
A. H. Fouly
Department Plant Production and Protection,College of Agriculture and Veterinary Medicine, Qassim University,51452 Buraydah, P.O. Box 6622, Saudi Arabia
S. M. Al-Rehiayani
Department Plant Production and Protection,College of Agriculture and Veterinary Medicine, Qassim University,51452 Buraydah, P.O. Box 6622, Saudi Arabia
Journal of Entomology
Year: 2011 | Volume: 8 | Issue: 2 | Page No.: 139-151







ABSTRACT
Incidence of predaceous mite species inhabiting different habitats were recorded in Saudi Arabia for the first time in 2008-2009. Predatory mites were mounted, classified and identified in Zoology Lab at the College of Agriculture and Veterinary Medicine, Qassim University. Thirty one predaceous mites were collected from samples of plant shoot and root systems of cultivated plants, debris and organic manure in Al-Qassim region. These mite species belong to three suborders, 16 families and 29 genera. Acarid mites represented 6.46%, Actendid mites represented 25.8%, while Gamasid mites, which was the most abundant group of predaceous mites, represented 67.74% of the whole collected species during the study. Occurrence, site of collection, habitat and remarks of predatory mites are listed. The surveyed mite species are considered new records for the Saudi fauna. Illustration and description of the two new laelapid species, Hypoaspis zaheri; H. dactylifera are included.
PDF Abstract XML References Citation
Received: March 14, 2010;
Accepted: May 04, 2010;
Published: July 22, 2010
How to cite this article
A. H. Fouly and S. M. Al-Rehiayani, 2011. Predaceous Mites in Al-Qassim Region, Saudi Arabia, with Description of Two New Laelapid Species (Acari: Gamasida: Laelapidae). Journal of Entomology, 8: 139-151.
DOI: 10.3923/je.2011.139.151
URL: https://scialert.net/abstract/?doi=je.2011.139.151
DOI: 10.3923/je.2011.139.151
URL: https://scialert.net/abstract/?doi=je.2011.139.151
INTRODUCTION
The acari exhibit various associations with other organisms. Since beneficial areal and soil mites especially predatory species must be conserved in the field to promote a more stabilized pest and natural balance (Fouly, 1997; Walter and Campbell, 2003; Heckmann et al., 2007). Although, little is known about the habits of many micro arthropods such as mites that form together the belowground food web. Gamasida (Mesostigmata) fauna is extremely rich and a considerable number of this group is an efficient biological control agents for mite and insect pests as well as those living on the shoot system (leaves and buds) and known as aerial predators (Moraes and McMurtry, 1985; Rasmy et al., 1987; Abbasipour et al., 2006; Wekesa et al., 2007; Ukabi et al., 2009). Certain species of predatory mites are used in biological control programs in many countries (Fouly et al., 1997; Takano-Lee and Hoddle, 2002; Cuthbertson et al., 2007). Among this group of mites, family Laelapidae Berlese may be considered the most important group of aggressive soil predators especially those in the subfamily Hypoaspidinea, including genus Geolaelaps and Stratiolaelaps. Stratiolaelaps species are often referred to the trashbin genus Hypoaspis Canestrini (Walter and Campbell, 2003). Recently, some species of this genus are commercially used in biological control against agriculture pests in glasshouses especially in the Northern Hemisphere (Walter and Oliver, 1990; Walter and Campbell, 2003). These species were previously found to prey on different food sources such as larvae of housefly Musca domestica L., vinegar fly, Drosophila melnogaster Morgan (Sherif and Afifi, 1980) mushroom sciarid fly Lycoriella solani Winnertz (Enkegaard et al., 1997); immature stages of the western flower thrips Frankliniella occidentalis (Pergande) (Berndt et al., 2004a, b) springtail Folsomia fimetaria L. (Baatrup et al., 2005) egg masses of root-knot nematode, Meloidogyne javanica Treub, citrus nematode, Tylenchulus semipenetrans (Cobb) and acarid mite Caloglyphus rodriguezi Samsinak (Al-Rehiayani and Fouly, 2006) and springtail Folsomia candida Willem and F. fimetaria L. and acrid mite Caloglyphus michaeli Oudemans and an oligochaeta Enchytraeus crypticus (Salmane and Brumelis, 2008).
The present study is conducted in Al-Qassim region, middle of Saudi Arabia for the first time, to determine the incidence of predaceous mite species living either on the shoot system of growing plants or leaf litter, organic manure and soil surrounding root system. Description and illustration of the two new laelapid mite species Hypoaspis zaheri n.sp. and H. dactylifera n.sp. are also included.
MATERIALS AND METHODS
Mites collection: Predaceous mites associated with different stages of insect, mite and nematode species infesting shoot or root system, living in debris and organic manure were surveyed during 2008-2009 at certain localities in Qassim region, Saudi Arabia. Samples of dead and alive mite individuals were collected and transferred into Zoology Laboratory at the College of Agriculture and Veterinary Medicine, Qassim University. Aerial mites were directly examined and collected by using a stereoscopic binocular. Mites inhabit debris, organic manure or root system were extracted by using the modified Tullgren's funnels for 24 h. After that, mite individuals were preserved in 70% alcohol until examination.
Taxonomical studies: The collected mites were placed in Nesbitt's solution as a clearing agent for a period not less than one day according to their sclerortization. After that, mite individuals were singly mounted on glass slides in Hoyer's fluid, covered with a glass cover and gently heated for few seconds to stretch mite bodies (Krantz, 1978). Date, site of collection and names of associated organisms were placed on each slide for further survey and classification studies. Identification of the collected predaceous mites were based on the terminology of Chant (1963), Chant and Baker (1965), Lindquist and Evanz (1965), Evanz and Till (1979), Summers and Price (1970), Hughes (1976), Bregetova et al. (1977), Chant and Yoshida (1978), Krantz (1978), Zaher et al. (1986), Denmark and Muma (1989), Karg (1993), Fouly et al. (1994) and Smiley et al. (1996). Taxonomical and identification studies of collected mites were carried out by using an Olympus Camera DP25 attached to an Olympus Microscope BX51, which both are connected to a HP Computer. Illustration of mite species was conducted by the aid of drawing eye piece attached to a phase contrast microscope. In most specimens, length of body is the distance between seta j1 and J5; width of that is the distance between bases of the seta s4 on propodosma and all measurements are given in micrometers (μm).
Statistical analysis: Data of the average numbers of mite individuals in each family and inhabiting different habitats and found at each locality were statistically analyzed by ANOVA test in completely randomized design. To compare the means of mite populations by Turkey's (p>0.05) test, after √X+0.5 transformation.
RESULTS
Based on literature and updated information, there is no previous information about taxonomy and identification of predaceous mite species inhabiting different habitats neither in Al-Qassim region nor in Saudi Arabia. It was noticed that aerial predators were observed feeding on phytophagous mites, scale insects and whiteflies. Soil predators were collected together with egg masses of nematodes, eggs and larvae of certain insect species, acarid mites and spring tails. The present study indicated the incidence of 31 predaceous mite species, of which 21 species in 19 families belong to the suborder Gamasida and presented 67.74%. Actinedid mites were eight species in seven families and presented 25.8%, while the Acarid mites were only two species in one family representing 6.46% of the total amount of collected mite species.
It is well known that some of the collected mite species such as uropodid, acarid and tarsonemid mites are considered fungivorous or of miscellaneous feeding habits. However, the collected uropodid species Leiodinychus sp. was frequently noticed feeding on small dipterous larvae in soil samples. Tarsonemus sp. was observed feeding on scale insects, while both acarid species Mycetoglyphus sp. and Tyrophagus puterscentiae were noticed feeding on egg masses of plant parasitic nematodes. The present data showed that suborder Gamasida were represented by the families, Pytoseiidae (four species in four genera); Ascidae (four species in four genera); Macrochelidae (two species in one genus); Parasitidae (one species); Laelapidae (four species in three genera); Uropodidae (one species); Rhodacaridae (two species in two genera); Ologamasidae (two species in two genera). From these results it can be noticed that mites belonging to families Phytoseiidae, Laelapidae and Ascidae were the most abundant species where four species were collected in each family, while only two species were found in family Macrochelidae, Ologamasidae and Rhodacaridae. A single species were found belonging to family Parasitidae and Uropodidae. On the other hand, Suborder Actinedida was represented by only one species in families Cheyletidae, Bdellidae, Cunaxidae, Neophylobiidae, Stigmaeidae, Raphignathidae and Tarsonemidae. Suborder Acaridida was represented by only one family, Acaridae (two species in two genera) as shown in Table 1. The results also showed that mites Agistemus exsertus and Mycetoglyphus sp., Leiodinychus sp. and Sessiluncus sp. were the most abundant species where their population considerably exceeds 15 individuals and recorded in two localities. Gamasiphis puleculus and Parasitus sp. were also abundant but they were only recorded in one location. The rest of the collected mites were found in moderate or few numbers. In general, there were no significant differences between number of mite individuals in each locality, while the average of total numbers of mite species were significantly differed between different sites of survey (LSD 1.49, p = 0.05).
Among the areal predatory mites, Metaseiulus (Typhlodromus) pyri was found associated with the two spotted spider mite Tetranychus urticae Koch. Euseius scutalis was found with T. urticae, citrus brown mite Eutetranychus orientalis Klein and Whitefly Bemicia tabaci (Genn.). Typhlodromips (Amblyseius) swirsiki was observed feeding on citrus brown mite, while Phytoseius plumifer was found associated with scale insects and two spotted spider mite. Concerning Amblyseius cydnodactylon, it was rarely found associated with immature stages of insects living in the surface leaf litter surrounding date palm trees.
On the other hand, the stigmaemid mite A. exsertus, which can be considered the most abundant predatory mite species throughout the present study, was frequently observed feeding on all stages of the two spotted spider mite T. urticae infesting plants in three locations.
| Table 1: | Taxonomical categories, incidence and habitats of predaceous mite species collected from different sites in Al-Qassim region in 2008-2009 |
 | |
| ++++: More than 15 mite individuals, +++: 10-15 mite individuals, ++: 5-9 mite individuals and +: 1-4 mite individuals | |
Taxonomy and identification studies showed that Hypoaspis zaheri; H. dactylifera (Laelapidae) are considered new species. Both species were recorded in moderate numbers in debris and leaf litter surrounding date palm trees in Onayzah and Melida districts.
Family laelapidae: This family was represented by four species in three genera, Hypoaspis dactylifera n.sp., H. zaheri n.sp., A. casalis casalis and C. simplex.
Hypoaspis canestrini: Type species: Gamasus krameri Canestrini
Diagnosis: Chelicerae typically chelate-dentate with the movable digit bidentate in female. Pilus dentilis normally short, setiform. Female with anal shield free. Metasternal setae not on sterna shield. Genu IV with nine setae where pl2 absent. Female with genital shield flask- or tounge-shape, if with opithogenital shield, never with whip-like setae, marginal setae smooth.
Hypoaspis zaheri n.sp.
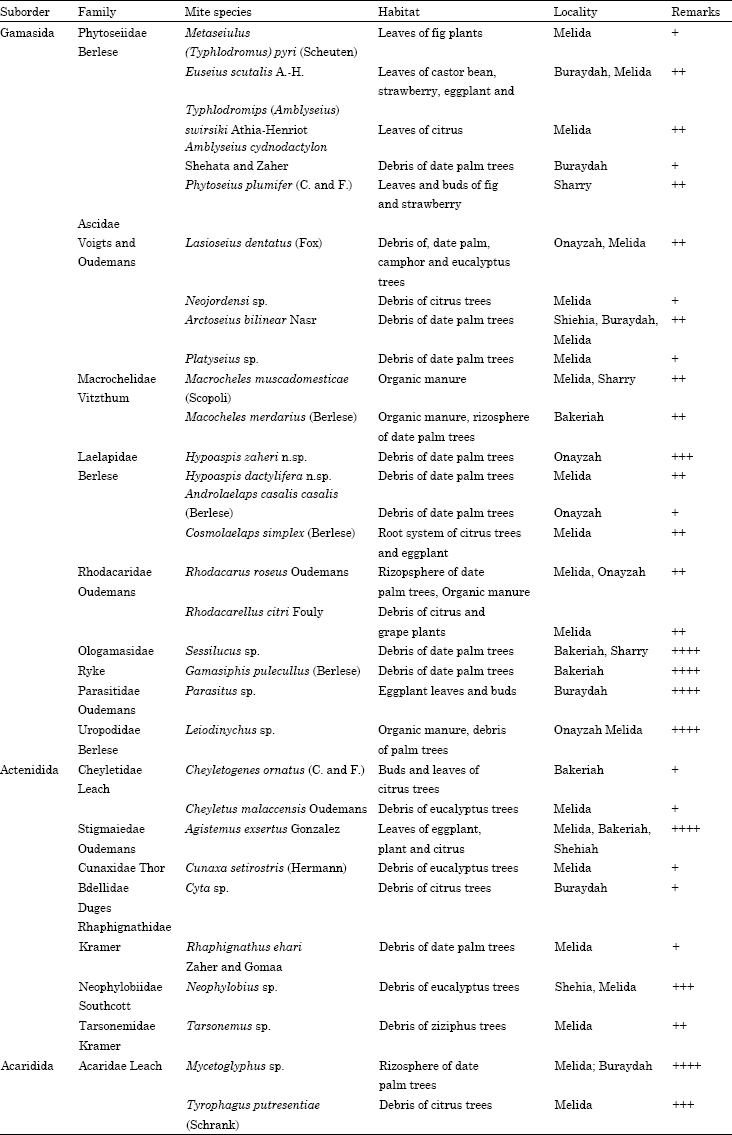
Female (Holotype) (Fig. 1a-g): This species is considered a new and stands nearly to H. petrovae Sherief and Afifi, 1980 but it can be easily distinguished by having only 35 pairs of simple setae, of which 22 pairs on podonotum and 13 pairs on opithonotum. setae S4 whip-like. female body ovate, light brown in color when alive, 563.12 μ long and 333.67 μ wide at level of setae j6 (Fig. 1a).
Dorsum: Dorsal shield entire, smooth, with shoulders, almost covers idiosoma. Podonotum covered with scattered sub-triangle and semi-quadrate striation, while opithonotum covered with a faintly network striation. Lateral setae on r, s, S and Z series longer than other dorsal setae; setae S4 whip-like and the longest. Eleven pairs of marginal setae arise on lateral membrane. Six pairs of pores locate on dorsal shield, of which five pairs circular and one crescent shape between setae J3 and S4 (Fig. 1a).
Venter: Presternal area punctuate with faintly dotted striation. Sternal shield faintly reticulate, 93 μ long and 89.2 μ wide, extend distally to middle of coxae III, with three pairs of sub-equal lanceolate sterna setae (st1-st3). Sternum with a pair of anterior lyriform pores, while the posterior pair circular (Fig. 1b). Metasternal seta (st4) arises on the membrane at the level of Coxae III. Genital shield tongue-shaped, covered with faintly network striation and with a pair of genital simple seta.
 | |
| Fig. 1: | Hypoaspis zaheri n.sp., (a) female dorsum, (b) female venter, (c) tectu, (d) female chelicerae, (e) genu-tibia leg II, (f) (genu-tibia leg III) and (g) genu-tibia leg IV |
Anal shield sub-triangle or pear-shape, reticulated with on its anterior half and bearing two sub-equal para-anal and a longer post-anal setae. Soft opithgasteric membrane surrounding genital and anal shields with five pairs of simple setae, setae Jv5 the longest. Peritreme well developed and with peritremal plate extending anterodorsally to reach the level of coxae I as shown in Fig. 1b. Tritosternum with an elongate base and coarsely serrate laciniae.
Gnathosoma: Deutosternum with seven rows of denticles. Corniculi well sclerotized horn-like and reaching to anterior margin of palp femur. Anterior margin of gnathotectum with sub-triangle denticulate edge (Fig. 1c). Fixed cheliceral digit bidentate and with eight small median teeth, three of which sub apical in addition to a setiform pilus dentilis, while the movable digit bidentate (Fig. 1d).
Legs: Leg setation formula for genu II 2,3/1,2/1,2 of which one macrosetae, tibia II 2,2/1,2/1,2 (Fig. 1e); genu III 2,2/1,2/1,1; tibia III 2,1/1,2/1,1 (Fig. 1f); genu IV 2,2/1,3/1,1 of which one macrosetae and tibia IV 2,1/1,3/1,2 (Fig. 1g).
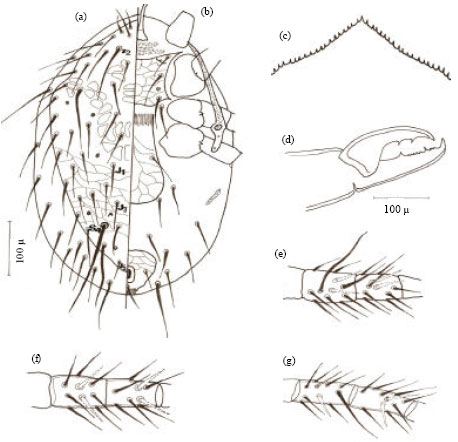
Male (allotype): Body smaller than female, 372.53 μ long and 199.16 μ wide, dorsal setation similar to that of female and dorsum similarly ornamented with reticula. Holoventral shield mostly covering ventral surface and sharply concaved from both sides anterior to the anus and bearing 10 pairs of sub equal stout setae (Fig. 2a).
 | |
| Fig. 2: | Hypoaspis zaheri n.sp., (a) male venter and (b) male chelicerae |
Fixed cheliceral digit with four small teeth between two big teeth. The movable digit with a single big tooth. Spermatodactyl long and about twice as long as chelicerae (Fig. 2b).
Remarks: Moderate numbers of mite individuals were found with different small insect and mite species living in leaf litter to a depth of 10cm surrounding date palm trees in Onayzah region on 20 May, 2008.
Specimens examined: Holotype female and allotype male were found in leaf litter samples in Onayzah. Twelve paratype females and males had the same information.
Etymology: This species is named after Dr. Mohamed A. Zaher, Professor of Acarology, Cairo University, Egypt, for his outstanding work and dedication to the Arabian Acarologists for more than 50 years.
Hypoaspis dactylifera n.sp.
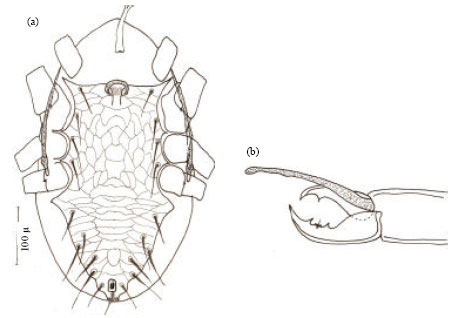
Female (Holotype) (Fig. 3a-g): This species is considered a new and stands nearly to H. aculeifer Canestrini and H. queenslandicus (womersley) but it can be easily distinguished by having dorsal shield with shoulders, an obvious sub-triangular posterior margin and with only 37 pairs of simple setae. Podonotal setae 23 pairs longer than opithonotal setae (14 pairs). Setae z1 longer than j1 and setae Z5 the longest on dorsal shield (Fig. 3a). Female body ovate, light brown in color when alive, body 456.13 μ long and 239.76 μ wide.
Dorsum: Dorsal shield smooth, interscutal membrane surrounding dorsal shield with nine pairs of simple setae. Dorsal shield with nine pairs of circular pores and a single pair of crescent-shape pores just behind setae z1 (Fig. 3a).
 | |
| Fig. 3: | Hypoaspis dactylifera n.sp., (a) female dorsum, (b) female venter, (c) tectum, (d) female chelicerae, (e) genu-tibia leg II, (f) genu-tibia leg III and (g) genu-tibia leg IV |
Venter: Presternal area obviously punctuate with faintly striation. Sternal shield longer than wide, 107.07 μ long and 86.62 μ wide, faintly reticulate, with three pairs of sub-equal sternal setae and two pairs of pores, the first one lyriform, while the second pair circular and locate between st2 and st3. A single pair of circular pores occur behind sternal shield. Metasternal setae st4 free on the membrane at the level of middle of coxae III (Fig. 3b). Genital shield tongue shaped, 143.90 μ long and 68.46 μ wide, faintly reticulate, obviously separated from anal shield and with one pair of genital setae. Anal shield pear shape, faintly reticulated only on its anterior half and with two para-anal and one post-anal seta. Primary metapodal plate narrow and smaller than secondary elongate metapodal plate. Soft opithogastric membrane with five pairs of simple sub-equal setae. Peritreme well developed and with peritremal plate extending anterodorsally to reach setae z1as shown in Fig. 3a-b. Tritosternum with obvious base and coarsely serrate laciniae (Fig. 3b).
Gnathosoma: Deutosternum with seven denticules. Corniculi horn-like. Tectum with rounded denticulate edge (Fig. 4c). Cheliceral fixed digit bi-dentate with seven small teeth, three of which sub apical as well as a setiform pilus dentilis. Movable digit bidentate (Fig. 3d).
Legs: Leg II thicker than leg IV, while leg I the thinnest. Ventral setae on legs stout. Setation formula for genu II 1,3/1,1/1,1; tibia II 2,3/0,3/1,1 (Fig. 3e); genu III 1,2/1,2/1,2; tibia III 1,2/1,1/1,2 (Fig. 3f); genu IV 2,2/1,3/0,1 and tibia IV 1,2/2,3/1,1 (Fig. 3g).
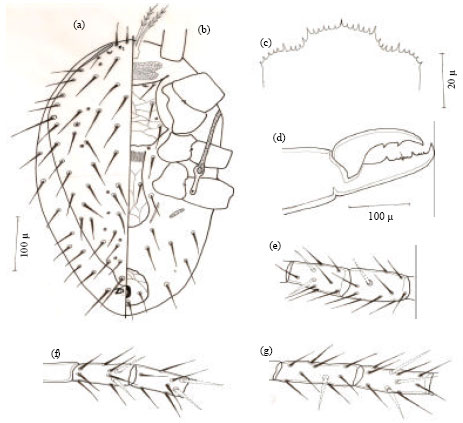
Male (Allotype): Smaller than female, dorsal shield measuring 398.56 μ long and 255.76 μ wide. Dorsal setae chaetotaxy similar to that of female. Holo-ventral shield mostly covering ventral surface, reticulate and bearing ten pairs of sub-equal simple setae and two pairs of circular pores (Fig. 4a).
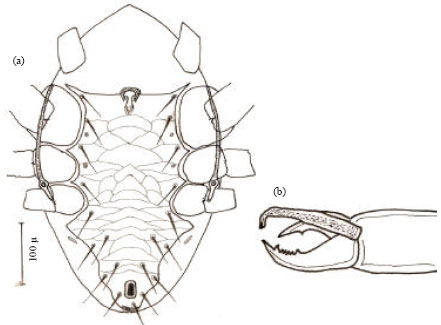 | |
| Fig. 4: | Hypoaspis dactylifera n.sp., (a) male venter and (b) male chelicerae |
Femur of leg II with a ventral spur, while femur, genu and tibia II with a spine like setae (plunt at tip). Cheliceral fixed digit with four small teeth between two bigger teeth. The movable digit with a single tooth and bearing an elongate L-shape spermatodactyl which hardly exceeding chelicerae in length (Fig. 4b).
Remarks: This species is named after date palm trees Pheonex dactylifera. Few mite individuals were found associated with different insect and mite species living in leaf litter under date palm trees in Melida. Specimens examined: Holotype female and allotype male were found in leaf litter samples in Melida district in November, 2008. Six paratype females and males had the same information.
DISCUSSION
Because we have no previous information about mite fauna especially predaceous mites neither in Al-Qassim region nor in Saudi Arabia, the present study is considered pioneer in this area. According to habitats and prey kinds, the collected predatory mite species could be classified into two major groups, (a) aerial predators that living on different plant parts above ground such as leaves, buds and branches and (b) soil predators that are collected from soil, debris and organic manure. In early study done in Egypt, Zaher et al. (1986) recorded some predatory mites species belonging to the families Cunaxidae, Raphignathidae and Cheyletidae. Zaher et al. (1986) mentioned that it is well known that most mites belonging to uropodid, tarsonemid and acarid mites are considered fungivorous or of miscellaneous feeding habits. However, in the present study it was found that the collected uropodid species Leiodinychus sp. was frequently noticed feeding on small dipterous larvae in soil samples. Tarsonemus sp. was observed feeding on scale insects, while both acarid species Mycetoglyphus sp. and Tyrophagus puterscentiae were noticed feeding on egg masses of plant parasitic nematodes. These observations agree with the findings of Al-Rehiayani and Fouly (2006) who successfully reared the aforementioned acarid mites on egg masses of root-knot nematode Meloidogyne javanica (Treub). Quilici et al. (1997) similarly surveyed the predatory mites inhabiting different habitats on the French Island of La Réunion (Indian Ocean) and found only twenty one species. They recorded ten phytoseiid, 1 tydeid, 2 bedellid, 2 cunaxid, 1 cheyeltid, 2 stigmaeid, 2 anystid and 1 erythraeid species.
The present data showed that mites belonging to families Phytoseiidae, Laelapidae and Ascidae were the most abundant species where four species were collected in each family, while we found only two species in families Macrochelidae, Ologamasidae, Rhodacaridae and Acaridae. A single species were found belonging to family Parasitidae, Uropodidae, Cheyletidae, Bdellidae, Cunaxidae, Neophylobiidae, Stigmaeidae, Raphignathidae, Tarsonemidae as shown in Fig. 1. Similar observations were obtained by El-Banhawy et al. (2006), who recorded 16 species of soil predatory mites in citrus orchards of the Nile Delta and Middle Egypt and they found that species of gamasid predators, especially those belonging to family Uropodidae, were the most common. In the present survey, we didn't find any predators in certain families such as Erytheridae and Eupalopsellidae. While Andrew et al. (2007) and Fadamiro et al. (2009) found some species in these two families in addition to species in Stigmaeidae, Anystidae, Bdellidae, Cheyletidae, Cunaxidae, Ascidae and Phytoseiidae. They added that population of Phytoseiidae (18 species) and Stigmaeidae (one species) were the most abundant.
The results obviously showed that A. exsertus and M. sp., L. sp. and S. sp., G. puleculus and P. sp., were the most abundant species. The aerial predatory mites, especially those in family Phytoseiidae, were found associated with tetranychid, tenuipalpid and eriophyid phytophagous mites as well as different stages of insects. These findings agree with those of Fouly et al. (1994), McMurtry and Croft (1997), Nomikou et al. (2002), Inbar and Gerling (2008) and Al-Shammery (2010), who mentioned that phytoseiid predators can feed on a wide range of preys and food substances. Moreover, most of these mites are commercially used as biological control agents against mite and insect pests in many countries (Chant and McMurtry, 2005a, b). Although, some of the collected mites have miscellaneous feeding habits such as acarid mites, it was noticed that M. qassimi and T. putrescentiae can feed on the gelatinous cover and egg masses of root-knot nematode. Al-Rehiayani and Fouly (2006) supported these findings where they successfully reared acarid mites on egg masses of nematodes.
On the other hand, Hypoaspis zaheri; H. dactylifera (Laelapidae) are considered new species. Similarly, Halliday (2005) found 56 species of predatory mites belonging to 14 families in different habitats in South Africa, where eight species in six families were described as new species, among those, H. calcarata, H. muellerae, Gamasiphoides muellerae, G. lootsi and G. rykei (Ologamasidae).
In conclusion, Predaceous mites are free living arthropods mainly inhabiting different habitats. Food preference and specialization are not clear enough so far. Field observations proved that the collected predatory mites have been seen feeding on mites, small insects and nematodes. However, the abundance and presence of predaceous mites might be related to growing plants, type of soil, organic matter, agrochemical applications and availability of prey as indicated in earlier studies (Al-Rehiayani and Fouly, 2005; El-Benhaway, 2002; El-Benhawy et al., 2006) .
ACKNOWLEDGMENT
This study was financially supported by the Deanship of Scientific Research at Qassim University, Saudi Arabia, Project No. SR-S-008-005 (2008-2009). The authors also highly appreciated the efforts of Mr. M.M. Belal and Mr. S. Al-Neghemshi, Zoology laboratory, College of Agriculture and Veterinary Medicine, Qassim University in collecting samples. We also thank Dr. M. A. Zaher, Professor of Acarology, Cairo University, Egypt for his valuable advice.
REFERENCES
- Abbasipour, H., A. Taghavi, K. Kamali and A. Sahragard, 2006. A new species of the genus tarsonemus canestrini et fanzago (Acari: Tarsonemidae) from tea gardens of Iran. J. Entomol., 3: 23-25.
CrossRefDirect Link - Al Rehiayani, S.M. and A.H. Fouly, 2005. Cosmolaelaps simplex (Berlese), a polyphagous predatory mite feeding on root-knot nematode Meloidogyne javanica and citrus nematode Tylenchulus semipenetrans. Pak. J. Biol. Sci., 8: 168-174.
CrossRefDirect Link - Al-Shammery, K.A., 2010. Different biological aspects of the predaceous mite Euseius scutalis (Athis-Henroit) and the effects due to feeding on three tetranychid mites in Saudi Arabia. Asian J. Biol. Sci., 3: 77-84.
Direct Link - Baatrup, E., M. Baley and J.A. Axelsen, 2005. Predation of the mite Hypoaspis aculeifer on the springtail Folsomia fimetaria and the influence of sex, size, starvation and poisoning. Entomol. Exp. Applic., 118: 61-70.
CrossRef - Berndt, O., R. Meyh�fer and H.M. Poehling, 2004. The edaphic phase in the ontogenesis of Frankliniella occidentalis and comparison of Hypoaspis miles and Hypoaspis aculeifer as predators of soil-dwelling thrips stages. Biological control, 30: 17-24.
CrossRef - Berndt, O., H.M. Poehling and R. Meyh�fer, 2004. Predation capacity of two predatory laelapid mites on soil-dwelling thrips stages. Entomol. Exp. Et Applicata, 112: 107-115.
CrossRef - Chant, D.A. and J.A. McMurtry, 2005. A review of the subfamily Amblyseiinae muma (Acari: Phytoseiidae): Part V. tribe Amblyseiini Wainstein: Subtribe Proprioseiopsina Chant and McMurtry. Int. J. Acarol., 31: 3-22.
CrossRef - Chant, D.A. and J.A. McMurtry, 2005. A review of the subfamily Amblyseiinae muma (Acari: Phytoseiidae): Part VII. The tribe Typhlodromipsini n. tribe. Int. J. Acarol., 31: 315-340.
CrossRef - El-Benhawy, S.M., 2002. Survey of predatory mites in the kingdom of Lesotho (Africa): Notes on altitudinal preference of predatory mites and description of a new species (Acari: Phytoseiidae). Int. J. Acarol., 28: 187-191.
CrossRef - El-Benhawy, S.M., A.K. Nasr and S.I. Afia, 2006. Survey of predaceous soil mites (Acari: Mesostigmata) in citrus orchards of the Nile Delta and Middle Egypt with notes on the abundance of the citrus parasitic nematode Tylenchulus semipenetrans (Tylenchida: Tylenchulidae). Int. J. of Tropical Insect Sci., 26: 64-69.
CrossRefDirect Link - Enkegaard, A., M.A. Sardar and H.F. Brodsgaard, 1997. The predatory mite Hypoaspis miles: Biological and demographic characteristics on two prey species, the mushroom sciarid fly, Lycoriella solani and the mould mite, Tyrophagus putrescentiae. Entomol. Exp. Et Applicata, 82: 135-146.
CrossRefDirect Link - Fadamiro, H.Y., Y. Xiao, M. Nesbitt and C.C. Childers, 2009. Diversity and seasonal abundance of predacious mites in Alabama Satsuma citrus. Ann. Entomol. Soc. Am., 102: 617-628.
CrossRef - Fouly, A.H., 1997. Effects of prey mites and pollen on the biology and life tables of Proprioseiopsis asetus (Chant) (Acari, Phytoseiidae). J. Applied Entomol., 121: 435-439.
CrossRefDirect Link - Fouly, A.H., M.M. Abou Setta and C.C. Childers, 1997. Redescription of Cosmolaelaps simplex berlese (Acari: Laelapidae) with notes on its male. Int. J. Acarol., 23: 33-37.
CrossRefDirect Link - Halliday, R.B., 2005. Predatory mites from crops and pastures in South Africa: Potential natural enemies of redlegged earth mite Halotydeus destructor (Acari: Penthaleidae). Zootaxa, 1079: 11-64.
Direct Link - Heckmann, L.H., A. Ruf, K.M. Nienstedt and P.H. Krogh, 2007. Reproductive performance of the generalist predator Hypoaspis aculeifer (Acari: Gamasida) when foraging on different invertebrate prey. Applied Soil Ecol., 36: 130-135.
CrossRef - Inbar, M. and D. Gerling, 2008. Plant-mediated interaction between whiteflies, herbivores and natural enemies. Ann. Rev. Entomol., 53: 431-448.
Direct Link - Moraes, G.J. and J.A. McMurtry, 1985. Comparison of T. evansi and T. urticae (Acari: Tetranychidae) as prey for eight species of phytoseiid mites. Entomophaga, 30: 393-397.
CrossRef - McMurtry, J.A. and B.A. Croft, 1997. Life styles of phytoseiid mites and their roles in biological control. Ann. Rev. Entomol., 42: 291-321.
CrossRef - Nomikou, M., A. Janssen, R. Schraag and M.W. Sabelis, 2002. Phytoseiid predators suppress populations of Bemicia tabaci on cucumber plants with alternative food. Exp. Applied Acarol., 27: 57-68.
CrossRef - Quilici, S., S. Kreiter, E. Ueckermann and D. Vincenot, 1997. Predatory mites (Acari) from various crops on Reunion. Int. J. Acarol., 23: 283-291.
CrossRef - Salmane, I. and G. Brumelis, 2008. The importance of the moss layer in sustaining biological diversity of Gamasina mites in coniferous forest soil. Pedobiologia, 52: 69-76.
CrossRef - Takano-Lee, M. and M. Hoddle, 2002. Predatory behaviors of Neoseiulus californicus and Galendromus helveolus (Acari: Phytoseiidae) attacking Oligonychus perseae (Acari: Tetranychidae). Exp. Applied Acarol., 26: 13-26.
Direct Link - Walter, D.E. and N.J.H. Campbell, 2003. Exotic vs endemic biocontrol agents: Would the real Stratiolaelaps miles (Berlese) (Acari: Laelapidae), please stand up? Biol. Control, 26: 253-269.
CrossRefDirect Link - Zaher, M.A., M.I. Mohamed and Abdel-Halim, 1986. Incidence of mites associated with stored seeds and food products in Upper Egypt. Exp. Applied Acarol., 2: 19-24.
CrossRef - Denmark, H.A. and M.H. Muma, 1989. A Revision of the Genus Amblyseius Berlese, 1914 (Acari: Phytoseiidae). Vol. 4, Florida Department of Agriculture and Consumer Services, USA., pp: 145-148.
Direct Link - Hughes, A.M., 1976. The Mites of Stored Food and Houses. 2nd Edn., H.M. Stationery Office, London, ISBN-10: 0112409091, Pages: 400.
Direct Link - Lindquist, E.E. and G.O. Evans, 1965. Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata). Mem. Entomol. Soc. Canada, 97: 5-66.
CrossRefDirect Link - Wekesa, V.W., G.J. Moraes, M. Knapp and I. Delalibera Jr., 2007. Interactions of two natural enemies of Tetranychus evansi, the fungal pathogen Neozygites floridana (Zygomycetes: Entomophthorales) and the predatory mite, Phytoseiulus longipes (Acari: Phytoseiidae). Biological Control, 41: 408-414.
CrossRefDirect Link - Cuthbertson, A.G.S. and Archie K. Murchie, 2007. A review of the predatory mite Anystis baccarum and its role in apple orchard pest management schemes in Northern Ireland. J. Entomol., 4: 275-278.
CrossRefDirect Link