
Jacob H. Jacob
Department of Biological Sciences, Faculty of Sciences, Al al-Bayt University, Mafraq, Jordan
Sohail Alsohaili
Department of Biological Sciences, Faculty of Sciences, Al al-Bayt University, Mafraq, Jordan
Journal of Biological Sciences
Year: 2010 | Volume: 10 | Issue: 2 | Page No.: 162-165







ABSTRACT
In this study, we aimed at isolating fungal strains from the indoor environment that tolerate or/and degrade phenol as sole carbon and energy source. Subsequently, we isolated two fungi (IF2 and IF3, identified as Alternaria sp. and Penicillium sp., respectively) showed the ability to degrade phenol as sole carbon and energy source in mineral medium. Growth with phenol was monitored by increase in biomass measured as dry weight and depletion of phenol measured by gas chromatography. Both strains were also grown with lactose as reference. The IF2 showed higher phenol degradation capacity (18.6 μg L-1) than IF3 (5.4 μg L-1).
PDF Abstract XML References Citation
How to cite this article
Jacob H. Jacob and Sohail Alsohaili, 2010. Isolation of Two Fungal Strains Capable of Phenol Biodegradation. Journal of Biological Sciences, 10: 162-165.
DOI: 10.3923/jbs.2010.162.165
URL: https://scialert.net/abstract/?doi=jbs.2010.162.165
DOI: 10.3923/jbs.2010.162.165
URL: https://scialert.net/abstract/?doi=jbs.2010.162.165
INTRODUCTION
Phenol is a monosubstituted aromatic compound and is recognized as major environmental pollutant. It is produced through natural and anthropogenic processes. It is found in nature as a constituent of coal tar and is also formed during natural decomposition of organic material (WHO, 1994; EPA, 2002). However, the major part of phenol present in the environment is due to industrial use. Currently, the largest use of phenol is as an intermediate in the production of phenolic resins, which are used in the plywood, adhesive, construction, automotive and appliance industries (EPA, 2002).
Phenol poses several toxic cellular effects (EPA, 2002). Membrane toxicity is the most pronounced effect (Sikkema et al., 1995). Phenol toxic effect on membranes is mainly due to its lipophilicity. Thus, phenol tends to accumulate into the lipophilic layer of the cell membrane disturbing its integrity. Hence, it is not surprising to use phenol in disinfectants (EPA, 2002).
Despite phenol toxicity, a number of microorganisms tolerates and degrades it as source of carbon and energy. Subsequently, several enrichment cultures were obtained and many pure cultures were isolated. Among these isolates are prokaryotic and eukaryotic microorganisms. In the world of prokaryotes, phenol degradation was demonstrated under both conditions, aerobic (Baraniecki et al., 2002; Bej et al., 2000; Kotturi et al., 1991) and anaerobic conditions (Shinoda et al., 2000; Khoury et al., 1992; Tschech and Fuchs, 1987; Healy and Young, 1978).
In respect to biodegradation of phenol by eukaryotes, phenol has been shown to be degraded by several fungal strains including mold strains (Leitao et al., 2007; Bergauer et al., 2005; Hofrichter and Scheibner, 1993) and yeasts (Fialova et al., 2004; Santos and Linardi, 2001). The commonly isolated fungal genera include Fusarium, Alternaria and Penicillium (Leitao et al., 2007; Hofrichter and Scheibner, 1993) and most of these isolates originated from soil.
We aimed in this study initially at exploring the capacity of indoor fungi to degrade phenol to demonstrate the use of sick walls as source of aerobic eukaryotic aromatic-compounds degraders. Since, the paint-coated walls are expected to harbor various aromatic compounds, any growing microorganism is expected to be either tolerant to the toxic properties of these aromatics or posses the potential for degrading such compounds. Such degraders are of particular interest in the field of mycobioremediation.
MATERIALS AND METHODS
Fungi sampling and cultivation: Isolated fungi originated from painted sick-building walls located in Irbid, Jordan during March, 2009. The samples were retrieved by applying the adhesive side of a clear cellotape onto surfaces showing different molds. Molds were transferred to plates containing the growth medium, Potato-Dextrose Agar (PDA). Molds were then incubated at room temperature for at least one week in dark. Growth was inspected daily. Based on colony morphology, three different mold isolates could be obtained. Microscopic observation revealed that they belong to two genera. The identification was guided by comparing the key characteristics to those explained by Watanabe (2002). The two isolates were selected for further experiments, i.e., preparation of spore suspension and quantitative analysis of phenol.
Preparation of spore suspension: Fungal isolates were subcultured on PDA slants (50 mL screw capped bottles containing 20 mL PDA) and incubated for one week. Spores were then suspended in 10 mL 0.8% Tween 80.
Cultivation with phenol and lactose as sole carbon and energy source: Mineral medium lacking carbon and energy sources was prepared to test phenol degradation capacity. The medium contained per liter distilled water: CaCO3 (4 g), MgSO4 (2 g), KH2PO4 (7 g), NaCl (0.5 g), NH3SO4 (6 g), CuSO4 (0.5 g), ZnSO4 (0.5 g), NaSO4 (0.5 g), MnSO4 (0.2 g) and FeSO4 (0.1 g). The medium was dispensed into 250 screw-capped flasks. Each flask contains 100 ml mineral medium. The medium was then supplemented with 10 and 20 μg L-1 liquid phenol as sole carbon and energy source. Parallel flasks containing mineral medium supplemented with lactose (20 g L-1) were also included in the experiment. Additionally, an abiotic control containing only phenol was also included. Complete mineral media containing either phenol or lactose were inoculated with 1 mL of spore suspension of the two selected mold isolates. Duplicate cultures were incubated in shaking incubator (50 rpm) at room temperature over 14 days. Samples (0.5 mL) were retrieved from phenol-supplemented cultures that showed mycelia growth and increase in biomass for GC analysis by the means of sterile syringes.
Biomass measurements: Growth of the isolated fungi was monitored by increase in mycelial biomass measured as dry weight. Semi-permeable filter papers were dried in oven for at least 2 h at 50°C and weighed. Cultures were passed through the pre-weighed dry filter papers under vacuum. The loaded wet filter papers were allowed to dry overnight in oven at 50°C. Dry weight was then estimated by balancing.
Quantitative analysis of phenol by gas chromatography: Concentrations of phenol were determined by Gas Chromatography (GC). The used GC system was GCMS QP5050A (Shimadzu), ionization mode EI. The used column was TRB-1MS (30 mx0.25 mmx0.25 μm). GC program was 60-350 with rate of 35°C min-1 and hold for 15 min. Total flow was 20.6 and the column pressure was 100 kPa with split mode (1:10). Injection temperature was 220°C and detector temperature was 250°C.
RESULTS
Isolation and characterization of fungal strains: Based on colony morphology and microscopic analysis, two fungal strains; IF2 and IF3, were isolated. IF2 and IF3 were identified as Alternaria sp. and Penicillium sp., respectively.
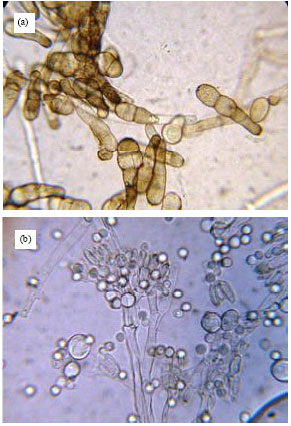 | |
| Fig. 1: | Light micrograph of the fungal strains (a) IF2 and (b) IF3. Image is magnified 1000x |
Alternaria sp. is characterized by pale brawn conidiophores. Conidia are cylindrical and catenulate with walls, while penicillium sp. showed hyaline, erect conidiophores with hyaline round conidia (Fig. 1a, b).
Mycelial growth utilizing phenol and lactose: The ability of the fungal isolates to grow with phenol as sole energy and carbon source was first monitored by following mycelia growth. Cultures of IF2 containing 10 and 20 μg L-1 phenol showed an increase in mycelial biomass after 14 days of incubation (Table 1). However, IF3 showed increase in biomass only in cultures containing 10 μg L-1. Increase in biomass indicates the ability of strains to degrade phenol even though IF2 was able to tolerate higher concentration. The two strains were also grown with lactose as sole carbon and energy source as reference. A significant growth was obtained in case of lactose. The biomass obtained in lactose-containing cultures was always higher than that obtained in phenol-containing cultures (Table 1). It is about the 2 folds in case of IF2 and about 3 folds in case of IF3.
Phenol consumption: Cultures of IF2 and IF3 that showed mycelial growth were further analyzed by gas chromatography to demonstrate phenol consumption over the course of growth.
| Table 1: | Dry weights of mycelia of the two fungal strains IF2 and IF3 grown with lactose and phenol |
 | |
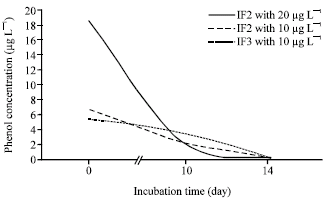 | |
| Fig. 2: | Phenol consumption by cultures of IF2 growing with 20 μg L-1, 10 μg L-1 and IF3 growing with 10 μg L-1 |
A significant decrease in phenol concentration was detected in these cultures over the time of incubation (Fig. 2). The parallel decrease in phenol concentration and the increase in mycelial growth indicate the phenol degradation capacity of IF2 and IF3.
DISCUSSION
We isolated two fungi: IF2 and IF3 identified as Alternaria sp. and Penicillium sp., respectively. The two strains were obtained from indoor environment and showed the capacity to grow with phenol as sole energy and carbon source. The capacity of Alternaria and Penecillium to degrade aromatic compounds was reported by Nambudiri et al. (1970) and Leitao et al. (2007). However, phenol-degrading Penecillium was more frequently reported in the literature. Most of Penecillium isolates originated from soil samples. For example, the halotolerant Penicillium Chrysogenum was isolated from soil samples of salt mine (Leitao et al., 2007) and Penicillium strain Bi 7/2 was isolated from contaminated soil (Hofrichter and Scheibner, 1993). The latter strain was also able to degrade other aromatic compounds. However, among the early work that demonstrated the mycobiodegradability of phenolic compounds was that of Lyr in 1963. Hammel (1995) showed that the fungus Trametes versicolor was able to degrade polychlorinated phenols.
Present findings support the findings of others about the biodegradability of phenol by fungi. Most of phenol-degrading fungi reported in the literature, however, originated from soil, whereas our strains originated from indoor environment.
Growth of IF2 with phenol concentration up to 18.6 μg L-1 (measured in the medium) and IF3 with less concentration (5.4 μg L-1, measured in the medium) indicates that IF2 has higher tolerance or/and degradation capacity than IF3.
The toxic effect of phenol on both strains was apparent when the growth yields of cultures of both strains growing with lactose as sole carbon and energy source with those growing with phenol over the same time of incubation were compared assuming no substrate limitation. However, we primarily prepared lactose-containing cultures to check the viability of spore suspension and the suitability of mineral growth medium.
At the cellular level, the most notable effect of phenol is directed to cell membranes. Membrane-directed toxicity of aromatic compounds correlates with the logarithm of its partition coefficient with n-octanol and water (log Pow), which is used as a measure of the relative lipophilicity of a compound. Phenol has a log Pow of 1.46. Organic compounds with a logPow between 1 and 4 generally have a high affinity for cellular membranes and interfere with the functions of the cytoplasmic membrane (Sikkema et al., 1995). This group of compounds also includes the toxic BTEX (benzene, toluene, ethylbenzene, xylene) compounds.
Exploring the diversity of aerobic aromatic-compound biodegraders broaden the organism basis for studies interested in the diversity of aromatic-compounds biodegradation pathways and bioremediation applications.
Phenol degradation pathways were studied in certain prokaryotic and eukaryotic microorganisms. In fungi, phenol was found to be initiated by either ortho or meta cleavage (Leitao et al., 2007; Leonard and Lindley, 1998; Muller and Babel, 1994) where phenol is degraded to catechol. Both pathways have a phenol hydroxylase in the first step of degradation, but in the second degradation step, eukaryotes generally utilize catecol 1, 2-(ortho) dioxygenase and prokaryotes use 2, 3-(meta) dioxygenase (Leitao et al., 2007; Muller and Babel, 1994; Neujahr and Gaal, 1973). On the basis of enzymatic studies, two pathways were proposed for the metabolism of phenol in Aspergillus fumigatus. In one route, phenol undergoes ortho-hydroxylation to give catechol; in the other, phenol is hydroxylated in the para-position to produce hydroquinone, which is converted to 1, 2, 4-trihydroxybenzene and then, by the action of 1, 2 4-trihydroxybenzene dioxygenase to maleylacetate (Leitao et al., 2007).
CONCLUSION
We isolated two fungal strains from indoor environment with phenol biodegradation capacity. This demonstrates that some indoor fungal strains tolerate phenol and can degrade it as source of carbon and energy.
ACKNOWLEDGMENTS
The authors are grateful to Mr. Muhannad Masad from the Institute of Earth and Environmental Sciences at Al al-Bayt University for his help in GC phenol measurements. The authors are also grateful to Mr. Abdulla Shra'ah and Ms. Eman Al-rgaibat from Biology Department at Al al-Bayt University for their help in fungi cultivation.
REFERENCES
- Bej, A.K., D. Saul and J. Aislabie, 2000. Cold-tolerant alkane-degrading Rhodococcus species from Antarctica. Polar Biol., 23: 100-105.
CrossRef - Bergauer, P., P. Fonteyne, N. Nolard, F. Schinner and R. Margesin, 2005. Biodegradation of phenol and phenol-related compounds by psychrophilic and cold-tolerant alpine yeasts. Chemosphere, 59: 909-918.
CrossRef - Fialova, A., E. Boschke and T. Bley, 2004. Rapid monitoring of the biodegradation of phenol-like compounds by the yeast Candida maltosa using BOD measurements. Int. Biodet. Biodeg., 54: 69-76.
CrossRef - Healy, J. and L. Young, 1978. Catechol and phenol degradation by a methanogenic population of bacteria. Applied Environ. Microbiol., 35: 216-218.
Direct Link - Hofrichter M. and K. Scheibner, 1993. Utilization of aromatic compounds by the Penincillium strain Bi7/2. J. Basic Microbiol., 33: 227-232.
CrossRef - Khoury, N., W. Dott and P. Kampfer, 1992. Anaerobic degradation of phenol in batch and continuous cultures by a denitrifying bacterial consortium. Applied Microbiol. Biotechnol., 37: 524-528.
CrossRef - Kotturi, G., C.W. Robinson and W.E. Inniss, 1991. Phenol degradation by a Psychtrotrophic strain of Pseudomonas putida. Applied Microbial. Biotechnol., 34: 539-543.
CrossRef - Leitao, A.L., M.P. Duarte and J.S. Oliveira, 2007. Degradation of phenol by a halotolerant strain of Penicillium chrysogenum. Int. Biodeter. Biodeg., 59: 220-225.
CrossRef - Leonard, D. and N. Lindley, 1998. Carbon and energy flux constrains in continuous cultures of Alcaligenes eutrophus grown on phenol. Microbiology, 144: 241-248.
CrossRef - Muller, R. and W. Babel, 1994. Phenol and its derivatives as heterotrophic substrates for microbial growth and energetic comparison. Applied Microbiol. Biotech., 42: 446-451.
CrossRef - Nambudiri, A., R. Subba and B. Janardhana, 1970. Metabolism of aromatic compounds by an Alternaria species. Phytochemistry, 9: 687-693.
Direct Link - Neujahr, H. and A. Gaal, 1973. Phenol hydroxylase from yeast purification and properties of the enzymes from Trichosporon cutaneum. Eur. J. Biochem., 35: 386-400.
PubMed - Santos, V. and V. Linardi, 2001. Phenol degradation by yeasts isolated from industrial effluents. J. Gen. Applied Microbiol., 47: 213-221.
Direct Link - Shinoda, Y., Y. Sakai, M. Ue, A. Hiraishi and N. Kato, 2000. Isolation and characterization of a new denitrifying Spirillum capable of anaerobic degradation of phenol. Applied Environ. Microbiol., 66: 1286-1291.
Direct Link - Sikkema, J., J.A. de Bont and B. Poolman, 1995. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Mol. Biol. Rev., 59: 201-222.
PubMedDirect Link - Tschech, A. and G. Fuchs, 1987. Anaerobic degradation of phenol by pure cultures of newly isolated denitrifying pseudomonads. Arch. Microbiol., 148: 213-217.
CrossRefPubMedDirect Link