
B.M. Md-Zain
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
S.J. Lee
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
M. Lakim
The Board of Trustees of Sabah Parks, Kinabalu Park, Kota Kinabalu, Sabah, Malaysia
A. Ampeng
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
M.C. Mahani
School of Environmental and Natural Resource Sciences, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Journal of Biological Sciences
Year: 2010 | Volume: 10 | Issue: 4 | Page No.: 348-354







ABSTRACT
This study was carried out to ascertain the molecular phylogenetic position of Tarsius bancanus among Malaysian primates based on the partial of Cytochrome b (cyt b) gene sequences. A total of five samples of Tarsius bancanus from Sabah, Malaysia, were used in this study. Several other Malaysian primates were also included in the analysis (Leaf monkeys (Presbytis and Trachypithecus), Macaques (Macaca), Siamang (Symphalangus) and Slow loris (Nycticebus). We also included DNA sequences of several prosimians (Galago, Cheirogaleus, Daubentonia, Indri, Avahi, Lemur and Lepilemur) from GenBank. In addition, one individual of orang utan (Pongo pygmaeus) and human (Homo sapiens) were used as outgroups to root the tree. All taxa were analysed using character method (Maximum Parsimony, MP) and distance method (Neighbor-Joining, NJ). From the 375 examined characters, 43.2% were constant characters while 4.8% characters were parsimony uninformative whereas 52.0% characters were parsimony informative. Tree topologies discriminated three major clades in which primitive primates, Old World Monkeys and Anthropoids belongs to their own monophyletic clades. Both MP and NJ trees showed that T. bancanus was placed in primitive primates group.
PDF Abstract XML References Citation
How to cite this article
B.M. Md-Zain, S.J. Lee, M. Lakim, A. Ampeng and M.C. Mahani, 2010. Phylogenetic Position of Tarsius bancanus Based on Partial Cytochrome b DNA Sequences. Journal of Biological Sciences, 10: 348-354.
DOI: 10.3923/jbs.2010.348.354
URL: https://scialert.net/abstract/?doi=jbs.2010.348.354
DOI: 10.3923/jbs.2010.348.354
URL: https://scialert.net/abstract/?doi=jbs.2010.348.354
INTRODUCTION
The Bornean tarsier, Tarsius bancanus is a nocturnal primate (Napier, 1970), which is classified under the family Tarsiidae (Brandon-Jones et al., 2004; Szalay and Delson, 1979). They can be found on the island of Borneo (Groves, 2001). Tarsiers have sometimes been called ‘living fossils’. The proportions of the limbs, which indicate their tree-hopping gait, are very similar to those of early primates of the Eocene period (Napier, 1970). Tarsier possesses problem in the primate systematics. Its evolutionary position is hard to be clarified. Mittermaier et al. (1999) stated that there are evidence suggesting tarsiers to be most closely related to the monkeys and apes than prosimians. For example, like anthropoid, tarsiers have a short muzzle rather than typical prosimians that externally have relatively long muzzles terminating in a naked, moist snout (Mafham and Mafham, 1992).
Recent developments in molecular techniques allow evolutionary biologists with an additional tool for making phylogenetics inferences, by comparing DNA sequences between homologous DNA segments using mitochondrial genes (Shahrom et al., 2005; Lim et al., 2010). These mitochondrial DNA (mtDNA) genes are widely used to infer phylogenetic relationships on primates (Masters et al., 2007; Roos et al., 2008). Cyt b gene of the mitochondrial is well-known as rapidly evolve gene and can shows variations within and between species and has been used in phylogeny and biogeography studies (Caine′ et al., 2006; Karanth et al., 2008).
Many molecular systematic researches have studied on primates focusing on Great Apes (Steiper, 2006; Gonder et al., 2006) and Old World Monkeys (Karanth et al., 2008; Osterholz et al., 2008). In addition, there are also many molecular studies on primitive primates particularly tarsiers (Raina et al., 2005; Uddin et al., 2008). Andrew et al. (1998) have looked on cyt b gene of T. bancanus and Murayama et al. (1998) worked on D4 dopamine receptor (D4DR) genes of prosimians (including tarsier) and tree shrew. In addition, Schmitz et al. (2002) had sequenced the entire mitochondrial genome of T. bancanus. Most of the molecular systematic studies on Malaysian primates focused only at the genus level, for instance the genus Presbytis, Trachypithecus and Nasalis (Ernie et al., 2005; Md-Zain et al., 2005, 2008). Other molecular studies from neighbouring country (Thailand) also incorporated many samples of cercopithecidae (Chaveerach et al., 2007; Tanee et al., 2009). Therefore, very limited inferences can be made on the molecular systematics of Malaysian primitive primates. Data obtained from this research is paramount important as to resolve difficulties in prosimian primate classifications. This study was carried out to determine phylogenetic position of T. bancanus from Sabah, Malaysia, compared to the other primates particularly the Malaysian primates using partial DNA sequences of cyt b gene.
MATERIALS AND METHODS
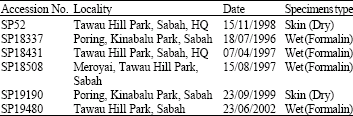
Taxa sampled: There were six T. bancanus tissue samples used in this study (Table 1). All the tarsier samples were obtained from Maklarin Lakim of Sabah Parks. Several institutions (Sabah Parks, Department of Wildlife and National Parks and Sarawak Forestry Department) provided the necessary facilities and assistance for tissue sampling collection of the other Malaysian primates since, 1998.
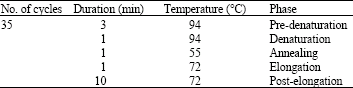
DNA extraction, amplification and sequencing: The cyt b gene was chosen to reflect sequence variation in mitochondrial DNA (mtDNA). Total DNA was extracted from tissue and skin using Gene All Tissue SV (Plus!) Mini Extraction Kit (Gene All). L14724 and H15149 (Table 2) were used as primers to amplify the cyt b gene (Irwin et al., 1991; Kocher et al., 1989). We managed to get the optimum condition of PCR components (MgCl2, Taq DNA polymerase, dNTP mix, PCR buffer, DNA template) in DNA amplification of cyt b gene (Table 3). Estimated fragment size of the cyt b gene for all amplified samples, observed after electrophoresis on 1.5% agarose gel is approximately 500 bp (Fig. 1).
| Table 1: | Tissue samples of Tarsius bancanus used in this study |
 | |
| Table 2: | Oligonucleotide primer pair and PCR conditions |
 | |
| Forward/Reverse Primer Sequences. L14724: 5'-CGAAGCTTGATATGAAAAACCATCGTTG-3' H15149:5'-AAACTGCAGCCCCTCCGAATGATATTTGTCCTCA-3‘ | |
The PCR products were purified using QIAquick gel purification kit protocol (QIAGEN) and sent to company (1st Base, Kuala Lumpur) for sequencing purposes.
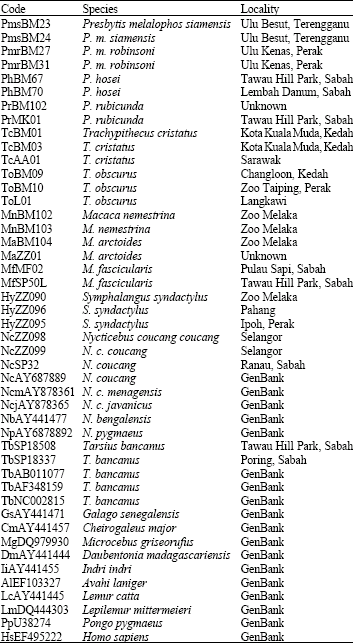
Sequence alignment and data analysis: Sequences were aligned using the ClustalW program, with minor adjustments made manually. We added several other Malaysian primate DNA sequences that are available in our laboratory database and other primate sequences from GenBank (Table 4). Pongo pygmaeus and Homo sapiens were used as outgroup to properly rooting the generated tree. Data matrix was analyzed by excluding 15 characters with the remaining 375 characters.
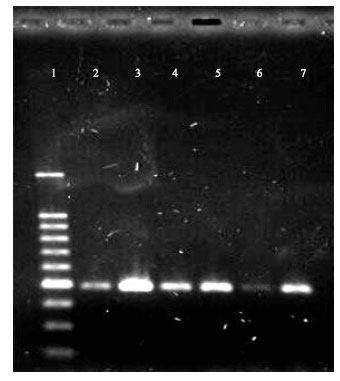 | |
| Fig. 1: | Amplified products of T. bancanus of mtDNA cyt b gene. Lanes: 1, 100 bp ladder; 2, T. bancanus (SP52); 3 T. bancanus (SP18337); 4, T. bancanus (SP18431); 5, T. bancanus (SP18508); 6, T. bancanus (SP19190); 7, T. bancanus (SP19480) |
| Table 3: | PCR components in DNA amplification of cyt b gene for tissue samples |
 | |
| *The quality depends on DNA concentration from DNA extraction | |
| Table 4: | Details of taxa used in the phylogenetic analysis |
 | |
Two primary methods were used to discern phylogenetic relationships, Maximum Parsimony (MP) and Neighbor Joining (NJ). The analysis were conducted using PAUP version 4.0. For unweighted MP, trees were obtained by heuristic searches treating all nucleotide substitutions as unordered. Data were also subjected to bootstrap analysis with 1000 replications. Tree topology constructed using the NJ method employing the Kimura 2 parameters distance option of PAUP. Homoplasy was quantified using the consistency index (CI) and the homoplasy index (HI).
RESULTS
All DNA from the six samples of T. bancanus were successfully amplified (Fig. 1). However, only two of them (SP18337 and SP18508) were successfully sequenced with the length of more than 400 bp.
| Table 5: | Summary of variations along the sequences across taxa |
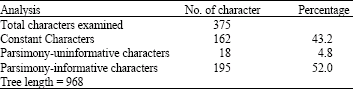 | |
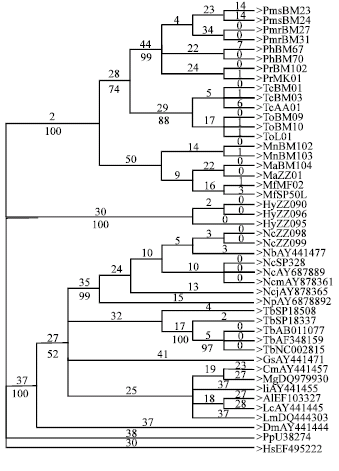 | |
| Fig. 2: | Maximum Parsimony tree of partial cyt b gene sequences obtained from heuristic searches. The bootstrap values are shown below the branches of the parsimony tree |
The other samples failed to produce any sequence that can be analysed. This problem was due to the degradation of samples where most of the samples have been stored for a long period and those samples are the formaldehyde-fixed tissue.
In phylogenetic tree obtained from the analysis of partial cyt b gene sequences, 15 characters were excluded because of missing data during data generated and the sequences do not represent variation. Thus, 375 remaining characters were examined with 43.2% constant characters. 4.8% characters are parsimony uninformative and 52.0% characters are parsimony informative (Table 5).
Figure 2 and 3 are useful in indicating the tree topologies obtained from Unweighted Maximum Parsimony and Neighbor-Joining analyses of the partial cyt b gene sequences.
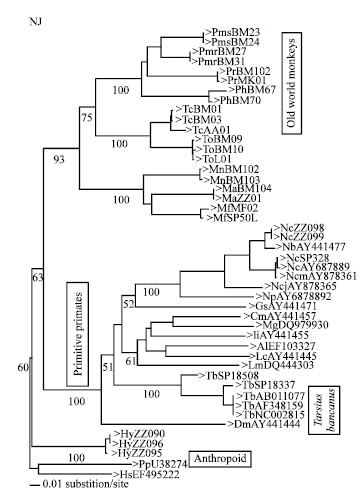 | |
| Fig. 3: | Neighbor-Joining phylogram with bootstrap values |
| Table 6: | Average percentages of genetic distance among anthropoids, old world monkeys and primitive primates using kimura 2 parameters |
 | |
Unweighted MP analysis produced a bootstrap tree (length = 968, CI = 0.3428, HI = 0.6572) with T. bancanus to form its own monophyletic clade with supported 100% of bootstrap value. NJ tree performed clearer topology, which also distinguish Tarsius in its own monophyletic clade supported by 100% bootstrap value. Both MP and NJ placed Tarsius within primitive primate clade and discriminated them from Old World Monkeys and Hylobatidae. Besides looking at tree topologies and bootstrap value, support for distinct monophyletic relationship among these three major clades can also be derived from Kimura 2 Parameters distance matrix. Table 6 summarized the average percentage of genetic distance among three major groups: anthropoids, Old World Monkeys and primitive primates (including Tarsius). Among these three groups, genetic distance between Old World Monkeys and primitive primates is the greatest, which is 37.199%, followed by anthropoids-primitive primates, 30.776%. The Old World Monkeys and anthropoids are the closest; their average percentage of genetic distance is 26.023%.
DISCUSSION
Tarsiers placed at a special position among the primates, in that they are generally intermediate between lemur and loris groups and also between monkeys and apes (Martin, 1990). In this study, both MP and NJ analysis (Fig. 2 and 3) indicated that T. bancanus is more closely related and group together with primitive primates (Strepsirrhini) rather than Old World Monkeys and anthropoid (Symphalangus). Results in this study support findings of many previous studies (Eizirik et al., 2001; Murphy et al., 2001; Jow et al., 2002; Schmitz et al., 2002; Hudelot et al., 2003) in which tarsiers have a close relationship with strepsirrhini. The results support Murphy et al. (2001), which grouped tarsiers with lemurs and separated them from New World Monkeys and great apes. While comparison of DNA sequences of the α A-crystalline gene (Jaworski, 1995) and γ-globin genes (Hayasaka et al., 2003) also indicated a difference between tarsiers and simians. In Schmitz et al. (2002) study, phylogenetic trees reconstruction based on amino acid and DNA sequences support a cluster of Tarsius and slow loris.
The position of tarsier depends on the molecular and morphological data sets (Matsui et al., 2009). Some previous data have been contradicted with our findings and even suggested tarsiers to be a close to anthropoidea (Schmitz et al., 2001; Poux and Douzery, 2004; Gibson et al., 2005) or as a basal group of primates (Arnason et al., 2002). At morphological level, Shoshani et al. (1996) stated that Tarsius is grouped with anthropoidea based on of 264 morphological characters on 18 primate taxa and four outgroups. Ross et al. (1998) also concluded that cranial evidence supports the tarsier-anthropoid hypothesis. Phylogenetic trees resulted from molecular analysis of the gene encoding Interphotoreceptor Retinoid Binding Protein (IRBP) and ε-globin sequences indicated that tarsier is grouped with Haplorhini (Stanhope et al., 1993). Moreover, a study of the existence pattern from short interspersed elements (SINEs), successfully showed a sister group relationship of T. bancanus with anthropoids to the exclusion of the Strepsirhini representatives (Schmitz et al., 2001). Mitochondrial cytochrome c oxidase subunit II reveals only a weak affinity of tarsiers to anthropoids (49.6% of bootstrap value) (Adkins and Honeycutt, 1994). In the study by Madsen et al. (2003), which utilised a new phylogenetic marker, apolipoprotein B (APOB) had showed a strong support (84-98% bootstrap value) in maximum likelihood for an association of tarsier and anthropoidea within primates.
Schmitz et al. (2002) stated that there is a general conflict exists at the molecular level between nuclear and mitochondrial DNA data. Phylogenetic analysis based on nuclear DNA sequences traditionally represent Tarsius as a sister group to anthropoids. In contrast, mitochondrial DNA data only marginally support this affiliation or even exclude Tarsius from primates. They suggested two possible scenarios causing this to be in conflict: a period of adaptive molecular evolution or a shift in the nucleotide composition of higher primate mitochondrial DNA through directional mutation pressure. Andrew et al. (1998) suggested that an episode of adaptive evolution might have taken place on the lineage leading to higher primates after Tarsius diverged. This is based on a higher relative rate of non-synonymous to synonymous substitutions. These evolutionary forces caused tarsiers to shift from their historical place in the mitochondrial DNA tree.
Results of this study indicate the phylogenetic problems concerning tarsiers. However, this study has increased our understanding of phylogenetic relationships among Malaysian primates particularly T. bancanus. Results showed that T. bancanus form its own monophyletic clades and clustered together with other primitive primates (strepsirrhini) towards Old World Monkeys and ape. Thus, T. bancanus belongs to primitive primates as supported by tree topologies, bootstrap values and genetic distance of cyt b DNA sequences. What now needs to be done is further work to define the molecular phylogenetic relationships of T. bancanus among other members of tarsiers as have been previously well described using morphological data (Groves, 1998; Merker and Groves, 2006). More population genetic studies should also incorporated other tarsier species in the Southeast Asian region as previously conducted by Merker et al. (2007, 2008) using microsatellite markers.
ACKNOWLEDGMENTS
We are deeply indebted to several institutions that provided necessary facilities and assistance including Universiti Kebangsaan Malaysia, Department of Wildlife and National Parks, Zoo Melaka, Zoo Taiping, Sarawak Forestry Department and Sabah Parks. We specifically thank Vun Vui Fui and Norlindawati Abd. Pateh for sharing their DNA sequences data. We are indebted to Ang Khai Chung for sharing his time during the study. We wish to thank Farhana Shukor and anonymous reviewers for their comments on the manuscript. This research was made possible under grants IRPA 0802020019 EA301, UKM-GUP-ASPL-07-04-146, UKM-KRIB-16/2008, UKM-OUP-PLW-14-59/2008 and UKM-OUP-TKP-20-97/2009.
REFERENCES
- Adkins, R.M. and R.L. Honeycutt, 1994. Evolution of the primate cytochrome c oxidase subunit II gene. J. Mol. Evol., 38: 215-231.
PubMed - Madsen, H.A, K.P. Koepfli, R.K. Wayne and M.S. Springer, 2003. A new phylogenetic marker, apolipoprotein B, provides compelling evidence for eutherian relationships. Mol. Phylogenetics Evol., 28: 225-240.
CrossRef - Andrew, T.D., L.S. Jermiin and S. Easteal, 1998. Accelerated evolution of cytochrome b in simian primates: adaptive evolution in concert with other mitochondrial proteins? J. Mol. Evol., 47: 249-257.
PubMed - Arnason, U., J.A. Adegoke, K. Bodin, E.W. Born and Y.B. Esa et al., 2002. Mammalian mitogenomic relationships and the root of the eutherian tree. Proc. Natl. Acad. Sci. USA., 99: 8151-8156.
CrossRefPubMedDirect Link - Brandon-Jones, D., A.A. Eudey, T. Geissmann, C.P. Groves and D.J. Melnick et al., 2004. Asian primate classification. Int. J. Primatol., 25: 97-164.
Direct Link - Caine, L., G. Lima, L. Pontes, D. Abrantes, M. Pereira and M.F. Pinheiro, 2006. Species identification by cytochrome b gene: Casework samples. Int. Cong. Ser., 1288: 145-147.
CrossRef - Chaveerach, A., T. Tanee, N. Sattayasai, A. Tanomtong, S.A. Suarez and S. Nuchadomrong, 2007. Genetic relationships of langur species using AFLP markers. Pak. J. Biol. Sci., 10: 1445-1451.
CrossRefPubMedDirect Link - Eizirik, E., W.J. Murphy and S.J. O'Brien, 2001. Molecular dating and biogeography of the early placental mammal radiation. J. Hered., 92: 212-219.
PubMedDirect Link - Gibson, A., V.G. Shankar, P.G. Higgs and M. Rattray, 2005. A comprehensive analysis of mammalian mitochondrial genome base composition and improved phylogenetic methods. Mol. Biol. Evol., 22: 251-264.
PubMed - Gonder, M.K., T.R. Disotell and J.F. Oates, 2006. New genetic evidence on the evolution of chimpanzee populations and implications for taxonomy. Int. J. Primatol., 27: 1103-1127.
CrossRef - Groves, C., 2001. Primate Taxonomy. Smithsonian Institution Press, Washington, ISBN-10: 156098872X.
Direct Link - Hayasaka, K., C.G. Skinner, M. Goodman and J.L. Slightom, 2003. The γ-globin genes and their flanking sequences in primates: findings with nucleotide sequences of capuchin monkey and tarsier. Genomics, 18: 20-28.
PubMedDirect Link - Hudelot, C., V.G. Shankar, H. Jow, M. Rattray and P.G. Higgs, 2003. RNA-based phylogenetic methods: Application to mammalian mitochondrial RNA sequences. Mol. Phylogenet. Evol., 28: 241-252.
PubMed - Murayama, M.I., O. Takenaka and Y. Murayama, 1998. Origin and divergence of tandem repeats of primate D4 dopamine receptor genes. Primates, 39: 217-224.
CrossRef - Irwin, D.M., T.D. Kocher and A.C. Wilson, 1991. Evolution of the cytochrome b gene of mammals. J. Mol. Evol., 32: 128-144.
CrossRefPubMedDirect Link - Jaworski, C.J., 1995. A reassessment of mammalian α A-crystalline sequences using DNA sequencing: implications for anthropoid affinities of tarsier. J. Mol. Evol., 41: 901-908.
CrossRef - Jow, H., C. Hudelot, M. Rattray and P.G. Higgs, 2002. Bayesian phylogenetics using an RNA substitution model applied to early mammalian evolution. Mol. Biol. Evol., 19: 1591-1601.
Direct Link - Karanth, K.P., L. Singh, R.V. Collura and C.B. Stewart, 2008. Molecular phylogeny and biogeography of langurs and leaf monkeys of South Asia (Primates: Colobinae). Mol. Phylogenet. Evol., 46: 683-694.
Direct Link - Kocher, T.D., W.K. Thomas, A. Meyer, S.V. Edwards, S. Paabo, F.X. Villablanca and A.C. Wilson, 1989. Dynamics of mitochondrial DNA evolution in animals: Amplification and sequencing with conserved primers. Proc. Natl. Acad. Sci. USA., 86: 6196-6200.
PubMedDirect Link - Lim, L.S., K.C. Ang, M.C. Mahani, A.W. Shahrom and B.M. Md-Zain, 2010. Mitochondrial DNA polymorphism and phylogenetic relationships of proto malays in peninsular Malaysia. J. Biol. Sci., 10: 71-83.
CrossRefDirect Link - Martin, R.D., 1990. Primate Origins and Evolution: A Phylogenetic Reconstruction. Chapman and Hall Ltd., London, 0412127105.
Direct Link - Masters, J.C., M. Boniotto, S. Crovella, C. Roos, L. Pozzi and M. DelPero, 2007. Phylogenetic relationships among the Lorisoidea as indicated by craniodental morphology and mitochondrial sequence data. Am. J. Primatol., 69: 6-15.
CrossRefPubMedDirect Link - Matsui, A., F. Rakotondraparany, I. Munechika, M. Hasegawa and S. Horai, 2009. Molecular phylogeny and evolution of prosimians based on complete sequences of mitochondrial DNAs. Gene, 441: 53-66.
CrossRef - Md-Zain, B.M., J.C. Morales, M.N. Hassan, J. Abdul, M. Lakim, J. Supriatna and D.J. Melnick, 2008. Is Presbytis a distinct monophyletic genus: Inferences from mitochondrial DNA sequences. Asian Primates J., 1: 26-36.
Direct Link - Merker, S. and C.P. Groves, 2006. Tarsius lariang: A new primate species from western central sulawesi. Int. J. Primatol., 27: 465-485.
CrossRef - Merker, S., C. Driller, I.N. Arboleda and H. Zischler, 2008. Polymorphic microsatellite markers for Philippine tarsiers (Tarsius syrichta). Conserv. Genet., 9: 1349-1351.
CrossRef - Murphy, W.J., E. Eizirik, W.E. Johnson, Y.P. Zhang, O.A. Ryder and S.J. O'Brien, 2001. Molecular phylogenetics and the origins of placental mammals. Lett. Nature, 409: 614-618.
CrossRefDirect Link - Osterholz, M., L. Walter and C. Roos, 2008. Phylogenetic position of the langur genera Semnopithecus and Trachypithecus among Asian colobines and genus affiliations of their species groups. BMC Evol. Biol., 8: 58-58.
CrossRefDirect Link - Poux, C. and E.J. Douzery, 2004. Primate phylogeny evolutionary rate variations and divergence times: a contribution from the nuclear gene IRBP. Am. J. Phys. Anthropol., 124: 1-16.
PubMedDirect Link - Raina, S.Z., J.J. Faith, T.R. Disotell, H. Seligmann, C.B. Stewart and D.D. Pollock, 2005. Evolution of base-substitution gradients in primate mitochondrial genomes. Genome Res., 15: 665-673.
Direct Link - Ross, C., B. Williams and R.F. Kay, 1998. Phylogenetic analysis of anthropoid relationships. J. Hum. Evol., 35: 221-306.
Direct Link - Roos, C., T. Nadler and L. Walter, 2008. Mitochondrial phylogeny, taxonomy and biogeography of the silvered langur species group (Trachypithecus cristatus). Mol. Phylogenet. Evol., 47: 629-636.
Direct Link - Schmitz, J., M. Ohme and H. Zischler, 2001. SINE insertions in cladistic analyses and the phylogenetic affiliations of Tarsius bancanus to other primates. Genetics, 157: 777-784.
Direct Link - Schmitz, J., M. Ohme and H. Zischler, 2002. The complete mitochondrial sequence of Tarsius bancanus: Evidence for an extensive nucleotide compositional plasticity of primate mitochondrial DNA. Mol. Biol. Evol., 19: 544-553.
Direct Link - Shoshani, J., C.P. Groves, E.L. Simons and G.F. Gunnell, 1996. Primate phylogeny: Morphological vs molecular results. Mol. Phylogenet. Evol., 5: 102-154.
Direct Link - Stanhope, M.J., W.J. Bailey, J. Czelusniak, M. Goodman and J.S. Si et al., 1993. A Molecular View of Primate Supraordinal Relationships from the Analysis of both Nucleotide and Amino Acid Sequences. In: Primates and Their Relatives in Phylogenetic Perspective, MacPhee, R.D.E. (Ed.). Plenum Press, New York, pp: 251-292.
- Steiper, M.E., 2006. Population history, biogeography, and taxonomy of orangutans (Genus: Pongo) based on a population genetic meta-analysis of multiple loci. J. Human Evol., 50: 509-522.
PubMedDirect Link - Tanee, T., A. Chaveerach, R. Sudmoon, A. Tanomtong and P. Mokkamul, 2009. Genetic relationships of cercopithecidae in thailand as inferred from rDNA ITS regions. J. Biol. Sci., 9: 51-56.
CrossRefDirect Link - Uddin, M., J.C. Opazo, D.E. Wildman, C.C. Sherwood, P.R. Hof, M. Goodman and L.I. Grossman, 2008. Molecular evolution of the cytochrome c oxidase subunit 5A gene in primates. BMC Evol. Biol., 8: 8-8.
PubMedDirect Link