
Kangtao
Shanxi Provincial People�s Hospital, No. 256, Youyi West Road, Xi�an, 710068 Shanxi, People�s Republic of China
LiveDNA: 86.23837
Yangqian
Shanxi Provincial People�s Hospital, No. 256, Youyi West Road, Xi�an, 710068 Shanxi, People�s Republic of China
Souravh Bais
Department of Pharmacology, Rayat Institute of Pharmacy, Railmajra, SBS Nagar District, 144506 Punjab, India
LiveDNA: 91.5733
International Journal of Pharmacology
Year: 2018 | Volume: 14 | Issue: 6 | Page No.: 879-888







ABSTRACT
Background and Objective: Recent studies have revealed that monoamine oxidase-B (MAO-B) inhibitors effective in treating Alzheimer’s disease (AD), with a promotable extension of life span. It modulates nitric oxide (NO), which contribute to cognitive function in AD. The present study investigated the potential of protocatechuic acid (PCA) as MAO-B inhibitor and its effect on release of MAO, TNF-α, acetylcholine esterase enzyme, in cognitive dysfunctions associated with experimental dementia in rats. Materials and Methods: Aluminium chloride (AlCl3) was administered at a dose of 175 mg kg–1 per oral (p.o) for a period of 25 days in rats and then divided into different groups, i.e. standard group, negative control and two groups of PCA, (at a dose of 10 and 20 mg kg–1, p.o.), where these groups treated and observed until the 35th day of experimental trial. Morris water mazes (MWM), photoactometer test were performed on 5th, 16th, 26th and 36th day to access learning, memory and ambulatory movements. Later, the animals were sacrificed for biochemical and histopathological studies. Extent of oxidative stress was measured by estimating the levels of Glutathion (GSH), superoxide dismutase (SOD), nitrite, catalase. Brain acetylcholine esterase activity and MAO-A, MAO-B were also estimated. The brain levels of TNF-α was measured as marker of inflammation. Results: AlCl3 produced a marked decline in MWM performance and ambulatory movements’ of animals, reflecting impairment of memory and learning. PCA treatment significantly modulates AlCl3 induced memory deficits, biochemical and pathological alterations. Conclusion: The findings demonstrated that the memory restorative ability of PCA may be attributed due to its anti-cholinesterase, anti-oxidative and anti-inflammatory potential.
PDF Abstract XML References Citation
Received: February 19, 2018;
Accepted: March 30, 2018;
Published: July 15, 2018
Copyright: © 2018. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Kangtao, Yangqian and Souravh Bais, 2018. Neuroprotective Effect of Protocatechuic Acid Through MAO-B Inhibition in Aluminium Chloride Induced Dementia of Alzheimer’s Type in Rats. International Journal of Pharmacology, 14: 879-888.
DOI: 10.3923/ijp.2018.879.888
URL: https://scialert.net/abstract/?doi=ijp.2018.879.888
DOI: 10.3923/ijp.2018.879.888
URL: https://scialert.net/abstract/?doi=ijp.2018.879.888
INTRODUCTION
Alzheimer’s disease (AD) is a progressive and irreversible neurodegenerative disease, leads to death of neuronal cells. This manifested by deterioration in memory and cognition, impairment in performing activities of daily living and many behavioral neuropsychiatric illnesses and consequences are marked impairment of memory and abstract reasoning, with preservation of vision and movement1.
MAO-B inhibitors might offer an alternative for AD therapy as they have a long history of clinical use and are considered safe and nontoxic2. This article evaluates the current role of PCA as MAO-B inhibitor in the treatment of AD and explores the concept that actions other than antioxidants, MAO inhibition and TNF suppression may also contribute to therapeutic efficacy of the drug. MAO exists in two forms, MAO-A and MAO-B, coded by two distinct gene loci3, with different patterns of tissue distribution. MAO-A pre-ferentially deaminates serotonin (5-HT) and is more sensitive to inhibition by clorgyline, whereas, MAO-B deaminates pre-ferentially b-phenylethylamine and is inhibited by drugs such as L-deprenyl. The optical isomer D-deprenyl is biologically less active than L-deprenyl. Clinically the most important monoamine that is elevated by L-deprenyl is dopamine, which is a substrate for both MAO-A and MAO-B. L-Deprenyl (selegiline) has been used as an effective adjuvant to L-DOPA in the treatment of Parkinson’s disease. It has also been reported that L-deprenyl by itself can delay the onset of disability associated with early untreated Parkinson’s disease. L-Deprenyl also prevents MPTP-induced neurotoxicity and Parkinson-like symptoms in animals2,4, so the compound with selective inhibition or non selective approach may be the symptomatic cure in AD and Parkinson’s disease5. The more reactive oxygen species (ROS) will lead to imbalance between the formation of cellular oxidants and the anti-oxidative processes. Oxidative metabolism of acetylcholine plays an important role in AD pathogenesis6,7. PCA is traditionally reported as a significant antioxidant and anti inflammatory property, so the aim of present study is focused on modulation of various neurochemicals by PCA in brain tissues. PCA is reported for different pharmacological activities like antioxidant and free-radical scavenging property, cardioprotective, antihypertensive and cardiovascular remodeling8,9, anti-hyperglycemic action10, anti-cancer activity11, anti-ulcer activity12, anti-aging13, anti-inflammatory and analgesic14, anti-fibrotic effects15, anti-viral activity16 nephro-protective and hypolipidemic activity17-22 (Fig. 1). However the diverse pharmacology of this compound is still to be unexplored. The present study investigated the potential of PCA as monoamine oxidase (MAO-B) inhibitor, in cognitive dysfunctions associated with experimental dementia.
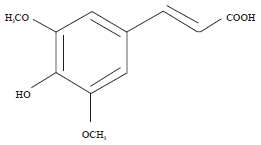 | |
| Fig. 1: | Structure of PCA |
MATERIALS AND METHODS
This study was conducted jointly by in RBGI, India and Shanxi Provincial People’s Hospital.
Drugs and chemicals: PCA (Pubchem CID 637775) was purchased from Sigma Aldrich (S. and G. Lab Supplies). selegiline was procured as a gift sample from Sun pharmaceutical Pvt. Ltd. Baddi (H.P.) India. Chemicals like aluminium chloride, DTNB, acetylthiocholine iodide, trichloroacetic acid, thiobarbituric acid, sodium carboxy methyl cellulose was procured from Himedia Pvt. Ltd. (Mumbai) S.D. Fine Chemicals Ltd. (Mumbai). Solvents like methanol, chloroform, dichloromethane, tween 80, n-butanol and ethyl acetate were of analytical grade (AR).
Animals: Age matched young wistar albino rats of either sex, weighing 120-150 g were selected as the study. The animals were kept in the paddy husk as bedding material. Husk changed every day. The animals were housed in a group of 6 per polypropylene cages kept under controlled room temperature (25±1°C) in 12 h light-dark cycle. The rats were allowed free access to food (Standard pallet) and water. The experiment was conducted in a noise-free environment between 9:00 AM to 2:00 PM. All procedures were approved and carried out as per the guidelines of animal experimentation (IAEC/PO/Re/S/1/5).
Acute toxicity study (LD50): LD50 was determined according to the guidelines of organization for economic cooperation and development (OECD) following the up and down method (OECD guideline No. 423) and fixed dose method (OECD guideline No. 420). Based on this guideline a limit test was to categorize the toxicity class (LD50) of the compound. The limit test was performed at 2000 mg kg–1, p.o.
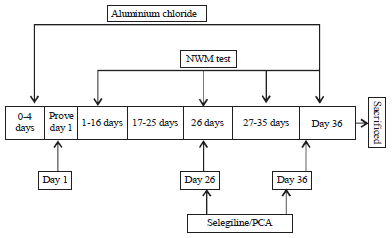 | |
| Fig. 2: | Experimental protocol for aluminium chloride induced dementia |
| Table 1: | Sequence of training trials on Morris water maze |
 | |
A dose range of 10, 20 mg kg–1 was selected for the pharmacological activity. For all the studies overnight fasted animals were used.
Experimental design: On prove day (day 5-consider as day 1), randomized animals were divided randomly into experimental groups (n = 8) (overall protocol-(5+36 = 42 days)). Control group received a normal saline (5 mL kg–1, p.o). AlCl3, selegiline and PCA suspensions were made freshly at a time of dosing. Morris water mazes (MWM), photoactometer test were performed on 5th, 16th, 26th and 36th day to access learning, memory and ambulatory movements. Later, the animals were sacrificed for biochemical and histopathological studies. Extent of oxidative stress was measured by estimating the levels of glutathion (GSH), superoxide dismutase (SOD), nitrite, catalase. Brain acetylcholine esterase (Ache) activity and monoamine oxidase (MAO-A, MAO-B) were also estimated. The brain levels of TNF-α was measured as marker of inflammation (Fig. 2, Table 1).
Group I: Control group (normal saline (0.9% NaCl)-5 mL kg–1, p.o., from 6th-36th day); rats were administered distilled water (10 mL kg–1, p.o.) 30 min before acquisition trial was conducted from day 1-4 and 30 min before retrieval trail was conducted on day 5.
Group II: Untreated AlCl3-affected rats; rats were administered with AlCl3 suspension (175 mg kg–1) orally from day 0 (i.e., 24 h after the completion of retention trial on day 5) to 36 days.
Group III: Selegiline-treated AlCl3-affected rats; after 25 days, rats were administered standard drug suspensions (selegiline-2.5 mg kg–1), (1% aqueous solution of Tween 80) orally from day 26-36.
Group IV and V: PCA-Treated AlCl3-affected rats; after 25 days, rats were administered PCA suspension (at two doses 10 mg and 20 mg kg–1 to respective group) (1% aqueous solution of Tween 80) orally from day 26-36.
Memory assessment (water maze test): The water maze consisted of a circular tank (150 cm diameter and 40 cm height). Water pool was divided into four equally spaced quadrants North-East (NE), South-East (SE), South-West (SW) and North-West (NW) along the circumference of the pool. An escape platform (10 cm diameter) submerged 2cm below the water surface and was placed in NW quadrant. Rats were trained to locate the hidden platform at a fixed location in NW quadrant. All rats were subjected to one session of four trials per day for five consecutive days (0-5th day). During each trial, the animal was placed in each quadrant to eliminate quadrant effects. All rats were left in the platform for 30 sec and then removed and towel dried. Rats failing to find the platform within 60 sec were guided to the platform (Table 1). In day 5 (Probe day/zero day), 24 h after previous training, escape platform was removed and probe trial was conducted. The cut-off time for animal to swim was set to 60 sec before the end to the session. Similarly, the retention trials were conducted on day 5th 16th, 26th and day 36th on different groups to evaluate memory. Time elapsed in escaping to the NW quadrant, i.e. escape latency time (ELT) and total time (TT) time spent in NW quadrant, was measured during the retention trials20 (Fig. 3).
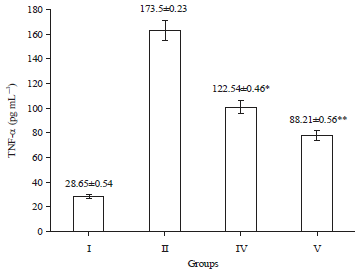 | |
| Fig. 3: | Effect of PCA on TNF-α level in brain cortex. All values were represented as Mean±SEM. Data are Mean±SEM values, n = 8. Data were analyzed by one-way ANOVA followed by Tukey’s-Kramer’s multiple comparisons test. *p<0.05, **p<0.01, Where I: Control, II: Untreated AlCl3-affected rats, IV: Test A-PCA (10 mg kg–1, p.o.)-treated AlCl3-affected rats and V: Test B-PCA (20 mg kg–1, p.o.)-treated AlCl3-affected rats |
Photoactometer: Animals were assessed for locomotor activity using digital photoactometer. The ambulatory movements were recorded over a period of 10 min and expressed in terms of total photo beam counts for 10 min per animal. Locomotor activity (L.A.) was assessed on day 0, 5th, 11th, 26th and day 36th through Morris’s water maze20.
Evaluation
Brain tissue sampling and preparation: After 24 h of the experimental period (after 35 days), the animals were sacrificed and their brains were removed and weighed. The whole brain was washed thoroughly with ice-cold isotonic saline. A 10% issue homogenate was prepared in 0.1 M phosphate buffer (pH 8, stored 2-8°C) for various neurochemical estimations and other anti oxidative parameters.
Biochemical assessment
Acetyl choline esterase (AchE) activity: This activity was measured by Ellman et al.21 method.
Estimation of glutathione (GSH): GSH level was measured by the method described by Ellman et al.21 with slight modifications. The absorbance was measured spectrophoto metrically at 412 nm (X-Rite 640B spectrophotometer). Different concentration of GSH standard was also processed similarly to prepare a standard curve (1-50 μg) simultaneously. Results were expressed as nmol of GSH mg–1 of protein.
Estimation of catalase: This activity was determined by Armstrong et al.22 method.
Estimation of superoxide dismutase (SOD): SOD enzyme activity was measured by the method described by Ohkawa et al.23 and Yagi24.
Estimation of TNF-α: The TNF-α level was estimated by ELISA kit.
Estimation of MAO-A and MAO-B: Estimation of MAO activity; it was assayed by the method described by Charles and McEwen25.
Estimation of nitrite: It was measured by Nazmun method26.
Histopathological examination: The second portion of each brain was fixed in formalin buffer (10%) for 24 h. The brains were washed in tap water and then dehydrated using serial dilutions of alcohol (methyl, ethyl and absolute ethyl). Specimens were cleared in xylene and embedded in paraffin in a hot air oven at 56°C for 24 h. Paraffin bees wax blocks were prepared for sectioning at 4 μm using a microtome. The obtained tissue sections were collected on glass slides, deparaffinized and stained with hematoxylin and eosin stains27 for histopathological examination using a light microscope.
Statistical analysis: The data were expressed as Mean±SEM. In all the tests, the criterion for the statistical significance was set at p<0.05. The data for all studies were analyzed using one-way ANOVA followed by Tukey’s-Kramer’s multiple comparisons test.
RESULTS
Acute toxicity study: PCA did not show any sign and symptom of toxicity and mortality up to 2000 mg kg–1, p.o.
Effect of PCA on Aluminium chloride induced behavioural parameters
Effect on ELT and time spent in target quadrant using the Morris water maze: The rats spent more time in the target quadrant (NW) during retention trial on Probe day (Day 5).
| Table 2: | Effect of PCA on aluminium chloride induced dementia of Alzheimer’s type in rat using Morris water maze (escape latency time) |
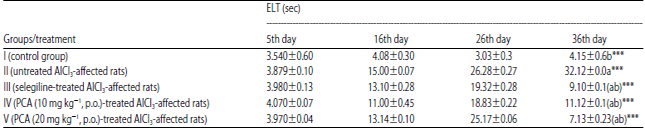 | |
Data are Mean±SEM values, n = 8. Data were analyzed by one-way ANOVA followed by Tukey’s-Kramer’s multiple comparisons test. ***p<0.001. a: Compared with control, b: Compared with inducer, ELT: Escape latency time | |
| Table 3: | Effect of PCA on aluminium chloride induced dementia of Alzheimer’s type in rat using Morris water maze |
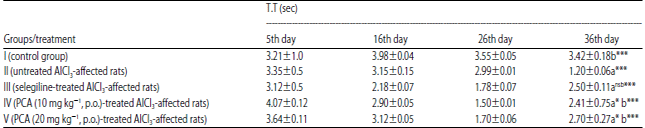 | |
Data are Mean±SEM values, n = 8. Data were analyzed by one-way ANOVA followed by Tukey’s-Kramer’s multiple comparisons test. *p<0.05, ***p<0.001 and nsp>0.05. a: Compared with control, b: Compared with inducer, nsNot significant, T.T: Total time | |
| Table 4: | Effect of PCA on aluminium chloride induced dementia of Alzheimer’s type in rat using photoactometer |
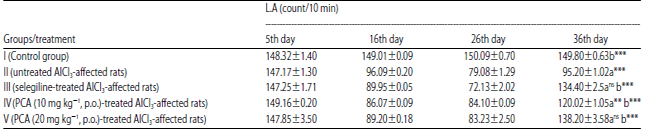 | |
Data are Mean±SEM values, n = 8. Data were analyzed by one-way ANOVA followed by Tukey’s-Kramer’s multiple comparisons test. **p<0.01, ***p<0.001 and nsp>0.05. a: Compared with control, b: Compared with inducer, nsNot significant, LA: Loco motor activity | |
Control group showed the normal retrieval of memory on 36th day. AlCl3 treated rats (Group II) significantly raised ELT and reduced, time spent in the target quadrant (NW). Untreated AlCl3-affected rats (Group III) significantly raised the ELT and reduced the TT during the retention trial on 16th and 26th day, but the selegiline-treated AlCl3-affected group resulted, a decrease in ELT and increased in TT during retention trial on 36th day. PCA-treated AlCl3-affected (10 and 20 mg kg–1) rats showed, decrease in ELT and increase in TT during retention trial on 36th day Table 2 and 3.
Effect on locomotor activity using photoactometer: Locomotor activities (ambulatory movements) of rats were recorded for a period of 10 min and expressed in terms of total photo beam counts for 10 min/animal. Control group rats (Group I) showed the normal locomotor activity on 36th day (149.8±0.63 counts/10 min). Aluminium chloride treated rats (Group II) resulted in a significant decreased in locomotor activity on 16th, 26th and 36th day (p<0.001) as compared to control group rats (Group, I). Untreated AlCl3-affected rats (Group III) resulted in a significant decreased in locomotor activity on 16th and 26th day, but after selegiline treatment results in a significant increased in locomotor activity on 36th day (p<0.001) when compared to AlCl3 treated rats (Group II). PCA-treated AlCl3-affected (10/20 mg kg–1) resulted, in a significant increase in locomotor activity (p<0.001) when compared to AlCl3 treated rats (Group II) Table 4.
Effect on biochemical parameters: The levels of AchE, TBARS and nitrite were found elevated in Untreated AlCl3-affected rats as compared to control group. The animals treated with selegiline showed a significant reduction in the levels of AchE, TBARS and nitrite on 36th day of trial as compared with untreated AlCl3-affected group. The elevated levels of AchE, TBARS and nitrite were declined on treatment with PCA in a dose-dependent manner, as compared with untreated AlCl3-affected group. The levels GSH, catalase and SOD were significantly decreased in untreated AlCl3-affected rats as compared to a control group.
| Table 5: | Effect of PCA on various biochemical parameters in rats’ brain |
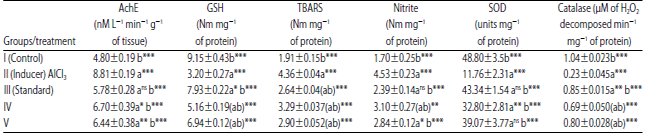 | |
Group I: Control Group, Group II: Untreated AlCl3-affected rats, Group III: Selegiline-treated AlCl3-affected rats, Group IV: PCA (10 mg kg–1, p.o.)-treated AlCl3-affected rats, Group V: 20 mg kg–1, p.o.-treated AlCl3-affected rats. Data are Mean±SEM values, n = 8. Data were analyzed by one-way ANOVA followed by Tukey’s-Kramer’s multiple comparisons test. *p<0.05, **p<0.01, ***p<0.001 as compared with inducer, nsNot significant | |
| Table 6: | Effect of PCA on AlCl3 induced toxicity with reference to change in body and brain weight |
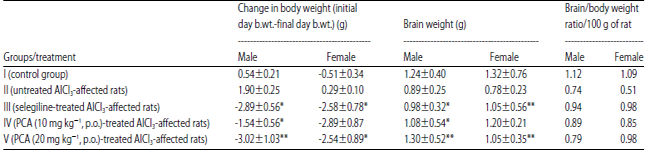 | |
Data are Mean±SEM values, n = 8. (male-4, Female-4). Data were analyzed by one-way ANOVA followed by Tukey’s-Kramer’s multiple comparisons test. *p<0.05, **p<0.01, as compared with inducer | |
| Table 7: | Effect of PCA on MAO level in brain tissues |
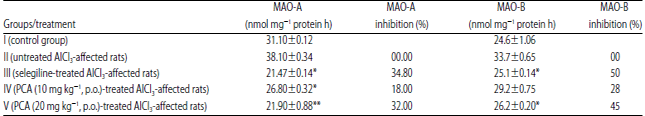 | |
Data are Mean±SEM values, n = 8. Data were analyzed by one-way ANOVA followed by Tukey’s-Kramer’s multiple comparisons test. *p<0.05, **p<0.01, as compared with inducer | |
The animals treated with selegiline showed a significant increase in the levels of GSH, catalase and SOD on 36th day of trial as compared with untreated AlCl3-affected group. The reduced levels of GSH, catalase and SOD were increased significantly with a dose of 20 mg kg–1 of PCA as compared with untreated AlCl3-affected group (Table 5).
Effect on brain TNF-α level: All AlCl3 treated rats showed an increase in TNF-α level (Fig. 3) in the brain. PCA treated rats showed a significant reduction in TNF-α levels as compared to control.
Effect on body weight and brain’s weight: Untreated AlCl3-affected rats showed a decrease in body weight and brain weight while other groups did not show any marked deference in body and brain weight of rats (Table 6).
Effect of PCA on MAO-A and MAO-B levels: The effects of PCA on MAO concentration were shown in Table 7. The MAO-A and MAO-B activities in a control group were 31.1±0.12 and 24.6±1.06 nmol mg–1 of protein, respectively. PCA at a dose of 10 mg kg–1, p.o. showed 18% (MAO-A) and 28% (MAO-B) inhibition. PCA at a dose of 20 mg kg–1, p.o. showed a marked inhibition in both type, MAO-A (32%) and MAO-B (45%).
Histopathological changes in brain: As shown in Fig. 4, the control animals (Fig. 4a), had neuronal cells were visualised normally under microscope; cells are active and relatively packed with prominent nuclei. AlCl3 treated rats showed (Fig. 4b) necrosis of cells and more neuronal vacuolation. The selegiline treated rats (Fig. 4c), showed a mild necrosis and deformed nuclei. The PCA treated rats (10 mg kg–1, p.o) showed slight neuronal decongestion and the rats treated with high dose of PCA (20 mg kg–1, p.o) showed, no cell decongestion cell deformity, Test A (Fig. 4d).
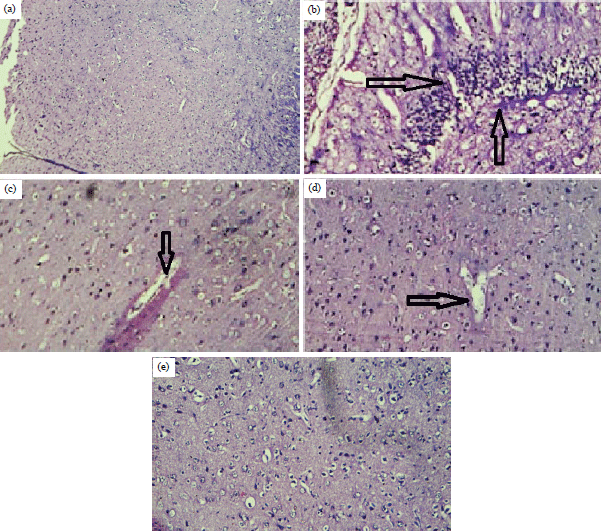 | |
| Fig. 4(a-e): | Histopathology of brain (cerebral cortex) showing neuronal degeneration and inflammation. a: Control, (group I), b: Negative (group II), c: Standard (group III), d: Test A (group IV) and e: Test B (group V) |
DISCUSSION
Administration of chronic AlCl3 causes severe memory deficits in rats as predicted by this study. Followed by PCA treatment, showed an improvement in memory deficits. Aluminium as metal ions commonly present in drinking water, soil and tooth paste, moreover, it is used to manufacture cooking utensils28. Literature reports showed that its chronic use leads to oxidative deterioration of cellular lipids, proteins and DNA29. Lipid peroxidation is one, can cause a tissue damage under its chronic use30,31 therefore, aluminium can be considered as a contributing factor in AD. After chronic administration, it accumulates in all brain regions with greater quantity in cortex and hippocampus32,33 and damaged it. These regions play an important role in learning and memory34.
In this study, a significant oxidative stress in aluminium treated rats was observed, as indicated by an increase in lipid peroxidation, nitrite levels, decrease in reduced glutathione levels and catalase and superoxide dismutase activity. This could be due to the reduced axonal mitochondria turnover, disruption of Golgi and reduction of synaptic vesicles induced by aluminium treatment and release of oxidative products like malondialdehyde TBARS, carbonyls, peroxynitrites and enzymes like superoxide dismutase within the neurons35. Moreover the groups treated with PCA showed significant antioxidant activity8 against the neuronal damage caused by aluminium.
Impaired cholinergic transmission is one of the complications seen in the etiopathogenesis of memory deficit in AD. The neurodegeneration in frontal cortex and hippocampus areas within the brain36 resulting in impaired cholinergic transmission by two ways. Firstly, (i) decline in Ach release, (ii) decreased choline acetyltransferase activity (ChAT), which results in the scarcity of acetylcholine37,38. Secondly, elevated acetyl cholinesterase (AchE) enzyme further adds to scarcity of Ach at the synapse by degrading the available Ach36.
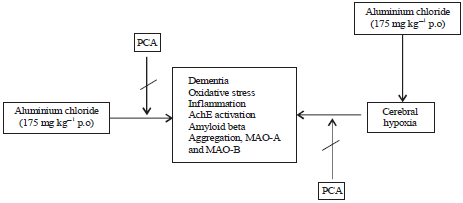 | |
| Fig. 5: | Proposed mechanism for therapeutic effect of PCA |
This degradation of Ach is abolished by PCA (AchE inhibitor) so it’s effective in AD through improvement in cholinergic transmission.
PCA (hydroxycinnamic acid) belongs to phenolic acids with bioactive carboxylic acids; this class mainly includes caffeic acid, ferulic acid and PCA39,40. It is a frequent phytochemical in the human diet12. It’s a potent inhibitor of an enzyme AchE41,42. This could be a possible mechanism by it inhibits cerebral hypoxia and improved memory disturbance by activating cholinergic function acetylcholine (Ach) and choline acetyltransferase (ChAT)43.
Chronic aluminium administration has been reported to result in behavioural changes such as cognitive44 and locomotor impairment45. This study also showed an impaired behaviour of rats treated with aluminium. The cognitive deficit is evident to declined performance in Morris water maze test46 and radial arm maze test47. In current study, aluminium treatment resulted in behavioral changes such as a spatial memory deficit. Selegiline and PCA antagonized the spatial memory deficit caused by aluminium. This suggested that the neuro protective role of PCA in correcting cognitive dysfunction was associated with aluminium exposure.
Assessment of locomotor activity is a requirement for evaluating any possible CNS depressant/stimulant effect of interventions on animals. Similar to previous reports43, a decline in locomotor activity in aluminium treated rats was observed, indicating the CNS depressant effect of chronic aluminium exposure. Treatment with Selegiline and PCA corrected the locomotor in-coordination caused by aluminium chloride.
MAO inhibitors are unique in that they exert their protective effect on both vascular and neuronal tissue by increasing nitric oxide (NO) production in brain tissues and cerebral blood vessels. Which is evidentially observed less in AD brain44. So this study also indicated that PCA may play a significant role as MAO inhibitor and increasing NO release and decreasing nitrite in brain tissues.
Histopathological examination of aluminium chloride induced AD brain showed spongy cell, neuronal necrosis. Cells degenerated with small nuclei leading to eosinophilic deposition. However, besides these pathological hallmarks, AD brain exhibited a clear evidence of chronic inflammation and oxidative damage1,44-47. These were also thought to play a significant role at the onset and progression of AD. Present study also supported an evidence of inflammation with high concentration of TNF-α in brain tissues as compared with control rats. Administration of PCA in AD rats improved the pathogenesis of AD as demonstrated by an improvement in the behavioral (levels of activity and motor coordination), Inflammatory (TNF-α) and biochemical parameters in the brains, which was further confirmed by an improvement in brain tissue characteristics on histopathological analysis.
Accumulation of free radicals may cause degenerative events of aging such as AD and because PCA showed its potential against this neurotoxic (AlCl3) agent so it should be used in treatment of dementia or AD (Fig. 5).
CONCLUSION
From the above pharmacological, biochemical and histopathological studies, it has been concluded that PCA showed a significant anti inflammatory activity, antioxidants and neuromodulatory effects against aluminium chloride induced dementia of Alzheimer’s type in rats.
SIGNIFICANCE STATEMENT
This study was designed to evaluate the potential role of PCA in memory deficits produced by AlCl3 and findings of this study suggested the significant MAO inhibition by PCA which creates further interest among scientific researchers to explore its molecular interactions with this enzyme. This study also mentions the therapeutic benefits of PCA as nutraceuticals in the treatment of neurodegenerative disease.
ACKNOWLEDGMENT
Not disclosed
REFERENCES
- Thomas, T., 2000. Monoamine oxidase-B inhibitors in the treatment of Alzheimers disease. Neurobiol. Aging, 21: 343-348.
CrossRefPubMedDirect Link - Heikkila, R.E., A. Hess and R.C. Duvoisin, 1984. Dopaminergic neurotoxicity of 1-Methyl-4-phenyl-1,2,5,6-tetrahydropyridine in mice. Science, 224: 1451-1453.
CrossRefPubMedDirect Link - Shih, J.C., 1991. Molecular basis of human MAO-A and MAO-B. Neuropsychopharmacology, 4: 1-7.
Direct Link - Kimura, R. and M. Ohno, 2009. Impairments in remote memory stabilization precede hippocampal synaptic and cognitive failures in 5XFAD Alzheimer mouse model. Neurobiol. Dis., 33: 229-235.
CrossRefDirect Link - Terry, A.V.Jr. and J.J. Buccafusco, 2003. The cholinergic hypothesis of age and Alzheimer's disease-related cognitive deficits: Recent challenges and their implications for novel drug development. J. Pharmacol. Exp. Ther., 306: 821-827.
CrossRefPubMedDirect Link - Daiello, L.A., E.K. Festa, B.R. Ott and W.C. Heindel, 2008. Cholinesterase inhibitors improve visual attention in drivers with Alzheimer’s disease. Alzheimer's Dementia: J. Alzheimer's Assoc., 4: T498-T498.
CrossRefDirect Link - Liu, Y., G. Fiskum and D. Schubert. 2002. Generation of reactive oxygen species by the mitochondrial electron transport chain. J. Neurochem., 80: 780-787.
CrossRefDirect Link - Sherer, T.B., J.R. Richardson, C.M. Testa, B.B. Seo and A.V. Panov et al., 2007. Mechanism of toxicity of pesticides acting at complex I: Relevance to environmental etiologies of Parkinson’s disease. J. Neurochem., 100: 1469-1479.
CrossRefDirect Link - Kakkar, S. and S. Bais, 2014. A review on protocatechuic acid and its pharmacological potential. ISRN Pharmacol.
CrossRefDirect Link - Ciftci, O., O.M. Disli and N. Timurkaan, 2013. Protective effects of protocatechuic acid on TCDD-induced oxidative and histopathological damage in the heart tissue of rats. Toxicol. Ind. Health, 29: 806-811.
CrossRefDirect Link - Scazzocchio, B., R. Vari, C. Filesi, M. D'Archivio and C. Santangelo et al., 2011. Cyanidin-3-O-β-glucoside and protocatechuic acid exert insulin-like effects by upregulating PPARγ activity in human omental adipocytes. Diabetes, 60: 2234-2244.
CrossRefDirect Link - Tanaka, T., T. Tanaka and M. Tanaka, 2011. Potential cancer chemopreventive activity of protocatechuic acid. J. Exp. Clin. Med., 3: 27-33.
CrossRefDirect Link - Kore, K.J., P.P. Bramhakule, R.M. Rachhadiya and R.V. Shete, 2011. Evaluation of antiulcer activity of protocatechuic acid ethyl ester in rats. Int. J. Pharm. Life Sci., 2: 909-915.
Direct Link - Shi, G.F., L.J. An, B. Jiang, S. Guan and Y.M. Bao, 2006. Alpinia protocatechuic acid protects against oxidative damage in vitro and reduces oxidative stress in vivo. Neurosci. Lett., 403: 206-210.
CrossRefDirect Link - Li, C., W. Jiang, H. Zhu and J. Hou, 2012. Antifibrotic effects of protocatechuic aldehyde on experimental liver fibrosis. Pharm. Biol., 50: 413-419.
CrossRefDirect Link - Zhou, Z., Y. Zhang, X.R. Ding, S.H. Chen and J. Yang et al., 2007. Protocatechuic aldehyde inhibits hepatitis B virus replication both in vitro and in vivo. Antiviral Res., 74: 59-64.
CrossRefDirect Link - Lende, A.B., A.D. Kshirsagar, A.D. Deshpande, M.M. Muley, R.R. Patil, P.A. Bafna and S.R. Naik, 2011. Anti-inflammatory and analgesic activity of protocatechuic acid in rats and mice. Inflammopharmacology, 19: 255-263.
CrossRefDirect Link - Borate, A.R., A.A. Suralkar, S.S. Birje, P.V. Malusare and P.A. Bangale, 2011. Antihyperlipidemic effect of protocatechuic acid in fructose induced hyperlipidemia in rats. Int. J. Pharm. Bio Sci., 2: 456-460.
Direct Link - Lee, J.H., H.J. Lee, W.C. Choi, S.W. Yoon and S.G. Ko et al., 2009. Rhus verniciflua stokes prevents cisplatin-induced cytotoxicity and reactive oxygen species production in mdck-i renal cells and intact mice. Phytomedicine, 16: 188-197.
CrossRefDirect Link - Bais, S., R. Kumari and Y. Prashar, 2017. Therapeutic effect of Sinapic acid in aluminium chloride induced dementia of Alzheimer's type in rats. J. Acute Dis., 6: 154-162.
CrossRefDirect Link - Ellman, G.L., K.D. Courtney, V. Andres Jr. and R.M. Featherstone, 1961. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol., 7: 88-90.
CrossRefDirect Link - Armstrong, D.M., M.D. Ikonomovic, R. Sheffield and R.J. Wenthold, 1994. AMPA-selective glutamate receptor subtype immunoreactivity in the entorhinal cortex of non-demented elderly and patients with Alzheimer's disease. Brain Res., 639: 207-216.
CrossRefDirect Link - Ohkawa, H., N. Ohishi and K. Yagi, 1979. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem., 95: 351-358.
CrossRefPubMedDirect Link - Yagi, K., 1998. Simple procedure for specific assay of lipid hydroperoxides in serum or plasma. Free. Radic. Antioxid. Protocol., 108: 107-110.
CrossRefDirect Link - Lyle, N., D. Bhattacharyya, T.K. Sur, S. Munsi, S. Paul, S. Chatterjee and A. Gomes, 2009. Stress modulating antioxidant effect of Nardostachys jatamansi. Indian J. Biochem. Biophys., 46: 93-98.
Direct Link - Bancroft, J., A. Stevens and D. Turner, 1996. Theory and Practice of Histological Techniques. 4th Edn., Churchill, Livingston, New York, London, San Francisco, Tokyo Pages: 740.
CrossRefDirect Link - Hansson, O., H. Zetterberg, P. Buchhave, E. Londos, K. Blennow and L. Minthon, 2006. Association between CSF biomarkers and incipient Alzheimer's disease in patients with mild cognitive impairment: A follow-up study. Lancet Neurol., 5: 228-234.
CrossRefDirect Link - Maccioni, R.B., J.P. Munoz and L. Barbeito, 2001. The molecular bases of Alzheimer's disease and other neurodegenerative disorders. Arch. Med. Res., 32: 367-381.
CrossRefPubMedDirect Link - Bjertness, E., J.M. Candy, A. Torvik, P. Ince and F. McArthur et al., 1996. Content of brain aluminum is not elevated in Alzheimer disease. Alzheimer Dis. Assoc. Disord., 10: 171-174.
Direct Link - Sargazi, M., A. Shenkin and N.B. Roberts, 2006. Aluminium-induced injury to kidney proximal tubular cells: Effects on markers of oxidative damage. J. Trace Elem. Med. Biol., 19: 267-273.
CrossRefPubMedDirect Link - Nourooz-Zadeh, J., A. Rahimi, J. Tajaddini-Sarmadi, H. Tritschler, P. Rosen, B. Halliwel and D.J. Betteridge, 1997. Relationships between plasma measures of oxidative stress and metabolic control in NIDDM. Diabetologica, 40: 647-653.
CrossRefDirect Link - Campbell, A., 2002. The potential role of aluminium in Alzheimer's disease. Nephrol. Dial. Transplant, 17: 17-20.
CrossRefDirect Link - Deloncle, R. and O. Guillard, 1990. Mechanism of Alzheimer's disease: Arguments for a neurotransmitter-aluminium complex implication. Neurochem. Res., 15: 1239-1245.
CrossRefDirect Link - Rodriguez-Puertas, R., A. Pazos, J.J. Zarranz and J. Pascual, 1994. Selective cortical decrease of high-affinity choline uptake carrier in Alzheimer's disease: An autoradiographic study using 3H-hemicholinium-3. J. Neural Transm.-Parkinson's Dis. Dementia Sect., 8: 161-169.
CrossRefDirect Link - Hammond, P. and S. Brimijoin, 1988. Acetylcholinesterase in Huntington's and Alzheimer's diseases: Simultaneous enzyme assay and immunoassay of multiple brain regions. J. Neurochem., 50: 1111-1116.
CrossRefDirect Link - Fishman, E.B., G.C. Siek, R.D. MacCallum, E.D. Bird, L. Volicer and J.K. Marquis, 1986. Distribution of the molecular forms of acetylcholinesterase in human brain: Alterations in dementia of the Alzheimer type. Ann. Neurol., 19: 246-252.
CrossRefDirect Link - Niciforovic, N. and H. Abramovic, 2014. Sinapic acid and its derivatives: Natural sources and bioactivity. Compr. Rev. Food Sci. Food Saf., 13: 34-51.
CrossRefDirect Link - Bunzel, M., J. Ralph, H. Kim, F. Lu and S.A. Ralph et al., 2003. Sinapate dehydrodimers and sinapate-ferulate heterodimers in cereal dietary fiber. J. Agric. Food Chem., 51: 1427-1434.
CrossRefDirect Link - He, L., H.T. Li, S.W. Guo, L.F. Liu, J.B. Qiu, F. Li and B.C. Cai, 2008. Inhibitory effects of sinapine on activity of acetylcholinesterase in cerebral homogenate and blood serum of rats. China J. Chin. Mater. Med., 33: 813-815.
Direct Link - Ferreres, F., F. Fernandes, C. Sousa, P. Valentao, J.A. Pereira and P.B. Andrade, 2009. Metabolic and bioactivity insights into Brassica oleracea var. acephala. J. Agric. Food Chem., 57: 8884-8892.
CrossRefDirect Link - Karakida, F., Y. Ikeya, M. Tsunakawa, T. Yamaguchi, Y. Ikarashi, S. Takeda and M. Aburada, 2007. Cerebral protective and cognition-improving effects of sinapic acid in rodents. Biol. Pharm. Bull., 30: 514-519.
CrossRefDirect Link - Erazi, H., W. Sansar, S. Ahboucha and H. Gamrani, 2010. Aluminum affects glial system and behavior of rats. Comptes Rendus Biologies, 333: 23-27.
CrossRefDirect Link - Khan, K.A., N. Kumar, P.G. Nayak, M. Nampoothiri and R.R. Shenoy et al., 2013. Impact of caffeic acid on aluminium chloride‐induced dementia in rats. J. Pharm. Pharmacol., 65: 1745-1752.
CrossRefDirect Link - Abdel-Aal, R.A., A.A.A. Assi and B.B. Kostandy, 2011. Rivastigmine reverses aluminum-induced behavioral changes in rats. Eur. J. Pharmacol., 659: 169-176.
CrossRefDirect Link - Pratico, D. and J.Q. Trojanowski, 2000. Inflammatory hypotheses: Novel mechanisms of Alzheimer's neurodegeneration and new therapeutic targets? Neurobiol. Aging, 21: 441-445.
CrossRefPubMedDirect Link - Pratico, D. and N. Delanty, 2000. Oxidative injury in diseases of the central nervous system: Focus on Alzheimer's disease. Am. J. Med., 109: 577-585.
CrossRefPubMedDirect Link