management of residual municipal solid waste. Environ. Technol., 30: 651-661.
CrossRefDirect Link - Rada, E.C., M. Ragazzi and V. Torretta, 2013. Laboratory-scale anaerobic sequencing batch reactor for treatment of stillage from fruit distillation. Water Sci. Technol., 67: 1068-1075.
CrossRefPubMedDirect Link - Radoglou, K.M., P. Aphalo and P.G. Jarvis, 1992. Response of photosynthesis, stomatal conductance and water use efficiency to elevated CO2 and nutrient supply in acclimated seedlings of Phaseolus vulgaris L. Ann. Bot., 70: 257-264.
Direct Link - Reinhart, D.R., 1996. Full-scale experiences with leachate recirculating landfills: Case studies. Waste Manage. Res., 14: 347-365.
CrossRefDirect Link - Reinhart, D.R. and A.B. Al-Yousfi, 1996. The impact of leachate recirculation on municipal solid waste landfill operating characteristics. Waste Manage. Res., 14: 337-346.
CrossRefDirect Link - Sasek, T.W., E.H. DeLucia and B.R. Strain, 1985. Reversibility of photosynthetic inhibition in cotton after long-term exposure to elevated CO2 concentrations. Plant Physiol., 78: 619-622.
Direct Link - Scheutz, C. and P. Kjeldsen, 2004. Environmental factors influencing attenuation of methane and hydrochlorofluorocarbons in landfill cover soils. J. Environ. Qual., 33: 72-79.
CrossRefPubMedDirect Link - Scheutz, C., J. Bogner, J.P. Chanton, D. Blake, M. Morcet, C. Aran and P. Kjeldsen, 2008. Atmospheric emissions and attenuation of non-methane organic compounds in cover soils at a French landfill. Waste Manage., 28: 1892-1908.
CrossRefDirect Link - Scheutz, C., P. Kjeldsen, J.E. Bogner, A. De Visscher and J. Gebert et al., 2009. Microbial methane oxidation processes and technologies for mitigation of landfill gas emissions. Waste Manage. Res., 27: 409-455.
CrossRefDirect Link - Scotford, I.M. and A.G. Williams, 2001. Practicalities, costs and effectiveness of a floating plastic cover to reduce ammonia emissions from a pig slurry lagoon. J. Agric. Eng. Res., 80: 273-281.
Direct Link - Sellers, P.J., F.G. Hall, H. Margolis, B. Kelly and D. Baldocchi et al., 1995. The boreal ecosystem-atmosphere study (BOREAS): An overview and early results from the 1994 field year. Bull. Am. Meteorol. Soc., 76: 1549-1577.
CrossRefDirect Link - Sionit, N., B.R. Strain and H.A. Beckford, 1981. Environmental controls on the growth and yield of Okra. I. Effects of temperature and CO2 enrichment at cool temperatures. Crop Sci., 21: 855-888.
CrossRefDirect Link - Sitaula, B.K., L.R. Bakken and G. Abrahamsen, 1995. CH4 uptake by temperate forest soil: Effect of N input and soil acidification. Soil Biol. Biochem., 27: 871-880.
CrossRefDirect Link - Smith, D.S., T.E. Huxman, S.F. Zitzer, T.N. Charlet and D.C. Housman et al., 2000. Elevated CO2 increases productivity and invasive species success in an arid ecosystem. Nature, 408: 79-82.
CrossRefDirect Link - Sommerfeld, R.A., A.R. Mosier and R.C. Musselman, 1993. CO2, CH4 and N2O flux through a Wyoming snowpack and implications for global budgets. Nature, 361: 140-142.
CrossRefDirect Link - Spokas, K.A. and J.E. Bogner, 2011. Limits and dynamics of methane oxidation in landfill cover soils. Waste Manage., 31: 823-832.
CrossRefDirect Link - Stein, V.B. and J.P.A. Hettiaratchi, 2001. Methane oxidation in three alberta soils: Influence of soil parameters and methane flux rates. Environ. Technol., 22: 101-111.
CrossRefDirect Link - Steinkamp, R., K. Butterbach-Bahl and H. Papen, 2001. Methane oxidation by soils of an N limited and N fertilized spruce forest in the black forest, Germany. Soil Biol. Biochem., 33: 145-153.
CrossRefDirect Link - Steudler, P.A., R.D. Bowden, J.M. Melillo and J.D. Aber, 1989. Influence of nitrogen fertilization on methane uptake in temperate forest soils. Nature, 341: 314-316.
CrossRefDirect Link - Sulisti, I.A. Watson-Craik and E. Senior, 1996. Studies on the co-disposal of o-cresol with municipal refuse. J. Chem. Technol. Biotechnol., 65: 72-80.
CrossRefDirect Link - Sundh, I., P. Borga, M. Nilsson and B.H. Svensson, 1995. Estimation of cell numbers of methanotrophic bacteria in boreal peatlands based on analysis of specific phospholipid fatty acids. FEMS Microbiol. Ecol., 18: 103-112.
CrossRefDirect Link - Sundh, I., C. Mikkela, M. Nilsson and B.H. Svensson, 1995. Potential aerobic methane oxidation in a Sphagnum-dominated peatland-controlling factors and relation to methane emission. Soil Biol. Biochem., 27: 829-837.
CrossRefDirect Link - Takeguchi, M. and I. Okura, 2000. Role of iron and copper in particulate methane monooxygenase of Methylosinus trichosporium OB3b. Catal. Surv. Jpn., 4: 51-63.
CrossRefDirect Link - Thomas, J.F., C.D. Raper, C.E. Anderson and R.J. Downs, 1975. Growth of young tobacco plants as affected by carbon dioxide and nutrient variables. Agron. J., 67: 685-689.
CrossRefDirect Link - Tlustos, P., T.W. Willison, J.C. Baker, D.V. Murphy, D. Pavlikova, K.W.T. Goulding and D.S. Powlson, 1998. Short-term effects of nitrogen on methane oxidation in soils. Biol. Fertil. Soils, 28: 64-70.
CrossRefDirect Link - Topp, E. and E. Pattey, 1997. Soils as sources and sinks for atmospheric methane. Can. J. Soil Sci., 77: 167-177.
CrossRefDirect Link - Tremblay, N. and A. Gosselin, 1998. Effect of carbon dioxide enrichment and light. HortTechnology, 8: 524-528.
Direct Link - Tremblay, N., S. Yelle and A. Gosselin, 1987. Effects of CO2 enrichment, nitrogen and phosphorus fertilization on growth and yield of celery transplants. HortScience, 22: 875-876.
Direct Link - Trulli, E., V. Torretta, M. Raboni and S. Masi, 2013. Incineration of pre-treated Municipal Solid Waste (MSW) for energy co-generation in a non-densely populated area. Sustainability, 5: 5333-5346.
CrossRefDirect Link - Tuskan, G.A. and M.E. Walsh, 2001. Short-rotation woody crop systems, atmospheric carbon dioxide and carbon management: A U.S. case study. For. Chronicle, 77: 259-264.
CrossRefDirect Link - Uprety, D.C., N. Dwivedi and R. Mohan, 1998. Characterization of CO2 responsiveness in a Brassica oxycamp interspecific hybrid. J. Agron. Crop Sci., 180: 7-13.
CrossRefDirect Link - Uprety, D.C., N. Dwivedi, V. Jain and R. Mohan, 2002. Effect of elevated carbon dioxide concentration on the stomatal parameters of rice cultivars. Photosynthetica, 40: 315-319.
CrossRefDirect Link - Uprety, D.C., S. Kumari, N. Dwivedi and R. Mohan, 2000. Effect of elevated CO2 on the growth and yield of rice. Indian J. Plant Physiol., 5: 105-107.
Direct Link - Uprety, D.C., R.S. Mishra and Y.P. Abrol, 1995. Effect of elevated CO2 on the photosynthesis, growth and water relation of Brassica species under moisture stress. J. Agron. Crop Sci., 175: 231-237.
CrossRefDirect Link - Urban, L., 1994. Effect of high-pressure mist and daytime continuous CO2 enrichment on leaf diffusive conductance, CO2 fixation and production of rosa hybrida plants grown on rockwool. Acta Horticulturae, 361: 317-324.
CrossRefDirect Link - Vaccari, M., V. Torretta and C. Collivignarelli, 2012. Effect of improving environmental sustainability in developing countries by upgrading solid waste management techniques: A case study. Sustainability, 4: 2852-2861.
CrossRefDirect Link - Van Der Nat, F., J. de Brouwer, J. Middelburg and H.J. Laanbroek, 1997. Spatial distribution and inhibition by ammonium of methane oxidation in intertidal freshwater marshes. Applied Environ. Microbiol., 63: 4734-4740.
Direct Link - Wand, S.J.S., G.Y.F. Midgley, M.H. Jones and P.S. Curtis, 1999. Responses of wild C4 and C3 grass (poaceae) species to elevated atmospheric CO2 concentration: A meta-analytic test of current theories and perceptions. Global Change Biol., 5: 723-741.
CrossRefDirect Link - Whalen, S.C., W.S. Reeburgh and K.A. Sandbeck, 1990. Rapid methane oxidation in a landfill cover soil. Applied Environ. Microbiol., 56: 3405-3411.
Direct Link - Wong, S.C., 1979. Elevated atmospheric partial pressure of CO2 and plant growth. I. Interactions of nitrogen nutrition and photosynthetic capacity in C3 and C4 plants. Oecologia, 44: 68-74.
CrossRefDirect Link - Woodrow, L., B. Grodzinski and A. Liptay, 1987. The effects of CO2 enrichment and ethephon application on the production of tomato transplants. Acta Horticulturae, 201: 133-140.
CrossRefDirect Link - Zak, D.R., K.S. Pregitzer, P.S. Curtis, J.A. Teen, R. Fogel and D.L. Randlett, 1993. Elevated atmospheric CO2 and feedback between carbon and nitrogen cycles. Plant Soil, 151: 105-117.
CrossRefDirect Link - Zhang, C., H. Su, J. Baeyens and T. Tan, 2014. Reviewing the anaerobic digestion of food waste for biogas production. Renew. Sustain. Energy Rev., 38: 383-392.
CrossRefDirect Link - Zhang, H., P. He and L. Shao, 2008. Methane emissions from MSW landfill with sandy soil covers under leachate recirculation and subsurface irrigation. Atmos. Environ., 42: 5579-5588.
CrossRefDirect Link