
O. Ghasemi Chapi
Agricultural and Natural Resources Research Center of Mazandaran Province, Iran
A.S. Hashemi
Agricultural and Natural Resources University of Sari, Iran
E. Yasari
University of Payam Nour, Iran
G.A. Nematzadeh
Agricultural and Natural Resources University of Sari, Iran
International Journal of Plant Breeding and Genetics
Year: 2008 | Volume: 2 | Issue: 1 | Page No.: 28-34







ABSTRACT
Parent selection, determining combining abilities and genetic variation are important in breeding programs. Diallel crosses, therefore, were tested among six canola varieties (Foseto, Option 500, Goliath, Kristina, Shiralee and PF-7045.90). Thirty hybrids and six parents were planted in a lattice design with 3 replications at Mazandaran Agricultural Researches Center, 2004-2005. Primary analysis of variation showed significant differences between genotypes; hence it seemed possible to study the desired traits through diallel analysis. Results of analysis of variation as per Griffing,s 1st method for the duration of 50% of germination, 50% of emergence, three leaf stage, four leaf stage, the time between three and four leaf stage, primary length of root, length of seedlings and seedling vigor index showed that there were significant differences between genotypes (p<0.01). Mean square of general and specific combining ability and reciprocal crosses, statistically were significant for all traits, but mean square of reciprocal crosses for primary length of root was not significant. Mean square (GCA)/mean square (SCA) ration in all traits was significant owing to additive and non-additive gene effects of above mentioned traits, but non-additive gene effects are more important. Estimation of degree of dominance more that one for traits of studied and low narrow heritability showed that non-additive gene effects plays an important role in genetically control of these traits. Heterosis for all traits was negatively significant. Over all it could be concluded that, cv. Shralee was the best parent and KristinaxShiralee was the best cross for all seedling traits.
PDF Abstract XML References Citation
How to cite this article
O. Ghasemi Chapi, A.S. Hashemi, E. Yasari and G.A. Nematzadeh, 2008. Diallel Analysis of Seedling Traits in Canola. International Journal of Plant Breeding and Genetics, 2: 28-34.
DOI: 10.3923/ijpbg.2008.28.34
URL: https://scialert.net/abstract/?doi=ijpbg.2008.28.34
DOI: 10.3923/ijpbg.2008.28.34
URL: https://scialert.net/abstract/?doi=ijpbg.2008.28.34
INTRODUCTION
Canola (Brassica napus L.) is one of the most important oilseeds in the world for the variation in its oil quality and uses. The remnant of its seeds (meal) after oil extraction contains considerable protein content which can be used in livestock feeds (Yasari et al., 2008).
Seedling germination and vigor are under the influence of many different factors. Seed quality, in this respect, is very important since negative effects of seeds of the low quality cannot be compensated by changing in plant cultivation or protection practices. Seed genetic is an important factor of seed quality and play vital role in germination and seedling vigor.
Seedling vigor is important factor for annual crops since it can help the crop in competition with weeds and causes in crop well establishment. In winter cropping, canola should grew vigorously before rosette (Taleie and Beigi, 1996). Having small size seed and seedling, canola initial growth is very slow hence, threatened by mono and dicotyledon weeds. In order to minimize weed loses and having vigorous plants in rosette before cold winter season commencement, vigorous seedling and is very important and helpful to pass through this critical stage of the plant growth (Yasari et al., 2007).
One of the main responsibilities of plant breeders is to create different population of plant and evaluate and compare them with each other (Brown and Caligari, 2008).
Determination of the genetically structure of canola and combining ability usually resulted in accurate and logic selection in hybridation practices, therefore production of best hybrids can be possible. Availability of these genetically information would be possible via diallel crossing following methods described by of Griffing (1956) and Hayman (1960). The quality and quantity of the genetic parameters like heterosis, gene action, general and specific heritability of parents and their offsprings is possible via diallel analysis (Thiagarajah and Stringam, 1993). Genotype has significant effects on germination components (Latifi et al., 2004). Xiao et al. (1992) showed that the germination percentage of hybrids was more than their parents and hybrids root growth rate was more and leaf and stem were thicker and more developed in canola. Sharma et al. (1992) reported that both additive and non-additive gene effects on the length of radicle, shoot let and seedling vigor and specific heritability for these traits in F2 and F1 were high and intermediate, respectively and there was no allelic interaction for all the traits in Indian mustard (Sharma et al., 1992). Sharma and Singh (1996) studied the combinability of the length of radicle, shoot let and seedling vigor of Indian mustard and figured out that the variation of general and specific combining ability (GCA) and (SCA) for these traits in F1 and F2 and their parents were statistically significant. The gene action of these traits was non-additive and the heritability was also low. Thakara et al. (1994) showed both additive and non-additive effects on the genes of seedling vigor on (Brassica carinata) and pointed out that non-additive gene action is more important. Shi et al. (1991) reported a vigorous heterosis in seedling stage of F1 hybrids. The main aim of the present study therefore, was to evaluate the effects of controller genes of the traits and appraisal of the GCA and SCA and determination of the best parent and cross of each trait.
MATERIALS AND METHODS
Six varieties of canola (Foseto, Option 500, Goliath, Kristina, Shiralee and PF-7045.90) along with F2 generation of diallel crossing in the all possible ways were studied in lattice design with 3 replications at Mazandaran Agricultural Researches Center during 2004-2005 cropping season. The traits comprising the duration of (50% of germination, 3 leaf stage, 4 leaf stage, time between 3 leaf to 4 leaf) in green house (Gharakheil Agricultural Researches Station) and duration of 50% of germination, length of radicle and shoot let and seedling vigor index the ratio of seedling dry weight to seedling wet weight (Shi et al., 1991) in the laboratory of collage of agriculture, Mazandaran University have been evaluated.
In order to evaluate the studied traits in green house experiment number of 100 seeds were put in plot containing field soil which then were covered with 1 cm of mixture of sand and soil. In laboratory, the require seed were put in Petri dishes of 8 cm in diameter in which 20 healthy and vigorous seeds were used. Petri dishes were sterilized with sodium hypo chloride (10%) for 1 min prior to seed accommodation in them. The bottom of petri dishes was covered with butter paper. The petri dishes then were soaked with distilled water after putting seed on them. The seeds with 2 mm of radicle length were considered as germinated seed. The number of days to 50% of germination were calculated and adjusted to the number of hours. The length of radicle, shoot let and seedling vigor for each sample was calculated one week after sowing. Data was analyzed using triple lattice design and diallel variation analysis and determination of genetic parameter, like heterosis, the ratio of GCA to SCA, were determined following Griffing,`s (1956) first methods. Analysis of variation was conducted using MSTAT-C and diallel soft wares. Additive variation of degree of dominance was calculated using the following equation.
σ2A = [4/(1+F)] σ2gca |
| σ2D = [4/(1+F)] σ2sca |
where, F was consaquinity σ2 GCA and σ2 SCA were the variance of (GCA) and (SCA), respectively. Since the F2 generation was used, consequently of F was considered 0.5 in the present study. Specific heritability was determined using the following equation:
h2B = (σ2A+ σ2D)/(σ2A+ σ2D+M`e) |
h2N = σ2A/(σ2A+ σ2D+M` e) |
M`e was mean square of error divided to number of replications. Mean of dominance degree was calculated using the following equation:
A = (σ2D/σ2A)0.5 |
RESULTS AND DISCUSSION
Analysis of variances base on Griffing`s 1st method for the all traits showed significant differences (at 0.01 level) between genotypes, mean square of GCA, SCA and reciprocal crosses for the studied traits except the length of the root (Table 1).
The significant differences in GCA and SCA were owing to both additive and non-additive effects in control of these traits. Falconer (1989) suggested that the differences in GCA was due to additive effects and genetic variance as well as alleles interactions and differences in SCA was related to non-additive effect and predominance variance, epistasis and their interaction. The results of analysis of variation for GCA and SCA as per Griffing`s 1st (1956) method for each traits showed that there were significant differences between the ration of GCA/SCA mean square for the traits of days to germination, length of radicle, shootlet, seedling vigor, days to emergence, days to 3 leaf stage, days to 4 leaf stage, days of 3 to 4 leaf stage. It was therefore concluded that both additive and non-additive control the genes of these traits, however non-additive was more important than additive. Degrees of dominance more than 1 in each studied traits and low narrow heritability either in laboratory or green house condition showed the non-additive action of the studied genes (Table 2).
The maximum narrow sense heritability appeared at the duration of 3 to 4 leaf trait (0.31) whereas the minimum was for duration of emergence (0.14). The maximum general heritability was related to duration of germination (0.58) (Table 2).
The highest value of general heritability, in spite of seedling vigor index which has intermediate GCA showed the importance of genetic variation comparing to the environmental variation. It was in harmony with the relative minimum value of cv (coefficient of variation). A portion of genetic variation was related to interaction of environment and genotype seemingly because of controlled situation of the experiments. The degrees of dominance more than 1 showed that the gene was under the control of predominance action. This predominance action might be as a result of imbalanced gamete stage due to gene linked loci. The observations confirmed the findings of Udall et al. (2006) for rapeseed; Sharma et al. (1992) for Indian mustard, Thakara et al. (1994) for Ethiopian mustard and Shamsuddin (2004) for wheat though in their report the narrow sense heritability was more than that of present study. However, Sharma et al. (1992) reported that effects of these genes in Indian mustard was non-additive and the heritability was low.
| Table 1: | Mean square of Diallel analysis following Griffing,s 1st method |
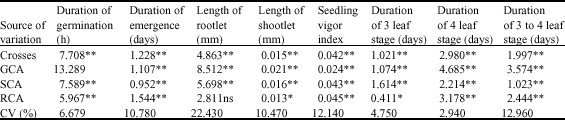 | |
| *,** are significant at 5 and 1 levels, respectively; ns: Not significant, GCA: General Combining Ability, SGA: Specific Combining Ability, RCA: Relative Combining Ability, CV: Coefficient of Variation | |
| Table 2: | Genetic parameters for different traits by diallel crosses of canola |
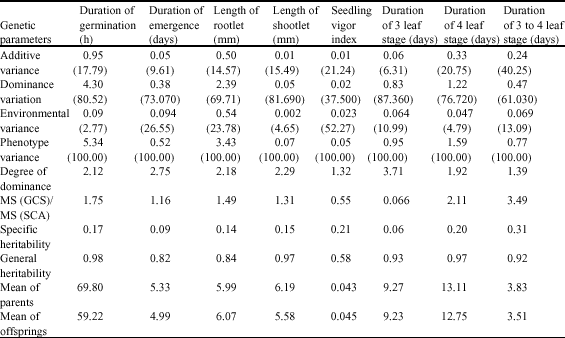 | |
| Values in parenthesis are shown (%) | |
| Table 3: | The combining ability of parents for different traits |
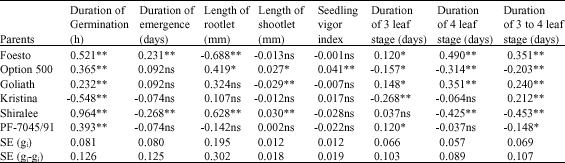 | |
| *,** are significant at 5 and 1 levels, respectively; ns: Non significant | |
The parents of Kristina and Shiralee whose GCA in negative direction were significant can be distinguished as the best parents for decreasing the duration of germination, As decrease in the duration of germination can be defined as an ideal trait (Table 3). The crosses of Kristinax PF-7045/91, Kristina xShiralee, FoestoxGoliath, GoliathxPF-7045/91, FoestoxOption 500, have been chosen as the best crosses because of their significance SCA in the negative direction (Table 4).
In order to have a successive competition with weeds, winter canola should have healthy growth during the seedling stage to maintain vigorous and healthy stand during cool winter, therefore in the four traits of duration of germination as duration of emergence, 3 leaf, 4 leaf and 3 to 4 leaf, only those parents and their offspring are considered as suitable having the shorter period. In the other word the effects of their GCA and SCA would be significant in the negative direction. According to diallel analysis following Griffit,s first method Shiralee was the best parent for the duration of germination, while Foesto parent showed a reverse behavior and resulted in maximum duration of germination (Table 3). Among the studied parents Kristina, Shiralee and Option 500 were figured out the best in respect of the traits of duration of 3 leaf, duration of 4 leaf and duration of 3 to 4 leaf, respectively (Table 3).
| Table 4: | Specific combining ability in different traits |
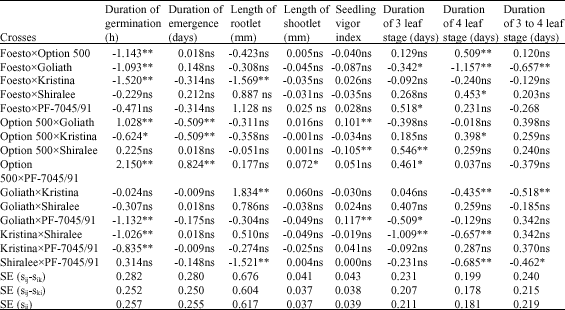 | |
| *,** are significant at 5 and 1 level, respectively; ns: Non significant | |
The crosses of Option 500xGoliath and Option 500xKristina were suitable for the traits of duration of 3 leaf. The crosses of KristinaxShiralee, ShiraleexP.F-7045/91, FoestoxGoliath and GoliathxKristina were proper for the duration of 4 leaf and the crosses of FoestoxGoliath and GoliathxKristina have been figured out to be the best also for the trait the duration of 3 to 4 leaf (Table 4). The parents of Shiralee and Option 500, among the parents, were found to be the best in respect of the length of rootlet and the parents of Shiralee and Option 500 for the length of shootlet and vigor of seedling, respectively (Table 3).
The results also showed that the crosses GoliathxP.F-7045/91 and GoliathxKristina, were the best, among the crosses, for the traits of seedling vigor and length of rootlet, respectively (Table 4). These parents therefore make stable and vigorous rosette before winter cool season.
Mean comparison of the treatments following Duncan`s Multiple Range Test (DMRT) showed that for the trait duration of emergence the best crosses were ShiraleexPF-7045/91, Option 500x Goliath, Option 500xKristina, FoestoxKristina, KristinaxP.F-7045/91 and for the trait 3 leaf the crosses of KristinaxShiralee, Option 500xKristina, ShiraleexP.F-7045/91 and for the trait duration of 3 to 4 leaf the crosses of Option 500xP.F-7045/91, ShiraleexOption 500, ShiraleexPF-7045/91 showed the most encouraging results.
The parents of Shiralee have been identified as the best for the trait of the length of shootlet. The crosses P.F-7045/95xOption 500 and GoliathxShiralee, among the crosses, have been found to be the best for the seedling vigor and the length of rootlet, respectively. Overall it can be concluded that the parent of Shiralee and the cross of KristinaxShiralee were the best for the traits related to the seedling. The numerous studies showed that the parents with the highest mean are the best general combinators and those with the lowest mean are the poorest general combinator (Farshadfar, 1998; Dezhestan et al., 2004). The varieties with high level of general combine ability are suitable for making composite varieties and selection the proper cultivars.
The reverse effects were significant for all the traits except the trait of the length of root (Table 1). It means that there are differences between main crosses and reverse effects. These differences can be an account of cytoplasmic effects. As these effects were found almost in all the traits, the accuracy of these effects was confirmed. Therefore in order to control the traits related to seedling of canola the maternal effects should be taken into consideration. Heterosis effects were evaluated comparing to the better parent (Table 5). There were significant effects of heterosis in all the traits at levels 0.05 and 0.01. The Occurrence of heterosis is as a result of non-additive variance which was evidence of non-additive gene in all the traits. The relatively among of heterosis in F1 and F2 may be because of maternal effect, which makes more complicated the heterosis (Falconer, 1989).
| Table 5: | Heterosis base on the Better parent in different traits |
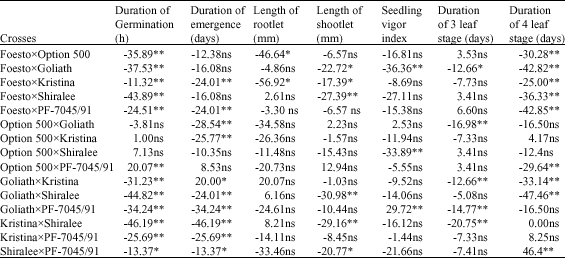 | |
| *,** are significant at 5 and 1 level, respectively; ns: Non significant | |
If some gene locations are dominant in specific direction and some other in other direction, these effects neutralize each other. Therefore despite of dominance for all gene locations no heterosis is found. Heterosis occurrence therefore depends on directional dominance when there is no heterosis it means that the single-gene locations do not show dominance. Lack of the seed in F1 generation causes difficulty in the evaluation of heterosis, this problem can be solve by evaluation of F2 (Engqvist and Becker, 1991) and or other methods descript by Dayteg et al. (2007). Significant variation of the traits of study (Table 1) showed the genetically diversion among the cultivars. Significant differences in GCA and SCA is a proof not only for the effects of importance of parents` combine ability and their hybrids, but also is a proof of the additive and non-additive effects of controller genes for these traits. The traits having the minimum heritability are considered as reproductive reputation (Weber et al., 2005).
REFERENCES
- Brown, J. and P. Caligari, 2008. An Introduction to Plant Breeding. 1st Edn., Wiley-Blackwell, Oxford, UK., ISBN : 978-1-4051-3344-9 pp:66-115.
Direct Link - Dezhestan, S., M. Moghaddam, M.R. Shakiba, H. Alliari and F. Javidfar, 2004. Genetically analysis of cold resistance in canola via diallel. Iran. J. Agric. Sci., 14: 105-121.
Direct Link - Dayteg, C. and S. Tuvesson, A. Merker, A. Jahoor and A. Kolodinska-Brantestam, 2007. Automation of DNA marker analysis for molecular breeding in crops: Practical experience of a plant breeding company. Plant Breed., 126: 410-415.
CrossRef - Engqvist, G.M. and H.C. Becker, 1991. Heterisis and epistasis in rapeseed estimated from generation mean. Euphytica, 85: 21-25.
Direct Link - Falconer, D.S., 1989. Introduction to Quantitative Genetics. 3rd Edn., Longman Scientific and Technical, New York, USA Pages: 438.
Direct Link - Griffing, B., 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci., 9: 463-493.
Direct Link - Hayman, B.I., 1960. The theory and analysis of diallel crosses. III. Genetics, 45: 155-172.
Direct Link - Latifi, N., A. Soltani and V.D. Espanser, 2004. Effects of temperature on the germination parameters of rapeseed cultivars. Iran. J. Agric. Sci. 35: 313-321.
Direct Link - Shamsuddin, A.K.M., 1985. Genetic diversity in relation to heterosis and combining ability in spring wheat. Theoret. Applied Genet., 70: 306-308.
CrossRefDirect Link - Taleie, E. and A.H. Beigi, 1996. Determination of combine ability and Heterosis in wheat varieties via diallel hybridation. Iran. J. Agric. Sci., 27: 67-74.
Direct Link - Thiagarajah, M.R. and G.R. Stringam, 1993. A comparison of genetic segregation in traditional and microspore-derived populations of Brassica juncea L. czern and coss. Plant Breed., 111: 330-334.
CrossRef - Udall, J.A., P. Quijada, B. Lambert and T. Osborn, 2006. Quantitative trait analysis of seed yield and other complex traits in hybrid spring rapeseed (Brassica napus L.): 2. Identification of alleles from unadapted germplasm. TAG Theoretical Applied Genet., 113: 597-609.
CrossRef - Weber, S., F. Ünker and W. Friedt, 2005. Improved doubled haploid production protocol for Brassica napus using microspore colchicine treatment in vitro and ploidy determination by flow cytometry. Plant Breed., 124: 511-513.
CrossRef - Yasari, E. and A.M. Patwardhan, 2007. Effects of (Azotobacter and Azosprillium) inoculants and chemical fertilizers on growth and productivity of canola (Brassica napus L.). Asian J. Plant Sci., 6: 77-82.
CrossRefDirect Link - Yasari, E., A.M. Patwardhan, V.S. Ghole, G.C. Omid and A. Ahmad, 2008. Relationship of growth parameters and nutrients uptake with canola (Brassica napus L.) yield and yield contribution at different nutrients availability. Pak. J. Biol. Sci., 11: 845-853.
CrossRefPubMedDirect Link