
Tajdar Husain Khan
Department of Pharmacology, College of Pharmacy, Salman Bin Abdulaziz University, P.O. Box 173, Al-Kharj 11942, Kingdom of Saudi Arabia
International Journal of Pharmacology
Year: 2012 | Volume: 8 | Issue: 1 | Page No.: 30-38







ABSTRACT
In this study, we have examines the impact of dietary soy on CCl4 induced oxidative stress, early tumor promoter markers and liver toxicity in male wistar rats. The dietary exposure mimics the repetitive nature of soy intake in human diets. Administration of soy flour (10 and 15% as diet) seven consecutive days as preventive intervention suppresses the lipidperoxidation (LPO), xanthine oxidase (XO) and release of serum toxicity marker enzymes viz., SGOT, LDH and SGPT significantly (p<0.001). Hepatic antioxidant status viz., reduced glutathione (GSH), glutathione reductase (GR), glutathione peroxidase (GPx), quinone reductase (QR), catalase (CAT) was concomitantly restored in soy flour pre-treated groups (p<0.001). In addition, soy diet pre-treatment also prevented the CCl4 enhanced ornithine decarboxylase (ODC) and hepatic DNA synthesis significantly (p<0.001). In conclusion, Carbon tetrachloride-induced liver toxicity was markedly attenuated by soy flour pre-treatment and study gives some insight of mechanisms involved in diminution of free radical generating toxicants and enhancement of antioxidant armory, hence preventing further tissue damage, injury and hyper-proliferation.
PDF Abstract XML References Citation
Received: October 10, 2011;
Accepted: December 18, 2011;
Published: February 15, 2012
How to cite this article
Tajdar Husain Khan, 2012. Soy Diet Diminish Oxidative Injure and Early Promotional Events Induced by CCl4 in Rat Liver. International Journal of Pharmacology, 8: 30-38.
DOI: 10.3923/ijp.2012.30.38
URL: https://scialert.net/abstract/?doi=ijp.2012.30.38
DOI: 10.3923/ijp.2012.30.38
URL: https://scialert.net/abstract/?doi=ijp.2012.30.38
INTRODUCTION
Oxidative stress occurs when there is an imbalance between Reactive Oxygen Species (ROS) formation and scavenging by antioxidants. ROS can cause oxidative damage to biomolecules resulting mutagenesis and carcinogenesis (Aggarwal et al., 2009; Rajendran et al., 2008; Jahan et al., 2011). ROS has been implicated in many degenerative diseases, collectively with aging and cancer (Aggarwal et al., 2009; Hamilton, 2007; Morse and Stoner, 1993; Mohamed et al., 2011; Avci et al., 2005). Carbon tetrachloride induced hepatotoxicity via hepatocellular fatty degeneration and centrilobular necrosis (Ha and Lee, 2003). The widely accepted mechanism of CCl4¯ initiated liver injury involves the bioactivation of CCl4 by the cytochrome P450 mediated reactions to the CCl3 free radical which is further converted to a peroxy radical, CCl3O2. These free radicals readily react with polyunsaturated fatty acids to initiate lipid peroxidation. Peroxy radicals can react with polyunsaturated fatty acids in presence of cellular O2, to initiate series of self-propagating chain reactions that lead to lipid peroxidation (Ha and Lee, 2003; Das et al., 2004; Alisi et al., 2011; Dhanasekaran and Ganapathy, 2011).
Antioxidants protect cells from free radical mediated oxidative damage. Cellular damage initiated by free radicals has been implicated in the development of cancer (Khan et al., 2006; Khan and Sultana, 2006; Khan and Sultana, 2011; Ramesh et al., 2007; Ali et al., 2010; Onocha et al., 2011). It is essential to identify antioxidant, anti-promoting agents present in human diet. Therefore, we have been constantly working on the identification of chemopreventive plants (Khan et al., 2006; Khan and Sultana, 2011, 2004a, 2004b, 2005; Khan et al., 2005; Khan and Sultana, 2009). The significance of nutrition in protecting living organisms from the toxic effects of environmental carcinogens has been realized (Aggarwal et al., 2009; Onocha et al., 2011; Gourineni et al., 2010). Herbal products are gaining increasingly attention due to less toxicity and high efficacy against various diseases (Khan et al., 2006; Hudecava and Ginter, 1992; Gourineni et al., 2010). Soybeans are unique foods, contains complex carbohydrates, protein, dietary fiber, oligosaccharides and isoflavones (Lichtenstein, 1998). Soybean decreases the risk of various diseases and pathological conditions, including various types of cancers, osteoporosis, menopausal symptoms and coronary heart disease (Archer 1988; Barnes, 2010; Nagata, 2010; Steinmetz and Potter, 1991; Pugalendhi and Manoharan, 2010). The chemopreventive effects of soybean and soy containing food may be related to genistein, daidzein and glycitein (Archer, 1988; Nagata, 2010; Messina and Barnes, 1991; Wei et al., 1995). These isoflavones may be conjugated with a 7-O-b-glucoside, a 6’’-O-malonylglucoside or a 6’’-O-acetylglucoside group which play an important role in the prevention atherosclerosis, hypertension (Park et al., 2005), breast cancer (DiSilvestro et al., 2005) and inflammatory bowel syndrome (Wiseman, 2006). Flavonoids have gained importance as scavengers of free radicals and a potent inhibitor of lipidperoxidation (Khan and Sultana, 2006; Bors et al., 1990; Hodgson et al., 1996). Isoflavones show DNA topoisomerase activity, synthesis and release of TGFβ, modulation of apoptosis and potent inhibitory activity of tyrosine specific protein kinases (Barnes, 2010; Nagata, 2010; Birt et al., 2001). It has been reported that population having high intake of isoflavones show lower incidence of cardiovascular diseases, osteoporosis, kidney diseases and cancer risk (Bors et al., 1990; Nagata, 2010).
Keeping in view of complex health promoting potential of soybeans and soy foods we have assumed that the soy diet may inhibit oxidative stress and toxicity induced by CCl4.
MATERIALS AND METHODS
Chemicals: EDTA, Tris, Reduced glutathione (GSH), oxidized glutathione (GSSG), nicotinamide adenine dinucleotide phosphate reduced (NADPH), Bovine Serum Albumin (BSA), 1,2-dithio-bisnitrobenzoic acid (DTNB), 1-chloro-2, 4-dinitro benzene (CDNB), phenyl methylsulfonylfluoride (PMSF), nitrobluetetrazolium (NBT), phenylmethylsulfonylfluoride (PMSF), Brij-35, Pyridoxal-phosphate, 2-mercaptoethanol, dithiothreitol, Tween 80 were obtained from Sigma chemicals Co (St Louis, MO). DL [14C] ornithine and [3H] thymidine were purchased from Amersham Corporation (Little Chalfort, UK). All other chemicals were of the highest purity and commercially available.
Animals: Eight week old adult male wistar rats (150-200 g) were obtained from the Central Animal House Facility of Hamdard University, New Delhi and were housed in a ventilated room at 25±2°C under a 12 h light/dark cycle. The animals were acclimatized for one week before the study and had free access to standard laboratory feed (Hindustan Lever Ltd., Bombay, India) and water ad libitum. All procedures using animals were reviewed and approved by the Institutional Animal Ethical Committee that is fully accredited by the Committee for Purpose of Control and Supervision on Experiments on Animals (CPCSEA) Chennai, India.
Experimental design: The treatment regimen for soy diet was based on the preliminary studies carried out in our laboratory. To study the biochemical, serological changes 25 male wistar rats were divided into five groups. Group I served as saline treated control. Group II served as negative control and was administered CCl4 (1 mL kg-1 body weight p.o., in corn oil [1:1]). Group III served as positive control and was given higher dose (D2) of soy diet for seven consecutive days. Groups IV and V were pretreated with soy diet at doses 10 and 15% soy flour as diet for seven consecutive days followed by CCl4 intoxication on the 7th day. All animals were sacrificed 24 h after CCl4 intoxication. Serum was separated and stored at 4°C for the estimation of GOT, GPT and LDH. Tissue was processed for the estimation of hepatic ornithine decarboxylase (ODC) activity and other biochemical parameters. For [3H] thymidine incorporation study, same treatment regimen was followed except all the animals were given intraperitoneal [3H] thymidine (25 μCi per animal) 2 h before killing. Time of sacrifice was after 48 h of CCl4 intoxication; liver sections were quickly excised, rinsed with ice-cold saline, freed of extraneous material and processed for the quantification of [3H] thymidine incorporation into the hepatic DNA.
Biochemical estimations: Tissue processing and preparation of Post Mitochondrial Supernatant (PMS) were done as described by Athar and Iqbal (1998). All the biochemical estimations were completed within 24 h of animal sacrifice.
Estimation of reduced glutathione: Reduced glutathione was determined by the method of Jollow et al. (1974). A 1.0 mL sample of PMS was precipitated with 1.0 mL of sulfosalicylic acid (4%). The samples were kept at 4°C for one hour and then centrifuged at 1200 g for 20 min at 4°C. The assay mixture contained 0.4 mL supernatant, 2.6 mL sodium phosphate buffer (0.1 M, pH 7.4) and 0.2 mL DTNB (100 mM) in a total volume of 3.0 mL. The yellow colour developed, was read immediately at 412 nm on a spectrophotometer.
Catalase activity: Catalase activity was measured by the method of Clairborne (1985). The reaction mixture consisted of 2 mL phosphate buffer (0.1 M, pH 7.4), 0.95 mL hydrogen peroxide (0.019 M) and 0.05 mL PMS in a final volume of 3 mL. Changes in absorbance were recorded at 240 nm. Catalase activity was calculated as nmol H2O2 consumed min-1 mg-1 protein.
Glutathione-S-transferase activity: Glutathione-S-transferase activity was estimated by the method of Habig et al. (1974).The reaction mixture consisted of 1.425 mL sodium phosphate buffer (0.1 M, pH 7.4), 0.2 mL reduced glutathione (1 mM), 0.025 mL CDNB (1 mM) and 0.3 mL PMS (10% w/v) in a total volume of 2.0 mL. The changes in the absorbance were recorded at 340 nm and enzymes activity was calculated as nmol CDNB conjugate formed min-1 mg-1 protein using a molar coefficient of 9.6x103 M-1 cm-1.
Glutathione reductase activity: Glutathione reductase activity was assayed by the method of Carlberg and Mannervik (1975), as modified by Mohandas et al. (1984). The assay system consisted of 0.1 M, pH 7.4 sodium phosphate buffer, 0.5 mM EDTA, 1 mM GSSG, 0.1 mM NADPH and PMS (10% w/v) in a total mixture of 2.0 mL. The enzyme activity was measured at 340 nm and calculated using a molar extinction coefficient of 6.22x103 M cm-1.
Glutathione peroxidase activity: Glutathione peroxidase activity was assayed by the method of Mohandas et al. (1984), as described by Athar and Iqbal (1998). The assay mixture consisted of 0.1 M, pH 7.4 sodium phosphate buffer, 1 mM EDTA, 0.2 mM NADPH, 1 mM sodium azide, 1 IU mL-1 glutathione reductase, 0.25 mM H2O2 and PMS (10% w/v) in a total volume of 2.0 mL. The activity was recorded at 340 nm and calculated using a molar extinction coefficient of 6.22x103 M-1 cm-1.
Glucose-6-phosphate dehydrogenase activity: The activity of glucose-6-phosphate dehydrogenase was assayed by the method of Zaheer et al. (1965). The reaction mixture consisted of 0.3 mL tris-HCl buffer (0.05 M, pH 7.6), 0.1 mL NADP (0.1 mM), 0.1 mL glucose-6-phosphate (0.8 mM), 0.1 mL MgCl2 (8 mM), 0.3 mL PMS and 2.4 mL distilled water in a total volume of 3 mL. The changes in absorbance were recorded at 340 nm and enzyme activity was calculated as nmol NADPH oxidized min-1 mg-1 protein using a molar extinction coefficient of 6.22x103 M-1 cm-1.
Quinone reductase activity: Quinone reductase activity was measured by the method of Benson et al. (1980). The assay mixture consisted of 0.1 mL cytosolic fraction (10%), 0.7 mL of BSA (0.1%), 0.02 mL tween-20 (1%), 0.1 mL of FAD (150 M), 0.02 mL of NADPH (0.2 mM), 0.05 mL of 2,6, DCIP (0.29%) and 2 mL of tris-HCl buffer (25 mM, pH 7.4) with a final volume of 3 mL and the optical density was read at 600 nm for 3 min. The enzyme activity was calculated as nmol 2,6, DCIP reduced min-1 mg-1 protein.
Xanthine oxidase activity: The activity of xanthine oxidase was assayed by the method of Stirpe and Corte (1969). The reaction mixture consisted of 0.2 mL PMS which was incubated for five minutes at room temperature with 0.8 mL phosphate buffer (0.1 M, pH 7.4). Then 0.1 mL xanthine (9 mM) was added to the reaction mixture and kept at 37°C for 20 min which was followed by the addition of 0.5 mL of 10% perchloric acid and 2.4 mL of double distilled water in a total volume of 4 mL. After 10 min, the mixture so obtained was centrifuged at 4000-rev min-1 for 10 min and μg uric acid formed min-1 mg-1 protein was recorded at 290 nm.
Estimation of lipid peroxidation: The assay of microsomal lipid peroxidation was done according to the method of Wright et al. (1981). The reaction mixture consisted of 0.58 mL phosphate buffer (0.1 M, pH 7.4), 0.2 mL microsome, 0.2 mL ascorbic acid (1 mM) and 0.02 mL ferric chloride (100 mM) in a total volume of 1 mL. The mixture was incubated at 37°C in a shaking water bath for 1 h. Then 1 mL 10% trichloroacetic acid and 1 mL 0.67% TBA was added. All the tubes were placed in a boiling water bath for 20 min. The tubes were placed in an ice bath and then centrifuged at 2500 g for 10 min. The amount of malanodialdehyde (MDA) formed in each of the samples was assayed by measuring the optical density of the supernatant at 535 nm.The results were expressed as nmol MDA formed h-1 g-1 tissue at 37°C using a molar extinction coefficient of 1.56x105 M-1 cm-1.
Lactate dehydrogenase (LDH) activity: Lactate dehydrogenase activity was estimated in serum by the method of Kornberg (1955). The reaction mixture consisted of serum, NADH (0.02 M), Sodium pyruvate (0.01 M), sodium phosphate buffer (0.1 M, pH 7.4) and distilled water in a total volume of 3.0 mL. The changes in the absorbance were recorded at 340 nm and enzyme activity was calculated as nmol NADH oxidized min-1 mg-1 protein.
Determination of serum oxaloacetate and pyruvate transaminases (GOT and GPT): Serum GOT and GPT were determined by the method of Reitman and Frankel (1957). Ach substrate (0.5 mL) (2 mM α-ketoglutarate and either 200 mM α L-Alanine or L-Aspartate was incubated for 5 min at 37°C in a water bath. Serum (0.1 mL) was then added and the volume was adjusted to 1.0 mL with sodium phosphate buffer. The reaction mixture was incubated for exactly 30 and 60 min for GPT and GOT, respectively. Then to the reaction mixture, 0.5 mL of DNPH (1 mM) was added and left for another 30 min at room temperature. Finally, the colour was developed by addition of 5.0 mL of NaOH (0.4 N) and product read at 505 nm.
Estimation for tumor markers
Ornithine decarboxylase activity: ODC activity was determined using 0.4 mL hepatic 105,000 g supernatant fraction per assay tube by measuring the release of CO2 from DL-[14C] ornithine by the method of O’Brien et al. (1975). The liver was homogenized in Tris-HCl buffer (pH 7.5, 50 mM) containing EDTA (0.4 mM), pyridoxal phosphate (0.32 mM), PMSF (0.1 mM), 2-mercaptoethanol (1.0 mM), dithiothreitol (4.0 mM) and Tween 80 (0.1%) at 4°C using a Teflon-glass homogenizer. In brief, the reaction mixture contained 400 μL enzymes and 0.095 mL co-factor mixture containing pyridoxal phosphate (0.32 mM), EDTA (0.4 mM), dithiothreitol (4.0 mM), ornithine (0.4 mM), Brij 35 (0.02%) and DL-[14C] ornithine (0.05 μCi) in total volume of 0.495 mL. After adding buffer and cofactor mixture to blank and others tubes, the tubes were closed immediately with a rubber stopper containing 0.2 mL ethanolamine and methoxyethanol mixture (2:1) in the central well and kept in water-bath at 37°C. After 1 h of incubation, injecting 1.0 mL citric arrested the enzyme activity acid solution (2.0 M) along the sides of glass tubes and the solution was continued for 1 h to ensure complete absorption of CO2. Finally, the central well was transferred to a vial containing 2 mL ethanol and 10 mL toluene based scintillation fluid. Radioactivity was counted in liquid scintillation counter (LKB Wallace-1410). ODC activity was expressed as pmol CO2 released h-1 mg-1 protein.
Hepatic DNA synthesis: The isolation of hepatic DNA and incorporation of [3H] thymidine in DNA was done by the method of Smart et al. (1986). Liver was quickly removed cleaned free of extraneous material and homogenate (10% w/v) was prepared in ice-cold water. The precipitate thus obtained was washed with cold trichloroacetic acid (TCA) (5%) and incubated with cold perchloric acid (PCA) (10%) at 4°C for overnight. After the incubation it was centrifuged and the precipitate was washed with cold PCA (5%). The precipitate was dissolved in warm PCA (10%) followed by incubation in boiling water bath for 30 min and filtered through Whatman 50. The filtrate was used for [3H] thymidine counting in liquid scintillation counter (LKB-Wallace-1410) by adding the scintillation fluid. The amount of DNA in the filtrate was estimated by diphenylamine method of Giles and Myers (1965). The amount of [3H] thymidine incorporated was expressed as DPM μg-1 DNA.
Protein estimation: Protein content in all samples was estimated by the method of Lowry et al. (1951) using bovine serum albumin as standard.
Statistical analysis: The level of significance between different groups is based on ANOVA test followed by Dunnett's t test.
RESULTS
CCl4 intoxication leads to depletion of hepatic glutathione, its metabolizing enzymes GST and GR and antioxidant enzymes CAT, GPx, QR and G6PD by (p<0.001), respectively as compared with the saline treated control group. CCl4 also caused elevation in the activity of XO and H2O2 content and increase in the levels of MDA formation and hepatic toxicity markers SGOT and SGPT by (p<0.001). Pretreatment with soy diet (10 and 15%) restored hepatic glutathione content and its dependent enzymes GST and GR significantly (p<0.001) as shown in Table 1. Other antioxidant enzymes like CAT GPx, G6PD and QR were significantly restored at (p<0.001) as shown in Table 2.
There was a marked depletion in levels of XO and MDA formation at (p<0.001) and concomitant down regulation of release of SGOT and SGPT in serum at (p<0.001). Figure 1 and 2 show the significant inhibition of early markers of tumor promotion like ODC activity (p<0.001) and hepatic DNA synthesis (p<0.005) in rat. The prophylactic treatment of animals with soy diet against CCl4 induced elevation in XO level, H2O2 and MDA formation is shown in Table 3.
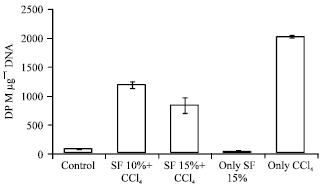 | |
| Fig. 1: | Effect of pretreatment of SF on 3[H] thymidine incorporation in hepatic DNA |
| Table 1: | Effect of pretreatment of soy diet on the CCl4 mediated depletion in the activity of glutathione content and its metabolizing enzymes in the liver of wistar rats |
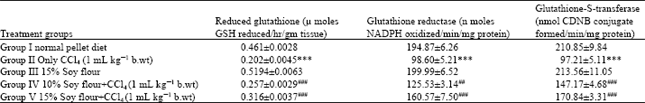 | |
| *p≤0.05, **p≤0.01 and ***p≤0.001 on comparison of normal control with CCl4 treated group and only soy group. #p≤0.05 ##p≤0.01 and ###p≤0.001 on comparison of soy diet treated groups with only CCl4 treated group | |
| Table 2: | Effect of pretreatment of soy diet on the CCl4 mediated depletion in the activity of antioxidant enzymes in the liver of wistar rats |
 | |
| *p≤0.05, **p≤0.01 and ***p≤0.001 on comparison of normal control with CCl4 treated group and only soy group. #p≤0.05, ##p≤0.01 and ###p≤0.001 on comparison of soy diet treated groups with only CCl4 treated group | |
| Table 3: | Effect of pretreatment of soy diet on the CCl4 mediated alteration in the activities of xanthine oxidase and enhancement of lipid peroxidation in the liver of wistar rats |
 | |
| *p≤0.05, **p≤0.01 and ***p≤0.001 on comparison of normal control with CCl4 treated group and only soy group. #p≤0.05, ##p≤0.01 and ###p≤0.001 on comparison of soy diet treated groups with only CCl4 treated group | |
| Table 4: | Effect of pretreatment of soy diet on the CCl4 mediated enhancement of liver markers in wistar rats |
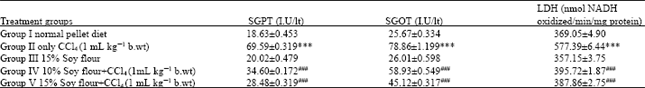 | |
| *p≤0.05, **p≤0.01 and ***p≤0.001 on comparison of normal control with CCl4 treated group and only soy group. #p≤ 0.05, ##p≤ 0.01 and ###p≤0.001 on comparison of soy diet treated groups with only CCl4 treated group | |
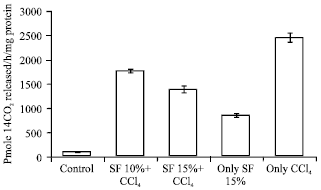 | |
| Fig. 2: | Effect of pretreatment of SF on ODC activity in liver |
Prophylactic administration of soy diet prior to CCl4 intoxication significantly decreased the MDA levels at significance (p<0.001), respectively in a dose dependent manner. However, soy diet alone group produced results near to saline control values. Soy diet showed a significant protection of GSH content by p<0.001 in a dose dependent manner is shown in Table 1. The effect of prophylactic administration of soy diet on CCl4 mediated leakage of liver marker enzymes in serum is shown in Table 4. Marked inhibition was recorded in serum toxicity markers in soy diet treated groups at (p<0.001). The effect of prophylactic treatment of rats with soy diet on CCl4 induced rate of [3H] thymidine incorporation into hepatic DNA has been shown in Fig. 2. Intoxication with CCl4 resulted in significant (p<0.005) increase in the rate of [3H] thymidine incorporation into hepatic DNA and significant (p<0.001) increase in ODC activity which is an early marker of tumor promotion. The prophylactic treatment of rats with soy diet showed a marked (p<0.001) suppressing effect on the rate [3H] thymidine incorporation into hepatic DNA of treated control and simultaneous and significant (p<0.001) inhibition of ODC activity was recorded in a dose dependent manner as shown in Fig. 1 and 2.
DISCUSSION
Single oral CCl4 dose induced significant depletion in the hepatic GSH content and its metabolizing enzymes. Glutathione is often considered as first line of defense against oxidative damage (Ali-Osman, 1989; Patrick-Iwuanyanwu and Wegwu, 2008). There was significant restoration of glutathione and dependent enzymes in soy diet pretreated groups. The restoration of depleted GSH and GST on pretreatment of animals with soy diet indicate its role as chemopreventive measure. CCl4 causes increased formation of pro-oxidants and a concomitant decrease in the antioxidant status of the cells. Hepatotoxic effects of CCl4 depend on the major unstable radical trichloromethyl radical which reacts with oxygen molecule and form trichloromethylperoxy radical which is reported to be highly reactive. These free radicals covalently bind to macromolecules and proteins and thus initiating a site for secondary biochemical process which ultimately leads to several pathological conditions of CCl4 metabolism (Singh et al., 2005; Hassan et al., 2008). It has been suggested that Soy diet protects by suppressing CCl4 mediated toxicity through decrease in levels of MDA formation and inhibition in hepatic XO levels. GST is responsible for GSH conjugation with various electrophiles and plays an important role in detoxification of various xenobiotics (Touliatos et al., 2000). Induction of GST by Soy diet could be one possible mechanism to combat hepatic oxidative stress (Singh et al., 2000; Dhanasekaran and Ganapathy, 2011). QR catalyzes hydroquinone formation from quinones, preventing cyt P450 dependent activation to toxic semiquinones and ultimate generation of Reactive Oxygen Species (ROS) (Khan and Sultana, 2005). Well documented correlation exists between its modulation and chemoprevention of cancer Substantial depression in hepatic GSH with concomitant decrease in GR, GPx, QR, GST and G6PD levels on CCl4 administration; however, dose dependent marked restoration was observed with prophylactic treatment of Soy diet.
The mechanisms by which Soy diet attenuates hepatic damage induced by CCl4 can be attributed to recovery of antioxidant status. Previous studies have shown that Soy potential to increase cellular antioxidants like GSH and other phase II metabolizing enzymes (Khan and Sultana, 2004a, b; Khan et al., 2005; Borras et al., 2010). Soy diet prior to CCl4 intoxication restored the levels of serum toxicity markers viz., SGOT and SGPT as compared to negative control. CCl4 administration resulted in significant induction in hepatic ODC and [3H] thymidine uptake into hepatic DNA, There is enough evidence that indicates that the polyamines play essential role in the regulation of various cellular and metabolic functions including DNA synthesis, modulation of membranous functions and numerous enzyme activities (Wallon and O'Brien, 2005). Prophylactic treatment with Soy diet prior to CCl4 intoxication showed profound suppression ODC activity and rate of [3H] thymidine incorporation.
CONCLUSION
Modulatory action of Soy diet is observed by several mechanisms, it reduces the risk of acute hepatic injury induced by CCl4 and thus protecting against post necrotic hepatic injury. In conclusion present study give some insight into mechanisms involved in modulatory action of Soy diet against CCl4 induced hepatic toxicity in rats. Thus, we suggest it may further be used as potent cancer chemopreventive agent.
ACKNOWLEDGEMENT
Dr Tajdar Husain Khan is thankful to the Council of Scientific and Industrial Research (CSIR), New Delhi, India, for providing the funds to carry out this study.
REFERENCES
- Ali-Osman, F., 1989. Quenching of DNA cross-link precursors of chloroethylnitrosoureas and attenuation of DNA interstrand cross-linking by glutathione. Cancer Res., 49: 5258-5261.
PubMed - Ali, H.F.M., F.M.A. El-Ella and N.F. Nasr, 2010. Screening of chemical analysis, antioxidant antimicrobial and antitumor activities of essential oil of oleander (Nerium oleander) flower. Int. J. Biol. Chem., 4: 190-202.
CrossRef - Alisi, C.S., A.O. Ojiako, G.O.C. Onyeze and G.C. Osuagwu, 2011. Normalisation of lipoprotein phenotypes by Chromolaena odorata-linn. in carbon tetrachloride hepatotoxicity-induced dyslipidaemia. Am. J. Drug Discov. Dev., 1: 209-219.
CrossRefDirect Link - Archer, V.E., 1988. Cooking methods, carcinogens and diet-cancer studies. Nutr. Cancer, 11: 75-79.
PubMed - Avci, A., M. Kacmaz, M. Kavutcu, E. Gocmen and I. Durak, 2005. Effects of an antioxidant extract on adenosine deaminase activities in cancerous human liver tissues. Int. J. Cancer Res., 1: 53-56.
CrossRefDirect Link - Athar, M. and M. Iqbal, 1998. Ferric nitrilotriacetate promotes N-diethylnitrosamine-induced renal tumorigenesis in the rat: Implications for the involvement of oxidative stress. Carcinogenesis, 19: 1133-1139.
PubMedDirect Link - Benson, A.M., M.J. Hunkeler and P. Talalay, 1980. Increase of NADPHl: Quinone reductase activity by dietary antioxidant: Possible role in protection against carcinogenesis and toxicity. Proc. Nat. Acad. Sci., 77: 5216-5220.
PubMed - Birt, D.F., S. Hendrich and W. Wang, 2001. Dietary agents in cancer prevention: Flavonoids and isoflavonoids. Pharmacol. Ther., 90: 157-177.
CrossRef - Bors, W., W. Heller, C. Michel and M. Saran, 1990. Flavonoids as anti-oxidants: Determination of radical scavenging efficiencies. Methods Enzymol., 186: 343-355.
PubMedDirect Link - Carlberg, I. and B. Mannervik, 1975. Purification and characterization of the flavoenzyme glutathione reductase from rat liver. J. Biol. Chem., 250: 5475-5480.
PubMed - Dhanasekaran, J.J. and M. Ganapathy, 2011. Hepatoprotective effect of Cassia auriculata L. leaf extract on carbon tetrachloride intoxicated liver damage in wister albino rats. Asian J. Biochem., 6: 104-112.
CrossRefDirect Link - Das, R.K., S. Das and S. Bhattacharya, 2004. Protective effect of diphenylmethylselenocyanate against carbon tetrachloride-induced hepatotoxicity in vivo. J. Environ. Pathol. Toxicol. Oncol., 23: 287-296.
PubMed - DiSilvestro, R.A., J. Goodman, E. Dy and G. Lavalle, 2005. Soy isoflavone supplementation elevates erythrocyte superoxide dismutase, but not plasma ceruloplasmin in postmenopausal breast cancer survivors. Breast Cancer Res. Treat., 89: 251-255.
PubMed - Giles, K.W. and A. Myers, 1965. An improved diphenylamine method for the estimation of deoxyribonucleic acid. Nature, 206: 93-93.
CrossRefDirect Link - Gourineni, V.P., M. Verghese and J. Boateng, 2010. Anticancer effects of prebiotics synergy1® and soybean extracts: Possible synergistic mechanisms in caco-2 cells. Int. J. Cancer Res., 6: 220-233.
CrossRefDirect Link - Ha, B.J. and J.Y. Lee, 2003. The effect of chondroitin sulfate against CCl4-induced hepatotoxicity. Biol. Pharmaceut. Bull., 26: 622-626.
PubMed - Habig, W.H., M.J. Pabst and W.B. Jakoby, 1974. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem., 249: 7130-7139.
CrossRefPubMedDirect Link - Hamilton, K.L., 2007. Antioxidants and cardioprotection. Med. Sci. Sports Exerc., 39: 1544-1553.
PubMed - Hassan, S.W., R.A. Umar, A.A. Ebbo, A.J. Akpeji and I.K. Matazu, 2008. Hepatoprotective effect of leaf extracts of Parkinsonia aculeata L. against CCl4 intoxication in albino rats. Int. J. Biol. Chem., 2: 42-48.
CrossRefDirect Link - Hodgson, J.M., K.D. Croft, I.B. Puddey, T.A. Mori and L.J. Beilin, 1996. Soybean isoflavonoids and their metabolic products inhibit in vitro lipoprotein oxidation in serum. J. Nutr. Biochem., 7: 664-669.
CrossRefDirect Link - Hudecava, A. and E. Ginter, 1992. The influence of ascorbic acid on lipid peroxidation in Guinea pigs intoxicated with cadmium. Food Chem. Toxicol., 30: 1011-1013.
CrossRef - Jahan, M.S., G. Vani and C.S. Shyamaladevi, 2011. Anti-carcinogenic effect of Solanum trilobatum in diethylnitrosamine induced and phenobarbital promoted heaptocarcinogenesis in rats. Asian J. Biochem., 6: 74-81.
CrossRefDirect Link - Jollow, D.J., J.R. Mitchell, N. Zampaglione and J.R. Gillette, 1974. Bromobenzene-induced liver necrosis. Protective role of glutathione and evidence for 3,4-bromobenzene oxide as the hepatotoxic metabolite. Pharmacology, 11: 151-169.
CrossRefPubMedDirect Link - Khan, N. and S. Sultana, 2004. Abrogation of potassium bromated-induced renal oxidative stress and subsequent cell proliferation response by soy isoflavones in wistar rats. Toxicology, 201: 173-184.
PubMed - Khan, N. and S. Sultana, 2004. Induction of renal oxidative stress and cell proliferation response by ferric nitrilotriacetate (Fe-NTA): Diminution by soy isoflavones. Chem. Biol. Interact., 149: 23-35.
PubMed - Khan, N. and S. Sultana, 2005. Anticarcinogenic effect of Nymphaeaalba against oxidative damage, hyperproliferative response and renal carcinogenesis in Wistar rats. Mol. Cell Biochem., 271: 1-11.
PubMed - Khan, T.H. and S. Sultana, 2006. Apigenin induces apoptosis in Hep G2 cells: Possible role of TNF-α and IFN-γ. Toxicology, 217: 206-212.
CrossRef - Khan, T.H. and S. Sultana, 2009. Antioxidant and hepatoprotective potential of Aegle marmelos Correa. against CCl4-induced oxidative stress and early tumor events. J. Enzyme Inhib. Med. Chem., 24: 320-327.
CrossRefPubMedDirect Link - Khan, T.H. and S. Sultana, 2011. Effect of Aegle marmelos on DEN initiated and 2-AAF promoted hepatocarcinogenesis: A chemopreventive study. Toxicol. Mech. Methods, 21: 453-462.
CrossRef - Khan, T.H., L. Prasad, A. Sultan and S. Sultana, 2005. Soy isoflavones inhibits the genotoxicity of benzo(a)pyrene in Swiss albino mice. Human Exp. Toxicol., 24: 149-155.
CrossRefDirect Link - Khan, T.H., T. Jahangir, L. Prasad and S. Sultana, 2006. Inhibitory effect of apigenin on benzo(a)pyrene-mediated genotoxicity in Swiss albino mice. J. Pharm. Pharmacol., 58: 1655-1660.
CrossRef - Lichtenstein, A.H., 1998. Soy protein, isoflavones and cardiovascular disease risk. J. Nutr., 128: 1589-1592.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Messina, M. and S. Barnes, 1991. The role of soy products in reducing risk of cancer. J. Nat. Cancer Inst., 83: 541-546.
PubMed - Mohamed, J., W.L. Wei, N.N.A. Husin, N.Y. Alwahaibi and S.B. Budin, 2011. Selenium supplementation reduced oxidative stress in diethylnitrosamine-induced hepatocellular carcinoma in rats. Pak. J. Biol. Sci., 14: 1055-1060.
CrossRefDirect Link - Mohandas, J., J.J. Marshall, G.G. Duggin, J.S. Horvath and D.J. Tiller, 1984. Differential distribution of glutathione and glutathione-related enzymes in rabbit kidney: Possible implications in analgesic nephropathy. Biochem. Pharmacol., 33: 1801-1807.
CrossRefPubMedDirect Link - Morse, M.A. and G.D. Stoner, 1993. Cancer chemoprevention: Principles and prospects. Carcinogenesis, 14: 1737-1746.
CrossRefPubMedDirect Link - Nagata, C., 2010. Factors to consider in the association between soy isoflavone intake and breast cancer risk. J. Epidemiol., 20: 83-89.
CrossRefPubMedDirect Link - Onocha, P.A., G.K. Oloyede and Q.O. Afolabi, 2011. Cytotoxicity and free radical scavenging activities of hexane fractions of Nigeria specie of African pear (Dacryodes edulis). Int. J. Biol. Chem., 5: 143-149.
CrossRefDirect Link - Patrick-Iwuanyanwu, K.C. and M.O. Wegwu, 2008. Prevention of carbon tetrachloride (CCl4)-induced liver damage in rats by Acanthus montanus. Asian J. Biochem., 3: 213-220.
CrossRefDirect Link - Pugalendhi, P. and S. Manoharan, 2010. Chemopreventive potential of genistein and daidzein in combination during 7,12-dimethylbenz(a)anthracene (DMBA) induced mammary carcinogenesis in sprague-dawley rats. Pak. J. Biol. Sci., 13: 279-286.
CrossRef - Rajendran, P., R. Venugopal, G. Ekambaram, A. Aadithya and D. Sakthisekaran, 2008. Rehabilating activity of mangiferin in benzo(a) pyrene induced lung carcinogenesis. Asian J. Biochem., 36: 118-125.
CrossRefDirect Link - Ramesh, T., R. Mahesh and V. Hazeena Begum, 2007. Effect of Sesbania grandiflora on lung antioxidant defense system in cigarette smoke exposed rats. Int. J. Biol. Chem., 1: 141-148.
CrossRef - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Singh, M., V. Tiwari, A. Jain and S. Ghosal, 2005. Protective activity of picroliv on hepatic amoebiasis associated with carbon tetrachloride toxicity. Indian J. Med. Res., 121: 676-682.
PubMed - Steinmetz, K.A. and J.D. Potter, 1991. Vegetables, fruit and cancer. I. Epidemiology. Epidemiol. Cancer Causes Control, 2: 325-357.
CrossRefPubMedDirect Link - Stirpe, F. and E.D. Corte, 1969. The regulation of rat liver xanthine oxidase: Conversion in vitro of the enzyme activity from dehydrogenase (type D) to oxidase (type O). J. Biol. Chem., 244: 3855-3863.
PubMedDirect Link - Wei, H., R. Bowen, Q. Cai, S. Barnes and Y. Wang, 1995. Antioxidant and antipromotional effects of the soybean isoflavone genistein. Proc. Soc. Exp. Biol. Med., 208: 124-130.
PubMed - Wiseman, A., 2006. Crohn's disease leading to bowel cancer may be avoided by consumption of soya isoflavones: Adjunct-chemotherapy with oxaliplatin. Med. Hypotheses, 66: 934-935.
CrossRefPubMedDirect Link - Zaheer, N., K.K. Tewari and P.S. Krishnan, 1965. Exposure and solubilization of hepatic mitochondrial shunt dehydrogenases. Arch. Biochem. Biophys., 109: 646-648.
PubMed