
Najari Sghaier
Laboratoire dElevage et de la Faune Sauvage des R6gions Arides et Desertiques, Institut des Regions Arides, M6denine, Tunisie
Gaddour Amor
Laboratoire dElevage et de la Faune Sauvage des R6gions Arides et Desertiques, Institut des Regions Arides, M6denine, Tunisie
Ouni Mabrouk
Laboratoire dElevage et de la Faune Sauvage des R6gions Arides et Desertiques, Institut des Regions Arides, M6denine, Tunisie
Abdennabi Mouldi
Laboratoire dElevage et de la Faune Sauvage des R6gions Arides et Desertiques, Institut des Regions Arides, M6denine, Tunisie
Ben Hamouda Mohamed
Institution de Recherche et d Enseignement Superieur Agricole de Tunisie, Alain Savary, Tunis, Tunisia
Journal of Biological Sciences
Year: 2007 | Volume: 7 | Issue: 6 | Page No.: 1005-1016







ABSTRACT
About 5000 individual weights issued from periodical survey of 722 indigenous kids in 8 herds and during 4 years were analyzed in the present study. The aim is to identify the non genetic effects upon the growth curve of local goat population raised under arid conditions as a necessary step to improve kids` meat production on range lands. The Gompertz model was adopted to assess individual kids` growth curves. A non linear regression iterative procedure allows estimating 688 kids curves parameters and convergence criteria was 10-8. For each kid, curve parameters, asymptotic weight, inflexion age and weight, were considered as quantitative traits. A GLM procedure was applied to decompose the total variance of the kid`s traits. A means comparison test (SNK, α = 0.05) was applied to identify homogenous class by factor. Results show that the GLM determination coefficient remains lower than 67% of all studied traits due to the observed data structure. All traits seem to be affected by the significant effects (p< 0.001 or 0.05) of the factors related to the restrictions and the irregularity of the technical and natural environment of pastoral husbandry. Specially, growth traits varied with year, herd, month of birth and natural region. Herd mobility contributes to stamp climatic stresses and resources irregularities. Equally, the year factor plays an important role upon the kids weight and it evolution till 6 months age. Kids growth curve loses its S-shape and becomes linear under harsh conditions of dry years. Also, the non-genetic factors impact increases with kids age and requirements. The sex, birth mode and mother age acts only during first 2 months age. So, arid environment affects both quantitatively and qualitatively individual kid`s growing behaviour and have to be considered for local goat rational genetic improvement modeling and planning.
PDF Abstract XML References Citation
How to cite this article
Najari Sghaier, Gaddour Amor, Ouni Mabrouk, Abdennabi Mouldi and Ben Hamouda Mohamed, 2007. Non Genetic Factors Affecting Local Kids` Growth Curve under Pastoral Mode in Tunisian Arid Region. Journal of Biological Sciences, 7: 1005-1016.
DOI: 10.3923/jbs.2007.1005.1016
URL: https://scialert.net/abstract/?doi=jbs.2007.1005.1016
DOI: 10.3923/jbs.2007.1005.1016
URL: https://scialert.net/abstract/?doi=jbs.2007.1005.1016
INTRODUCTION
Local goat pastoral breeding mode remains with a particular importance in the Tunisian arid region (Ouled Belgacem, 2006; Ouni, 2006), where were raised more than 60% of the national caprine herd estimated to 1500000 goats (DGPA, 2005; FAO, 2006). In the traditional breeding mode of the indigenous goat population, the main product of ambulant herds is the meat production (Kaosgey et al., 2006; Mahjoub et al., 2005) and kids were not weaned till their separation in summer when they were slaughtered (Najari et al., 2007; Gaddour, 2005). In southern Tunisia, kids' meat represents about 75% of the regional meat production (Najari, 2003). Regarding the birth season, the age of market class kids varies between 4 and 8 months. Consequently, kid’s growth during this early age represents an important phenotype conditioning the final cash flow of the ambulant herds (Gaddour et al., 2006; Ouni, 2006). Under pastoral harsh conditions with restricted and irregular resources, goat production is influenced more by non-genetic factors because of the extensive grazing management (Mahjoub et al., 2005). The kid’s genome, acquired at the fertilization, defines the target mature size and the growing potential (Simondon, 1991; Walkden-Brown et al., 1994; Schinckel and de Lange, 1996); whereas environmental factors affect the genotype expression and modify the resulting phenotype (Knnan et al., 2006). While the genotype remains invariable, non-genetic factors vary during the animal productive life illustrating natural and technical fluctuations (Genandoy et al., 2002). Like other animal phenotypes, growth curve parameters and shape change by all factors affecting the growth rather than the individual genetic potential differences (Alexandre et al., 1997a; Bocquier et al., 1998; Oltenacu, 1999). De Lange et al. (1998) suggest establishing growth curve by production factors to consider the variation in data due to the non genetic factors’ effects.
Rather than the growth study utilities to improve herd management (Gipson and Wildeus, 1994; Genandoy et al., 2002), its importance is crucial in pastoral husbandry when animals strive to express their genetic potentialities (Najari et al., 2002; Kaosgey et al., 2006). Under arid conditions, besides the non-genetic factors classic effects, a specific qualitative action upon growth kids was verified (Najari et al., 2002). Stresses and restrictions can avoid the genetic potential expression even though for rustic populations. This aspect can prevents also the verification of the quantitative genetic hypothesis and the methodologies application. Thus, to establish breeding plan or to estimate genetic parameters, statistical model should account correctly all environmental effects and interactions (Schinckel and De Lange, 1996; De Lange et al., 1998).
This study aims to estimate the natural and technical non-genetic effects on body weights and growth curve parameters of indigenous kids raised under pastoral conditions in the Tunisian arid region. Also, we purpose understanding kids' growing behaviour of indigenous kids towards arid environment factors by the analysis of a large data set collected within ambulant herds. Rather than the local population characterization, the results may improve the husbandry policy to optimise the herd production and land conservation considering the serious decertification risks in the studied area.
MATERIALS AND METHODS
Study zone: Data were collected from the arid region of Tunisia as ecologically defined by Floret and Pontanier (1982). The climate is arid Mediterranean, hard and precarious (Ouled Belgacem, 2006). With an annual average of 140 mm, precipitation presents a large spatial and seasonal variation. The arid zone is actually the most important of rangelands of the country and is mainly used for extensive camel and small ruminants grazing (Nasr et al., 2000).
Animal material: The indigenous goat population constitutes an animal group adapted to the arid rangelands harsh conditions (Najari et al., 2006). The complex adaptation criteria were acquired by a long natural and human selection process under local hard conditions (Najari, 2005). The indigenous goat population shows a large variability both in morphology and performances (Najari, 2003). Characteristics of the population include the ability to walk long distances, water deprivation resistance and good kidding ability. Fertility rate is about 87% and prolificacy rate varies between 110 and 130% (Najari et al., 2004). Kidding season begins in October and continues till February with a concentration in November and December when 69.2% of kids are born.
Data collection: About 5000 kids' weights were used in the present study. Data is carried out, during four years, from a periodical weighing plan of 722 indigenous kids raised in eight ambulant herds under arid land conditions in the southern Tunisia. Since the start of the kidding period and till summer beginning, kids were weighed once every two or three weeks. The controlled herds are distributed on different ecological zones of the arid region.
Each kid records included goat mother and kid identification, type of birth, date of birth, sex, herd, natural region and kid’s weights measured with the corresponding dates of controls.
Individual growth parameters and weights estimation: Regarding previous studies, (Najari, 2005; Najari et al., 2007), it was concluded that the Gompertz function is the convenient model to assess the indigenous kids’ growth during the first 6 months age. Hence, this model was used to estimate individual curve parameters and kid’s weights at some fixed ages.
According to Schnickel and Graig (2001), the Gompertz function is defined by:
| (1) |
t is the kid age (days); A is the curve asymptote. Parameters b and c adjust both slope and inflexion point. The first derivative, or the growth rate, is defined by:
| (2) |
The coordinates of the point of inflection are ti = c and Wi = A/e.
For each kid, the Gompertz growth curve parameters were estimated by an iterative procedure with zero as the starting value for all parameters. The number of the iteration is fixed less than 1000 and the convergence criterion was set at 10-8 (Najari et al., 2007).
Statistical analysis: The individual growth curve parameters were analysed as quantitative traits. A General Linear Model analysis was applied to decompose the total variance and to apply the F statistical significance test.
The statistical GLM model used with independent variables was as follows:
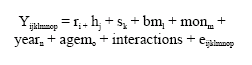 |
Where:
Yijklmnop is the performance analyzed: curve parameters (A, b and c), the age at inflection point and inflection weight.
| ri | = | the geographic zone effect (I = 1, 2 or 3, respectively costal, steppe and mountain zone); |
| hj | = | the herd effect (j = 1,….8); |
| sk | = | the sex effect (k = 1, 2; male or female) ; |
| bml | = | the mode of birth effect (l = 1,2; single or twin); |
| monm | = | the month of birth effect (m = 1,….12); |
| yearn | = | the year of birth effect (n = 1,…,4); |
| agemo | = | the goat mother age effect (o = 1,…,8); |
| and eijklmnopq | = | the model residual. |
Only interactions of two degree were considered.
Except the residual error, all factors and interactions were considered fixed. A SAS GLM procedure was used to resolve the linear equations system (SAS, 1999).
RESULTS AND DISCUSSION
Individual kid’s growth curve fitting: An iterative procedure was applied to estimate individual Gompertz growth curve parameters A, b and c for each kid (Eq. 1; Najari et al., 2007). The convergence was held only for 668 kids; for the rest, the iterative process diverged due to the data structure or insufficiency. In other similar studies (De Lange et al., 1998), the adjustment of individual growth curve do not allows to assess all kids' curves of Spanish breed data. In our case, the arid conditions heterogeneity added to the indigenous population large variability, between herds and years, can explain the difficulties showed for some kids curve adjustment (Najari, 2005). In fact, more the population variability is large more the individual growing behavior can deviate with respect to the population general behavior. Under harsh conditions, some animals can manifest non-sigmoid growth curve. Furthermore, the non-use of limitative values constraints into the iterative procedure can also generate individual parameters with illogic values. The iteration number varied between 6 and 463. The CD presents some low values for some kids’ curves (about 0.01).
Non-genetic factors affecting the indigenous kids’ growth: For all studied traits, the R2 values varied between 0.57 and 0.69 (Table 1). It seems that an important part of the total variability remains not represented into the model. Natural region, herd and year of birth manifest a high significant effect upon all analyzed traits (p<0.01). These three factors appear as the most important through their effects on the forage availability and the arid climatic stresses; whereas, the other factors effects remain varying with respect to the studied trait and the growth stage.
Among the interactions included in the decomposition model, we note the importance of the effect of those including the factor herd (management mode) with the effect of natural factors such as region, year or month (Table 1).
Regarding kid’s sex, birth mode and mother age which were known with a classic impacts upon the early kid growth, it seems that they have only reduced effects under extensive breeding system in an arid land. The influence of such factors is secondary towards the impact of factors conditioning the food availability and herd management under harsh and irregular circumstances. These resultants agree with those presented by Najari (2005).
Ecological zone effects: The natural zone affects kid’s growth through the resources and the herd management not homogenous in studied ecological zone such as steppic, coastal and mountainous zone (Najari et al., 2003; Ouled Belgacem, 2006; Najari et al., 2007). In the coastal zone, the livestock plays a secondary role in the family incomes and the pastoral lands are regressing in favor to oil culture. Whereas, in the second zone, the livestock remains he principal activity and herds graze extensively the vast communal lands and pasture without sensible mobility restrictions. While in mountainous zone, goatherds valorize accidental and vacant lands.
For each ecological zone, the Gompertz equation of kids’ growth was as follow:
Costal zone:Weight (kg) = 16.97*Exp (-Exp (0.6242-0.0132*Age))
Steppe zone:Weight (kg) = 22.16*Exp (-Exp (0.8225-0.0166*Age)
Mountain:Weight (kg) = 22.72*Exp (-Exp (0.74-0.012*Age))
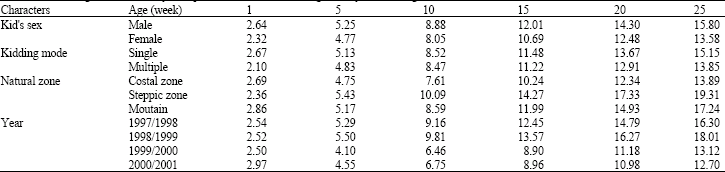
The zone corresponding growth curves are presented on the Fig. 1 and Table 2. The most regular, S-shaped growth curves are observed to the steppe and the mountain. The heaviest asymptotic weight was observed for the mountain kids having 22.72 kg, while, the lowest kid’s weight are those raised in the costal zone.
| Table 1: | Analysis of variance results for kid’s growth curves parameters with respect to non genetic factors |
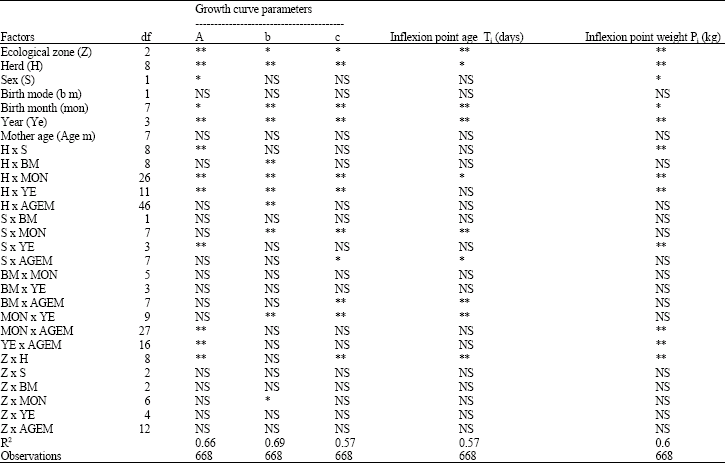 | |
| df: degree of freedom; A: Asymptotic weight (kg); b, c: growth curve parameters; Ti: Kid’s age at inflexion point; Pi: Kid’s weight at inflexion point; R2: Model determination coefficient; NS: Non Significant; *Significant (p<0.05); **Highly significant (p<0.01) | |
| Table 2: | Kids' weights, estimated by Gompertz model, till 25 week age and by some non-genetic factors |
 | |
| Similar effects of the natural and technical resources upon the kids growth was advanced in a numerous studies (Kochapakdee et al., 1994; Ogebe et al., 1995; Hussain et al., 1995, Najari et al., 2005). | |
Herd effect upon kid’s growth: Rather than the classic effect of herd upon animal performances, this non genetic technical factor has a particular importance in the pastoral breeding system of the arid region. The management response toward the resources scarcity and irregularity is determining upon the herd production (Santoir, 1990; Niamir, 1996). One of the pastoral management principles is the herd’s mobility, practiced since the nomadic period. Transhumance allows to valorize distinct pasture according to their vegetal state and thus, to stamp the arid resources restrictions and irregularities. Due to the herd mobility hardness and costs, only little number of herds remains able to practice duly the transhumance to satisfy the animal’s requirements. Also, in this extensive grazing mode, the herder ability remains the principal factor conditioning the herd cash flow (Najari et al., 2004). In fact, the herd management is based on the traditional knowledge to prevent natural stresses and to ensure the best offered conditions to satisfy animal requirements (Niamir, 1996; Najari et al., 2003).
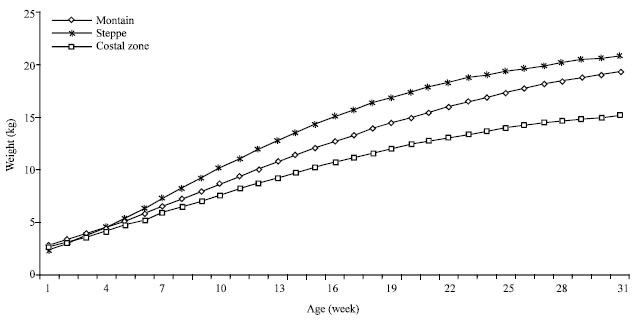 | |
| Fig. 1: | Kids' growth curve adjusted by Gompertz model and by ecological zone |
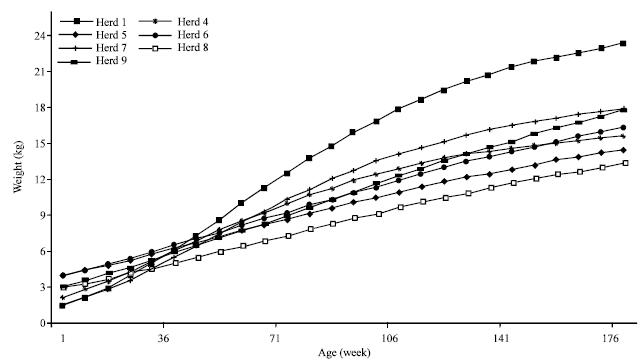 | |
| Fig. 2: | Kids' growth curve adjusted by Gompertz model by goat herds |
Rather than technical management differences, the genetic level varies between herds (Ouni, 2006). Morphologically, some herd specificities were easily revealed on the animals’ morphology due long intra-herd consanguine mating. Traditionally, each family selects its own herd on some visible characters to avoid animals’ mixture on pastures or during the watering days when many herds are gathered (Najari et al., 2006). These simplified selection schemes, can affect productive performances such as kids’ growth.
During the first month age, herds' differences are not enough clear (Fig. 2 and Table 2). Among the nine herds studied, (Fig. 2) kids of the herd number 1 show the best growing performances since 2 months age and till the age of 6 months. The asymptotic weight corresponding to the herd 1 reaches 25.28 kg.
Differences between herds’ performances increase with age to become remarkable after 40 days age (Fig. 2). At the age of 180 days, this difference reaches 17.4 kg.
Note that the interaction herd*region showed a significant effect upon the majority of analyzed traits; the herd management vary with respect to the natural resources of each region. In fact, herd management and the breeding mode differ according to herd importance and the forage availability. Najari (2005) mentioned a similar effects of herds upon kids' weights under harsh conditions.
Year of birth effect upon kids’ growth: Rather than its harsh conditions, the arid environment is essentially defined by its irregular conditions (Ferchichi, 1996; Ouled Belgacem, 2006). During the survey period, annual rainfall ranged between 37 mm for 2000/2001 and 307 mm for 1997/1998. Indeed, this variation is expressed by serious fluctuations of the forage available to animals.
Since early age (Table 1 and 2), the year effect is highly significative (p<0.01). This ambiantal effect becomes higher with the kids’ age which expresses the nutritive requirements increasing. The early kid’s growth remains comparable during the four years till the age of one month; it seems that reduced requirements can be covered by land grass during this young age. When the animal needs are low, we can expect phenotypic differences expressing various genetic potentialities; whereas, at older age and high needs, the phenotypic differences were essentially attributed to the nutritional availability defined by environmental factors. This qualitative aspect of the year effect upon the kid’s growth must be considered to manage genetic improvement plan adapted to the indigenous population and the arid conditions. When the animal’s needs are low, it will be possible to express genetic differences between animal’s phenotypes; which represents the environment behavior for aged animals under very restrictive circumstances.
During the years relatively rained, such as 1997/1998 and 1998/1999, the kids growth curve show the typical S shape with a clear inflexion zone before a rapid tendency of the concave curve which finished by a relative heavy asymptotic weight. Whereas, during the two other difficult years, the curves become linear and not illustrating the typical growing process; this growth curves testify the suffering life of both kids and mother, which strive to realize a reduced growth.
The year effect upon the kids’ growth was verified by several authors (Walkden-Brown et al., 1994; Gipson and Wildeus, 1994; Das et al., 1996; Alexandre et al., 1997c; Bocquier et al., 1998). All authors agree on the year impact witch acts directly on kid’s nutritional resources and indirectly by the bias of the mother milk production (Fig. 3).
The significant effect of the herd* year interaction (Table 1) testifies a different response of herds towards annual conditions. When some herds remain able to stamp the drastic conditions by the transhumance practice or by the herd supplementation; others seem having minor resources to safeguard their herds and remain exploiting the same lands. Thus, the role of the herd and the farmer technical capacity can vary the effect of the year.
Month of birth effect: The growth curves variation according to the month of birth is presented in Fig. 4. The kids early born, during November and December, show a weight evolution which retard to tend to the asymptotic weight. Whereas the curves corresponding to the end of the birth season (February and March) converge rapidly to stabilize the growth. The asymptotic weight for the later births was reduced to 12.45 and 9.85 kg, respectively for March and April’s births. Thus, it seems that indigenous kids grew till the spring end; and the growth rate decreases seriously at summer even if the kids' age is reduced. Also, kids born in the end of birth season are unable to realize heavy weight in spite of their genetic potentialities which can’t be expressed under summer conditions and heat stress. The spring season, judged having the better natural conditions (Floret and Pontanier, 1982; Ouled Belgacem, 2006) correspond to the better growth rate independently with the physiologic growing stage. Thus, having the majority of births during winter season can been considered as an adaptation aspect of the population to agree the period of maximal nutritional needs of the kids growth with the favorable season. Similar kidding season effects have been signaled by many authors (Ogebe et al., 1995). This typical effect of the month of birth led to recommend the reproduction regrouping in the aim to decrease the number of the later births (after February). In fact, this later kids not allow an additive meat production and rather. Equally, their presence in herd till summer complicates the herd mobility and, also, affects negatively the reproductive performances of lactating goats.
The kids’ sex effects upon growth performances: Figure 5 and Table 2 illustrate the growth curve for male and female kids. Since the birth, differences of kids' weights are remarkable for the two sexes. The birth weight, estimated at 2.64 and 2.32 kg, respectively explain the superiority of earlier male weights (Najari, 2005). This male superiority continues during the growth period. The male asymptotic weight was 18.09 kg when the female growth stops at 15.1 kg. Similar sex effect is commonly concluded in several studies (Barbato and Vasilatos-Younken, 1991; Barbato, 1991; Anthony et al., 1991a; Banda et al., 1993; Bathaei and Leroy, 1996; Alexandre et al., 1997b; Oltenacu, 1999).
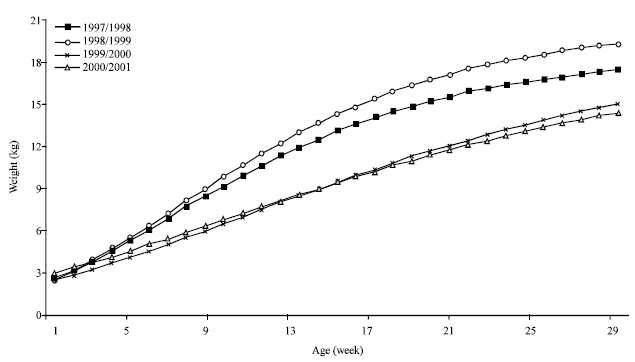 | |
| Fig. 3: | Kids' growth curve adjusted by Gompertz model and by year of birth |
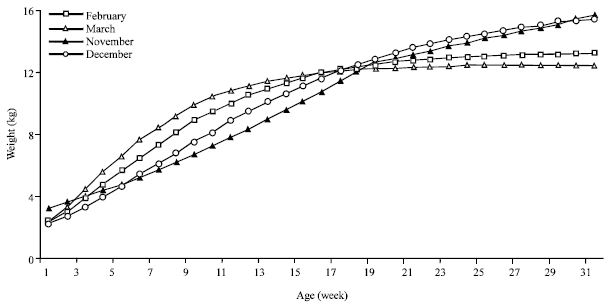 | |
| Fig. 4: | Kids' growth curve adjusted by Gompertz model and by month of birth |
However, the lower asymptotic weights permit to female to reach more rapidly the same physiologic state with respect to the male. In fact, the female kids reach the inflexion point since 37 days of ages, so 5 days before the male kids; the same physiologic state can be represented by the inflexion weight with was, respectively 6.65 and 5.52 kg for male and female kids.
According to these results, we can deduct that female kids can reach the maturity state rapidly and its can began reproductive process since the first year of age. This can be considered as an adaptation criteria of the indigenous population; the priority to guaranty the genetic continuity of the animal group under harsh condition by a rapid reproductive rhythm of young females (Najari, 2005). With low weights and reduced needs, female kids reach rapidly the physiology maturity required to begun the reproduction process since the first year of age. In favourable years, it is common that indigenous female kids can be covered during summer without any problem.
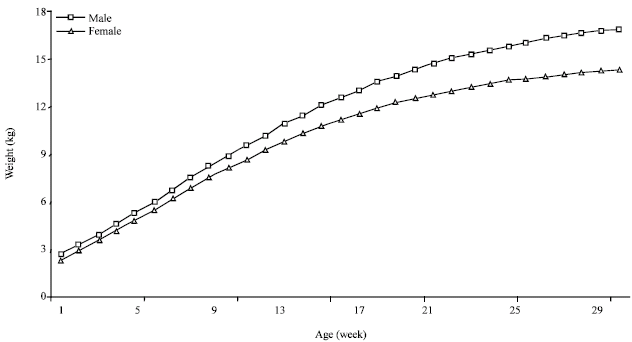 | |
| Fig. 5: | Kids' growth curve adjusted by Gompertz model and by kids' sex |
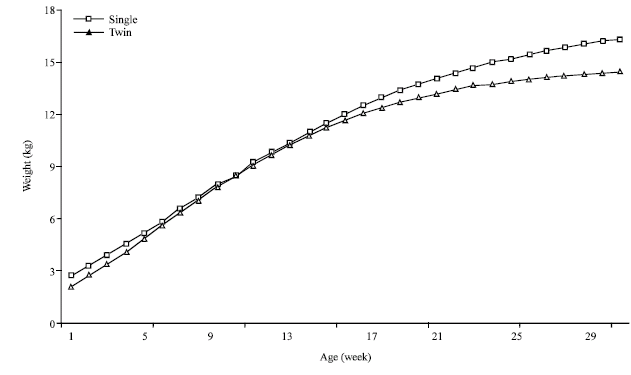 | |
| Fig. 6: | Kids' growth curve adjusted by Gompertz model and by kids' birth mode |
The often local breeds small body size allows native populations to success the reproduction process and by which, to guarantee the survival and the genetic continuity of the population even under drastic conditions.
The kidding mode or mother parity effects upon kids’ growth: The effect of this factor is considered classic in the bibliography (Lyatuu et al., 1992; Okello, 1993; Gromela et al., 1998; Gaddour et al., 2007; Ouni, 2006). The single kids have normally better conditions to realize heaviest weights. Alexandre et al. (1997a) indicate that the difference between simple and double could represent until 15% of weight at birth. The simple births have some best conditions to achieve some heavier weights (Alexandre et al., 1997b; Oltenacu, 1999). In our case (Fig. 6 and Table 2), this difference isn’t clearly showed till 3 months age which concord normally with the spring favourable season.
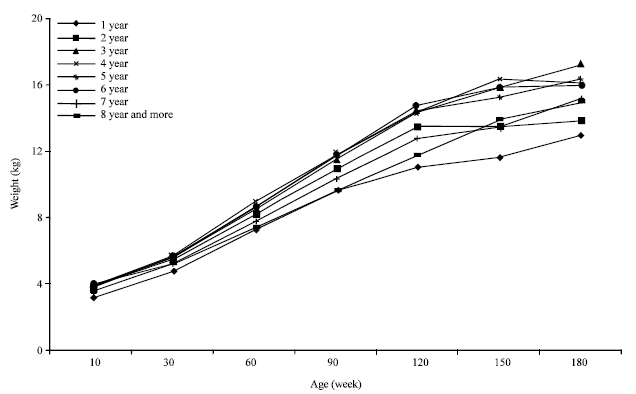 | |
| Fig. 7: | Kids' growth curve adjusted by Gompertz model and by kids' mother age (years) |
It seems that the restricted indigenous goat milk production (Najari et al., 2000) avoid the expression of the single kids superiority. In fact, at earlier age, the kid’s nutrition is essentially based on the mother milk which appears insufficient to express differences between single and multiple kids growing behaviour. Single kids have to wait the grazing age to converge to a heaviest asymptotic weight. To express genetic superiority, animals need favourable conditions. Such aspect, specific to arid zone, was mentioned by Najari (2005).
The mother age effect upon kids’ growth: The mother age effect upon the kids’ growth is illustrated in Fig. 7. The mother age represents essentially the lactation range with which varies the milk potentialities. In fact, goat lactation evolves to a maximum reached at the 3rd or 4th lactation (Ba diao et al., 1994). The relation between kids’ growth and the mother age was demonstrated in several studies (Wiggans, 1992; Pinkerton, 1994). In our case, the lowest kids' performances were registered for the kids of goats having the first lactation, followed by the category of the goat two years aged and the oldest goats. Goats aged between 3 and 7 years seem to allow the better kids growth; it seems that the forage scarcity avoid to distinguish differences into this group.
The variance analysis shows a significant effect of some interaction intervening the mother age (Table 1). The effect expression of other factors, such as herd, sex, birth mode seems to be conditioned by the mother age which can stamp the impacts of favorable or non favorable conditions. These results confirm those mentioned by Najari et al. (2007).
Regarding previous finding, the kids' growth curves under harsh conditions show some specificities which synthesize the arid environment impacts and the local goat genetic expression towards stresses and food scarcities (Najari et al., 2003; Najari, 2005; Gaddour, 2006; Gaddour et al., 2007; Najari et al., 2007).
CONCLUSIONS
The growing behavior of indigenous kids with respect to non genetic factors, illustrates the adaptation of the local population towards natural and technical conditions. Under arid harsh conditions, all environmental factors act on herd meat production by the bias of the resources variation and the climatic stresses intensity. Scarce, irregular and sparse pastoral resources, added to traditional husbandry factors modulated seriously the kids’ weight and the herd final meat production. Indeed, kids’ weights illustrate an important variation with respect to the environmental factors such as year, herd, region and kidding season. These factors importance seems to be higher when the kids were older and when requirements are considerable. The technical aspects represented by the herd factor, can stamp the impacts of the natural restrictions, although by herder knowledge, the herd mobility and the use of the food supplementation by local by products and barley. The notable regression of the common pastoral area and the number of qualified traditional farmers, will accentuate the effect, of natural conditions upon animals performances.
Other factors, such as kids’ sex, month of birth, mother age act upon kids’ growth dependently to other environmental factors. These later factors need some conditions to manifest their effect upon growth. When the nutritional resources are seriously reduced, the meat production evolution does not manifest individual genetic potentialities. So, an important qualitative affect of non-genetic factors can be illustrated under arid drastic conditions. These important variance aspects can affect the kids' genetic evaluation models which need to apprehend the factors to be corrected. So, the nature of the arid factors effects can avoid the correct definition of the fixed factors. These aspects can induce a bias or increase the prediction error using the classic phenotype modeling for selection plans.
To schedule genetic improvement of local population, some interactions and corrective indexes need to handle more appropriate numeric methodologies and statistical package. In fact, some selection hypotheses seem to be not verified considering indigenous population productive behavior and arid conditions specificities.
REFERENCES
- Alexandre, G., G. Aumont, J.C. Mainaud, J. Fleury and M. Naves, 1999. Productive performances of Guadeloupean Creole goats during the suckling period. Small Rumin. Res., 34: 155-160.
CrossRefDirect Link - Anthony, N.B., D.A. Emmerson and K.E. Nestor, 1991. Research note: Influence of body weight selection on the growth curve of turkeys. Poult. Sci., 70: 192-194.
Direct Link - Barbato, G.F., 1991. Genetic architecture of growth curve parameters in chickens. Theor. Applied Genet., 83: 24-32.
CrossRef - Barbato, G.F. and R.V. Asilatos-Younken, 1991. Sex-linked and maternal effects on growth in chickens. Poult. Sci., 70: 709-718.
PubMed - Bathaei, S.S. and P.P. Leroy, 1996. Growth and mature weight of Mehraban Iranian fat-tailed sheep. Small Rumin. Res., 22: 155-162.
Direct Link - Das, M.S., J.E.O. Rege and S. Mesfin, 1996. Phenotypic and genetic parameters of growth traits of blended goats at Malya, Tanzania. Proceedings of the 3rd Biennial Conference of the African Small Ruminant Research Network, December 5-9, 1994, ILRI., Nairobi, Kenya, pp: 63-70.
Direct Link - Goromela, E.H., I. Ledin and P. Uden, 1996. On-Farm Performance of Dual Purpose Goats and Farmers Attitudes Towards Introduction of Goats in HADO Areas of Kondoa. In: Integrated Farming in Human Development, Dolberg, F. and P.H. Petersen (Eds.). DSR-Forlag, Denmark.
Direct Link - Genandoy, H., T. Sahlu, J. Davis, R.J. Wang, S.P. Hart, R. Puchala and A.L. Goetsch, 2002. Effects of different feeding methods on growth and harvest traits of young Alpine kids. Small Rum. Res., 44: 81-87.
CrossRefDirect Link - Kaosgey, I.S., R.L. Baker, H.M.J. Udo and J.A.M. Van Arendonk, 2006. Successes and failures of small ruminant breeding programmes in the tropics. Small Rum. Res., 61: 13-28.
CrossRef - Kochapakdee, S., W. Pralomkarn, S. Saithanoo, A. Lawpetchara and B.W. Norton, 1994. Grazing management studies with Thai goats: 1. Productivity of female goats grazing newly established pasture with varying levels of supplementary feeding. Asian-Aust. J. Anim. Sci., 7: 289-293.
Direct Link - Mahjoub, O., C.D. Lu, M.S. Hameed, A. Richie, A.S. Al-Halhali and K. Annamalai, 2005. Performance of Omani goats fed diets containing various metabolisable energy densities. Small Rumin. Res., 58: 175-180.
Direct Link - Najari, S., A. Gaddour, M.B. Hamouda, M. Djemali and G. Khaldi, 2007. Growth model adjustment of local goat population under pastoral conditions in Tunisian arid zone. J. Agron., 6: 61-67.
CrossRefDirect Link