
S. Banerjee
Department of Physics, Jadavpur University, Kolkata-700 032, India
G. Bandyopadhyaya
Department of Physics, Jadavpur University, Kolkata-700 032, India
K. Chattopadhyay
Department of Chemical Technology, Calcutta University, Kolkata-700009, India
B. D. Chattopadhyay
Department of Physics, Jadavpur University, Kolkata-700 032, India
International Journal of Pharmacology
Year: 2010 | Volume: 6 | Issue: 4 | Page No.: 444-455







ABSTRACT
Tobacco smoking is continuously increasing the risk of severe diseases like blood cancer, malfunctioning of blood immune system and peroxidative damage of erythrocytes, especially in developing countries. However, the effect of nicotine in the blood cells of protein malnourished condition and its amelioration by curcumin has not yet been studied. The present study is an attempt in that direction where female albino rats maintained under normal/protein restricted diets were subcutaneously injected with nicotine tartrate (2.5 mg kg-1 b.wt. daily for 21 days) and orally supplemented with curcumin (80 mg kg-1 b.wt.) and subsequently in vivo and in vitro experiments were performed. We report that nicotine alters the blood cell composition (RBC counts, haemoglobin content) and aggravates loss of DNA content and DNA damage in blood cells of protein malnourished female rats in comparison to normal diet fed group and curcumin significantly (p<0.001) attenuates nicotine-induced changes in blood cells of protein malnourished rats. In vitro study shows that curcumin strongly interacts with nicotine and also DNA and thereby protects the DNA from nicotine induced damages. This study also confirms that more free nicotine molecules are available in protein restricted condition which aggravates the nicotine induced toxic effect on blood cells. Thus protein malnourished animals are more susceptible to nicotine induced toxicity and curcumin is effective enough to prevent nicotine induced damage even in protein malnourished condition. The molecular mechanism underlying nicotine induced toxicity and prevention by curcumin is discussed in this study.
PDF Abstract XML References Citation
How to cite this article
S. Banerjee, G. Bandyopadhyaya, K. Chattopadhyay and B. D. Chattopadhyay, 2010. Amelioration of Nicotine-Induced Damage of Blood Cells in Protein Malnourished Female Rats by Curcumin. International Journal of Pharmacology, 6: 444-455.
DOI: 10.3923/ijp.2010.444.455
URL: https://scialert.net/abstract/?doi=ijp.2010.444.455
DOI: 10.3923/ijp.2010.444.455
URL: https://scialert.net/abstract/?doi=ijp.2010.444.455
INTRODUCTION
Nicotine is the main factor for human tobacco addiction (Dani et al., 2001). Smoking results in premature death in human and is believed to contribute in the development of cardiovascular disease (Bjartveit and Tverdal, 2005), lung cancer (International Agency for Research on Cancer, 2004) and other diseases. Every cigarette reduces the life span by about 5 min and globally, smoking kills more than 4 million people in every year (Sasikala et al., 2003; Keloglu-Isler and Erdogan, 2007). Nicotine is easily absorbed in human airways which in turn affect blood cells that result in the malfunctioning of the immune system (Sela et al., 2002). It also causes the formation of Reactive Oxygen Species (ROS) and toxic chemical loads associated with peroxidative membrane damages of erythrocytes via ROS (Tedesco et al., 2000). On the other hand, oxidized erythrocytes are more prone to form aggregates and increase the viscosity of blood stream and thus, ROS impair blood flow in the microcirculation (Chung and Ho, 1999). Cooke (2007) has shown that nicotine may contribute to tumor angiogenesis and tumor growth; atherosclerotic plaque neovascularization and progression and other tobacco-related diseases.
Curcumin is an active ingredient of the rhizome of Curcuma longa with anti-inflammatory (Chun et al., 2003), antioxidant and antiproliferative activities (Kalpana et al., 2007). It protects from cellular damage and inhibits the gene that makes inflammatory COX-2 enzymes (Chun et al., 2003). Recent study has shown that curcumin is a potent DNA hypomethylation agent (Liu et al., 2009). Earlier studies have shown that nicotine causes more toxicological effects on lipid peroxidation in malnourished condition (Chattopadhyay and Chattopadhyay, 2008) and curcumin protects nicotine induced DNA damage in liver cells in such condition (Bandyopadhyaya et al., 2008). El-Demerdash et al. (2009) has recently reported that curcumin has beneficial influences and could be able to antagonize sodium arsenite toxicity.
Presently, most of the studies related to nicotine and curcumin are performed in normal dietary conditions and no data available on the effect of nicotine in protein-restricted females as nicotine seems to be a greater risk factor in women than man (Prescott et al., 1998). Smoking affects the reproductive health of women and causes infertility, birth defects, ectopic pregnancy and spontaneous abortions (De Mallo et al., 2001; Practice Committee of the American Society for Reproductive Medicine, 2004). The present study was designed to investigate the effect of nicotine on blood cells in female rats under protein restricted dietary condition and the protective role of curcumin thereof, in terms of different end points (haemoglobin percentage, red blood cells count, total DNA concentration, DNA damage by comet assay, in vitro interactions of nicotine, curcumin vs. DNA and protein and molecular docking).
MATERIALS AND METHODS
Chemicals: Nicotine hydrogen tartrate and curcumin were purchased in 2005 from Sigma Chemicals Company, St. Louis, USA. All other analytical grade chemicals were supplied in 2005 and 2006 by Spectrochem Pvt. Ldt. India and Genei, Bangalore, India.
Diets and treatments: Thirty adult female albino rats (body weights 120- 130 g) of Wistar strain were procured from the animal housing facility and acclimatized under standard conditions of temperature and humidity with 12 h light/dark cycles. They were maintained in accordance to the guidelines of Instructional Animal Ethics Committee of Jadavpur University, Kolkata, India as described elsewhere (Bandyopadhyaya et al., 2008). One week before the treatment, the animals were fed standard pellet diet (Hindustan Lever Ltd., India) and water ad libitum. Thirty female rats were divided equally (n = 5) into the following groups. Group 1: fed with normal protein diet (18% casein, 70% carbohydrate, 7% fat, 4% salt mixture and 1% vitamin mixture) Group 2: fed with normal protein diet and treated with nicotine; Group 3: fed with normal protein diet and treated with nicotine + curcumin; Group 4: female rats fed with protein restricted diet (5% casein, 83% carbohydrate, 7% fat, 4% salt mixture and 1% vitamin mixture); Group 5: fed with protein restricted diet and treated with nicotine; Group 6: fed with protein restricted diet and treated with nicotine + curcumin. Nicotine tartrate was administered subcutaneously daily at the dose of 2.5 mg kg-1 of body weight in 0.5 mL physiological saline and curcumin was administered orally at the dose of 80 mg kg-1 body weight/day. Compositions of diets were prepared as described by Hawk et al. (1954) and selection of dose, route of administration and the period of dosing were adopted from our earlier study (Mandal et al., 2004). Nicotine and curcumin administration continued for 21 days. The animals in Group 1 and 4 received subcutaneous injection of 0.5 mL physiological saline without nicotine. After the completion of treatments, animals were kept fasting overnight and sacrificed next day by decapitation.
Total Counts of Red Blood Corpuscles (RBC): RBC was counted by haemocytometer with the help of standard laboratory techniques (Chatterjee, 1994). For each animal in a group, count was made thrice and averaged. Data presented here represent the average of 10 animals in each group from two sets of experiments.
Haemoglobin estimation: Haemoglobin percentages of blood were measured by the standard Sahli’s Haemoglobinometer (Chatterjee, 1994). The instrument consisted of two tubes, one of which contained 20 mm3 of blood haemolysed with 0.1 N HCl and saturated with CO gas. The color of this tube was used as standard. In the other tube an equal volume of 0.1 N HCl was taken and 20 mm3 of test blood from the rats was added. HCl converts hemoglobin into acid haematin. The color developed was matched against the standard. If the color of the unknown was stronger, it was diluted with 0.1 N HCl until the tinge was same in both. The graduation up to which the blood has been diluted gives the percentage of haemoglobin. Data presented here are from 10 animals (2 sets of experiments).
Total DNA content estimation: The total DNA content was measured by slightly modifying the protocol of DNA preparation from blood as described by National Institutes of Health (2004). To 300 μL of whole blood (EDTA), 900 μL of lysis buffer (155 mM NH4Cl, 10 mM KHCO3, 0.1 mM Na2-EDTA, pH 7.4) was added, mixed, incubated for 30 min on ice and centrifuged (1200 g for 10 min at 4°C). The blood cell pellet was resuspended in 150 μL of SE buffer (75 mM NaCl, 25 mM EDTA) and centrifuged at 1200 g for 10 min at 4°C. After removing the supernatant, 7.5 μL of 20% SDS and 1.2 μL of proteinase K (10 mg mL-1) was added, mixed gently and incubated overnight at 37°C water bath. Next day 150 μL of SE buffer and 300 μL of phenol were added, mixed for 10 min and centrifuged at 3000 g for 5 min at 10°C. The supernatant was extracted with 300 μL of phenol/chloroform/isoamyl alcohol mixture (25:24:1 v/v) and centrifuged at 3000 g for 5 min at 10°C. The supernatant was transferred to a new tube and re-extracted with 300 μL of chloroform/isoamyl alcohol (24: 1) mixture similarly. Finally, 9 μL of 3 M sodium acetate (pH 5.2) and 300 μL of isopropanol were added and mixed gently until the DNA was precipitated. The precipitated DNA was rinsed carefully in 70% ethanol and dissolved in 0.2 mL TE buffer (pH 8) and its concentration determined spectrophotometrically at A230, A260 and A280. This experiment was also repeated twice and the data represents the average of 10 animals.
Comet assay: Comet assay was performed to study DNA damage (Bandyopadhyaya et al., 2008; Kido et al., 2006). Forty microliter of blood samples were collected from the hepatic vein of the experimental rats into a heparinized glass capillary. The samples were processed within 2 h to avoid any further damages. Two hundred microliter of 2% agarose (mp. 40-42°C, Genei, India) was quickly layered on a pre-cooled fully frosted slide and covered with a cover slip and allowed to solidify at 4°C. Meanwhile, 20 μL of whole blood was added to 100 μL of 0.5% low melting point agarose (Genei, India) in a micro centrifuge tube. After carefully removing the cover slip, a second layer of 100 μL whole blood mixed with low melting agarose was pipetted out on the slide, covered carefully with the cover slip again and allowed to solidify at 4°C for 15 min. The slide (without cover slip) was immersed in a freshly prepared and chilled lysing solution (2.5 M NaCl, 100 mM EDTA, 10 mM Tris-HCl, 1% sarkosyl, 10% DMSO and 1% triton X-100, at pH 10, DMSO added just before use) and kept at 4°C for 2 h. Slides were then placed on a horizontal gel electrophoresis platform in alkaline electrophoresis solution (300 mM NaOH and 1 mM EDTA, pH 13) and left in dark for 15 min to allow unwinding of the DNA to occur. The DNA was then electrophoresed at 4°C in the dark for 15 min at 1 v/cm and ~ 250 mA. The slides were gently rinsed in neutralization buffer (0.4 M Tris-HCl, pH 7.5) and stained with 50 μL of 20 μg mL-1 ethidium bromide and covered with a cover slip. The photomicrograph of each slide was taken in Leica fluorescent microscope (Model 300 FX) at 40x magnification.
Comet parameters measurement: Comet parameters (head diameter, tail length and tail moment) and percentage of DNA damage were assessed following the procedure of Helma and Uhl (2000). For each group 4 slides were prepared and total 50 cells were screened per slide.
Spectrophotometric study: For nicotine-curcumin interaction, 3 mL of 500 μM nicotine tartrate solution was taken in a quartz cuvette and its absorbance spectrum was recorded from 230-300 nm by Elico SL-196 spectrophotometer. The concentration of nicotine was taken in such that its optical density at 260 nm was 0.4-0.6. Curcumin dissolved in ethanol was directly added at different concentrations (final strength varied from 16.6 to 96.6 μM) to the quartz cuvette, incubated 15 min to allow sufficient time for the interaction and absorbance spectra of nicotine-curcumin complex were recorded similarly. Binding constant of nicotine-curcumin interaction was determined by Binding-Isotherm plot (Lakowicz, 1983) using the following equation:
| (1) |
where, ΔA was the change in absorbance intensity of nicotine, Am was the maximum absorbance intensity of nicotine without curcumin and C was the concentration of curcumin (quencher). A graph was plotted with 1/ΔAc vs. 1/C. The binding constant K for curcumin was determined from the value of the intercept 1/Am and slope 1/(Am . K) of the plot 1/ΔAc vs. 1/C. The binding nature of curcumin with nicotine was also observed by determining the Hill coefficient of nicotine-curcumin interaction (Barcroft and Hill, 1910).
Nicotine-DNA interaction was studied by taking purified blood DNA (30 μg in 3 mL sterile deionized water) in a quartz cuvette and its absorbance spectrum was recorded from 230-300 nm similarly. Nicotine with different concentrations (final strength varied from 5 nM-1 mM) was added to the DNA solution and the absorbance spectra were observed similarly. Curcumin (final concentration varied from 16.6-96.6 μM) was then added to the nicotine-DNA mixture and the absorbance spectra were recorded similarly. DNA isolated from blood cells of rat fed with normal protein diet (control) as well as normal protein diet treated with nicotine + curcumin was used in this study.
Time kinetics of the interaction of nicotine vs. Bovine Serum Albumin (BSA) was performed similarly by taking two concentrations of BSA (0.5 and 1.5%). Nicotine tartrate solution (833 μM) was taken in quartz cuvette and its absorbance was recorded at 260 nm. BSA was added to nicotine solution and the absorbance was recorded in every 5 sec interval up to 40 sec. From this experimental result the concentration of free (unbound) nicotine was calculated by using previously calibrated nicotine standard curve. The binding nature of nicotine with BSA was also determined by using Hill coefficient from their interaction.
CD-Spectrometric study: Jasco-720 CD-spectrometer was used to study the conformational change of DNA on treatment with nicotine alone and nicotine and curcumin together. Initially CD spectra for control plots of DNA (30 μg), nicotine (1 mM) and curcumin (100 μM) were taken. Then CD spectra of DNA (30 μg) treated with increasing concentrations nicotine (50 μM-1 mM) were observed. DNA treated with 1mM nicotine was then further treated with different concentrations of curcumin (15-100 μM) and the CD-spectra were observed.
Molecular docking: Docking experiments were performed using docking interactive software Hex 5.1 (http://software.informer.com/getfree-hex-5.1-software) and Chimera (http://www.cgl.ucsf.edu/chimera/download.html) to monitor the interactions of DNA, albumin, nicotine and curcumin. Protein Data Bank (PDB) files of p53 consensus sequence (GGGCATGCCTAGGCATGCC) of human DNA (PDB ID: 3KMD), human serum albumin (PDB ID: 1R4I), nicotine (elchem.kaist.ac.kr) and curcumin (CID_969516) were fed in these softwares for the experiments. BLAST results indicated 73% homology between human serum albumin and Rattus norvegicus serum albumin protein sequences; thus human serum albumin was selected to monitor docking phenomena.
Statistical analysis: The entire experiment was repeated twice and data (n = 10 animals) were averaged and tabulated as Mean±SD. The statistical analysis of the data estimated from each group of all conditions were done by One way analysis of variance (ANOVA) and all pair wise Multiple comparison procedures (Holm-Sidak method) using Sigma Stat (version 3.2). Significance levels were examined at p<0.01 or <0.001. The correlation coefficients of the observed experimental values between each group of all dietary conditions were determined by using CORREL function of Microsoft Excel.
RESULTS
Nicotine treatment caused decrease of RBC counts in both normal diet (26.3%) - and protein restricted diet-fed (48.6%) rats compared to their respective controls. Curcumin significantly improved the RBC counts in normal diet fed (23%; p<0.01) and -protein restricted diet fed (64.3%; p<0.001) female rats compared to their respective nicotine treated rats (Table 1).
Nicotine treatment significantly reduced the haemoglobin content of blood cells in normal diet fed rat (15.16%; p<0.01) and more significantly (23.14%; p<0.001) in protein-restricted diet fed group in comparison to their respective nicotine untreated controls. Interestingly, curcumin normalized the hemoglobin level in both normal (10%, p<0.01) - and protein restricted-diet fed (21%, p<0.001) animals (Table 2).
Table 3 represents the total DNA contents of blood cells in all the groups of rats. The total DNA contents of blood cells (per 300 μL blood) were 115.38 μg in normal diet fed group and 104.69 μg in protein restricted diet groups of rats. Nicotine reduced the total DNA content of normal diet fed rats (39.6%; p<0.001) and protein restricted diet fed rats (47.6%; p<0.001), respectively compared to their respective controls. Curcumin increased the total DNA content significantly (17.65%; p<0.01) for normal diet and (29%; p<0.001) for restricted diet fed rats in comparison to their respective nicotine treated controls.
| Table 1: | RBC count when animals were treated with nicotine and nicotine + curcumin under different dietary status |
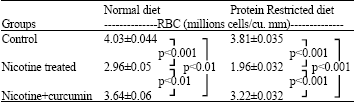 | |
| RBC is counted by haemocytometer with the help of standard laboratory techniques (Chatterjee, 1994). For each animal in a subgroup, count was made thrice and averaged. Data are shown as the average over 10 animals of each subgroup from two sets of experiments along with SD. The significance levels was p<0.01 significant and p<0.001 more significant | |
| Table 2: | Haemoglobin content in blood cells when animals were treated with nicotine and nicotine + curcumin under different dietary status |
 | |
| The standard Sahli’s Haemoglobinometer measured haemoglobin percentages of blood. The experimental setup was repeated twice and all data were averaged over n = 10 animals. Values are given Mean±SD. Significance levels were determined by ANOVA where p<0.01 was significant and p<0.001 was more significant | |
| Table 3: | Total DNA content in blood cells when animals were treated with nicotine and nicotine + curcumin under different dietary status |
 | |
| For total DNA content, to 300 μL of whole blood (EDTA), 900 μL of lysis buffer (155 mM NH4Cl, 10 mM KHCO3, 0.1 mM Na2-EDTA, pH 7.4) was added, mixed, incubated for 30 min on ice and centrifuged (1200 g for 10 min at 4°C). The blood cell pellet was resuspended in 150 μL of SE buffer (75 mM NaCl, 25 mM EDTA) and centrifuged at 1200 g for 10 min at 4°C. After removing the supernatant, 7.5 μL of 20% SDS and 1.2 μL of proteinase K (10 mg mL-1) was added, mixed gently and incubated overnight at 37°C water bath. Next day 150 μL of SE buffer and 300 μL of phenol were added, mixed for 10 min and centrifuged at 3000 g for 5 min at 10°C. The supernatant was extracted with 300 μL of phenol/chloroform/isoamyl alcohol mixture (25:24:1 v/v) and centrifuged at 3000 g for 5 min at 10°C. The supernatant was transferred to a new tube and re-extracted with 300 μL of chloroform/isoamyl alcohol (24:1) mixture similarly. Finally 9 μL of 3 M sodium acetate (pH 5.2) and 300 μL of isopropanol were added and mixed gently until the DNA was precipitated. The precipitated DNA was rinsed carefully in 70% ethanol and dissolved in 0.2 mL TE buffer (pH 8) and its concentration determined spectrophotometrically at A230, A260 and A280. The experimental setup was repeated twice and all data were averaged over n = 10 animals. Values are given mean±S.D. Significance levels were determined by ANOVA where p<0.01 was significant and p<0.001 was more significant | |
Photomicrographs of the Comet assay of blood cell DNA of animals under normal dietary condition are shown in Fig. 1A-C. Figure 1A represents the comet picture of blood cell DNA in normal diet fed rats (Gr.1) and Fig. 1B and 1C represent the comet pictures of nicotine treated (Gr. 2) and nicotine+curcumin treated (Gr. 3) normal diet fed rats, respectively.
 | |
| Fig. 1: | (A) Comet photograph of whole blood cells of female rat maintained under normal diet (Control), (B) Comet photograph of whole blood cells of female rat maintained under normal diet (treated with nicotine) and (C) Comet photograph of whole blood cells of female rat maintained under normal diet (treated with nicotine and curcumin) |
 | |
| Fig. 2: | (A) Comet photograph of whole blood cells of female rat maintained under with protein restricted diet (Control), (B) Comet photograph of whole blood cells of female rat maintained under protein restricted diet (treated with nicotine and (C) Comet photograph of whole blood cells of female rat maintained under protein restricted diet (treated with nicotine and curcumin) |
Similarly, Fig. 2A-C represent photomicrographs of the comet assay for blood cells DNA of rats fed with protein-restricted diet. The extent of blood cell DNA damage and comet tail moment were analyzed and shown in Table 4 for all the groups. Nicotine treatment increased DNA damages by 39.3% (p<0.001) in normal diet fed and by 40.4% (p<0.001) in protein-restricted diet fed animals compared to their respective controls. Whereas the DNA damages in nicotine treated and curcumin supplemented groups were 20% (p<0.01) and 11.3% (p<0.001) in normal and protein-restricted diet fed rats, respectively compared to their respective controls. Nicotine treatment showed increased tail moments by 19 times in normal diet fed rats and 10 times in protein restricted diet fed rats in comparison to their respective nicotine untreated controls. Treating animals with curcumin decreased the tail moments by 8 times in nicotine treated normal diet fed and 2.2 times in nicotine treated protein restricted diet fed group in comparison to their respective nicotine untreated controls. Data in this table are the average of observations from 4 animals in each group (for each animal 50 cells were screened). Correlation coefficients among the estimated values of DNA damage and tail moment for different groups were determined separately in each dietary status (Table 5). All these measured values appeared positive (<1) and therefore reflected the strong correlations between the observed values.
Nicotine-curcumin interaction is shown in Fig. 3A. The Binding constant of curcumin to nicotine was determined as 3.52x103 M-1 from the Binding-isotherm plot as shown in the inset of Fig. 3A. The Hill coefficient (n) for the nicotine-curcumin interaction was calculated as 1. Nicotine also effectively bound with DNA and imposed a conformational change on DNA because of which the characteristics absorbance peak suffered a shift (Fig. 3B). When nicotine concentration was increased, multiple absorbance peaks of DNA were appeared as seen from Fig. 3B.
| Table 4: | DNA damage and tail moment in the blood cells of female rats when animals were treated with nicotine and nicotine + curcumin (comet assay) |
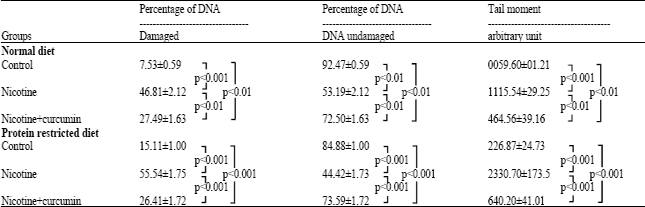 | |
| For comet assay, a layer of 200 μL of 2% agarose (mp. 40-42°C) was prepared on a pre-cooled fully frosted slide and covered with a cover slip. After carefully removing the cover slip, a second layer of 100 μL low melting agarose mixed with 20 μL of whole blood was pipetted out on the slide, covered again by cover slip and allowed to solidify at 4°C for 15 min. The slide (without cover slip) was immersed in a freshly prepared and chilled lysing solution (2.5 M NaCl, 100 mM EDTA, 10 mM Tris-HCl, 1% sarkosyl, 10% DMSO and 1% triton X-100, at pH 10, DMSO added just before use) and kept at 4°C for 2 h. Slides were then placed on a horizontal gel electrophoresis platform in alkaline electrophoresis solution (300 mM NaOH and 1 mM EDTA, pH 13) and left in dark for 15 min to allow unwinding of the DNA to occur. The DNA was then electrophoresed at 4°C in the dark for 15 min at 1 v/cm and ~ 250 mA. The slides were gently rinsed in neutralization buffer (0.4 M Tris-HCl, pH 7.5) and stained with 50 μL of 20 μg mL-1 ethidium bromide and covered with a cover slip. The photomicrograph of each slide was taken in Leica fluorescent microscope (Model 300 FX) at 40 x magnification. The comet head diameter, tail length, tail moment and percentage of DNA damage were measured. A total of 50 cells were screened per animal and percentage of DNA damages and tail moments were calculated. From four animals (n = 4) of each group, data were taken and averaged. Values are given Mean±SD. Significance levels were determined by ANOVA where p<0.01 was significant and p<0.001 was more significant | |
| Table 5: | Correlation coefficient of the DNA damage and tail moment of blood cells of female rats under normal and protein restricted diet |
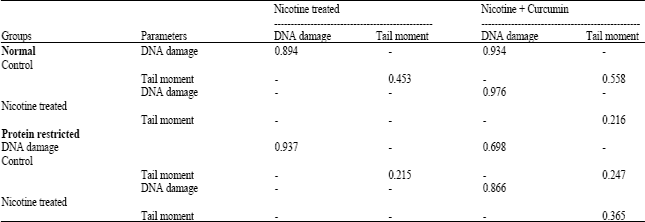 | |
| The percentages of DNA and tail moment of rat blood cells were calculated for each subgroup. To observe the significance of the measured data, correlation coefficients were determined for each dietary condition separately. The coefficients between the DNA damages of nicotine treated vs. control rats, nicotine + curcumin treated vs. control rats and also nicotine treated vs. nicotine + curcumin treated rats were determined. The coefficients between the tail moments of nicotine treated vs. control rats, nicotine + curcumin treated vs. control rats and also nicotine treated vs. nicotine + curcumin treated rats were determined. The coefficients were determined by CORREL function of Microsoft Excel | |
| Table 6: | Time kinetics of nicotine-BSA interaction |
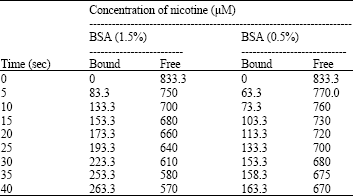 | |
| 833.3 μM of nicotine was added with two different concentrations of BSA separately, incubated at different intervals of time and absorbance of nicotine at 260 nm was recorded after each time interval. From these absorbance data, concentration of nicotine was calculated by using a pre-calibrated standard curve (nicotine concentration vs. absorbance at 260 nm) | |
Curcumin resists the nicotine-induced conformational change of DNA for which the characteristics absorbance peak of DNA at 260 nm reappeared (Fig. 3C). Curcumin failed to regain the characteristics absorbance peak of DNA when treated with 1 mM of nicotine.
The result for nicotine-BSA interaction is shown in Table 6. Nicotine interacts with BSA independently (Hill coefficient; n = 1). Result from Table 6 revealed that more unbound nicotine molecules were available in the solution for lesser concentrations of BSA present in the reaction mixture.
Normal DNA produced two negative peaks at 215, 245 nm and two positive peaks at 225, 275 nm (Fig. 4A) in CD spectrum. Nicotine treatment on DNA induced remarkable changes in the CD-spectra of DNA.
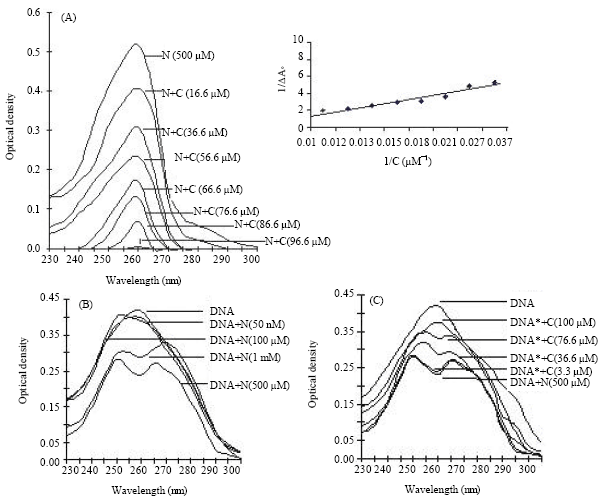 | |
| Fig. 3: | (A) Absorbance spectra of nicotine vs. curcumin interaction. Binding-isotherm plot of nicotine vs. curcumin interaction is shown in inset. (B) Absorbance spectra of DNA (30 μg) vs. nicotine interaction and (C) Absorbance spectra of nicotine treated DNA (30 μg) vs. curcumin interaction. N: Nicotine; C: Curcumin; DNA*: DNA+N(500 μm) |
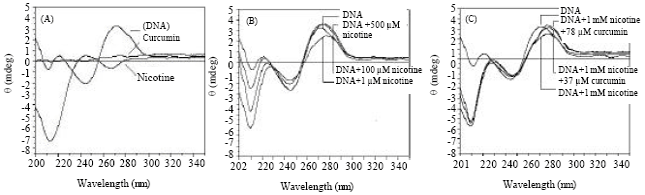 | |
| Fig. 4: | (A) CD spectra of control DNA (30 μg), nicotine (1 μM) and curcumin (100 μM). (B) CD spectra of DNA (30 μg) vs. nicotine interaction and (C) CD spectra of nicotine treated DNA (30 μg) vs. curcumin interaction |
With increasing concentrations of nicotine, the peaks of DNA at 215 and 225 nm kept suppressing which ultimately led to red-shift of the peak from 225 to 230 nm Fig. 4B). Addition of nicotine (up to 500 μM) initially caused a rise of the peaks at 245 and 275 nm and above those concentration of nicotine (1 mM) finally resulted in suppression of the DNA peak at 275 nm causing a red-shift to 280 nm (Fig. 4B). Curcumin was unable to alter the nicotine induced changes in the range 215 to 245 nm, but there was a significant change in 275 nm region (Fig. 4C). Addition of 78 μM curcumin could regain the peak height of DNA with slight recovery from the red-shift at 275 nm region and above this concentration of curcumin, there was no further change observed.
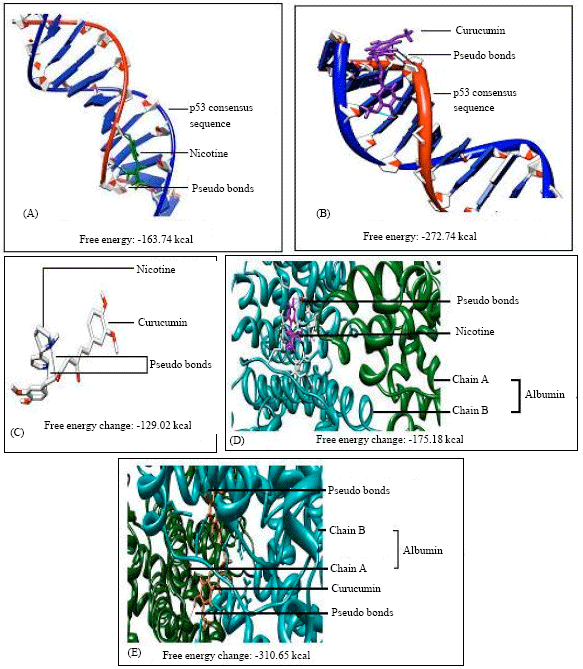 | |
| Fig. 5: | (A) DNA vs. nicotine docking. (B) DNA vs. curcumin docking. (C) Nicotine vs. curcumin docking. (D) Human serum albumin vs. nicotine docking and (E) Human serum albumin vs. curcumin docking |
DNA-nicotine and DNA-curcumin docking indicated that both drugs can bind to the thymidine 6 consensus sequence of p53 gene (GGGCATGCCTAGGCATGCC) (PDB ID 3KMD) of human with a free energy change of -163.74 and -272.75 kcal, respectively (Fig. 5A, B). Nicotine-curcumin docking showed a free energy change of -129.02 kcal and 2 pseudo-bond formation between C8 (carbon 8) and C18 of curcumin and 1C and 1N (1 nitrogen) of nicotine, respectively (Fig. 5C). Albumin-nicotine docking showed a free energy change of -175.18 kcal and 3 pseudo-bond formation between arginine 428 (chain B) of albumin and 1C, 1N and 1H (1 hydrogen) of nicotine (Fig. 5D). Albumin-curcumin docking showed a free energy change of -310.65 kcal, 1 hydrogen bond between tyrosine161 OH (hydroxyl ion) and curcumin O4 (Oxygen 4) and 9 pseudo-bond formations between leucine115 (chain B) and C3, H2 of curcumin, tyrosine161 (chain B) and H14, O4 of curcumin, arginine114 (chain B) and O5 of curcumin, alanine172 (chain A) and H13 of curcumin, asparagine173 (chain A) and O3 of curcumin (Fig. 5E).
DISCUSSION
India ranks third among the most important tobacco producing countries next to mainland China and USA and accounts for about 7.9% of the total tobacco production (National Institute of Occupational Health). In addition to cigarettes smoking, bidis (locally made smoking substance rolled by hand in tendu leaf, Diospyrus mebunoxylon or Diospyrus ebenum or white paper) are commonly smoked, as it is cheaper (Rahman and Fukui, 2000; Tseng, 2009) and widespread among socio economically handicapped population in developing countries. Use of all forms of tobacco is associated with higher all-cause mortality in the Indian populations (Gajalakshmi et al., 2003) and therefore the study is worth of investigation.
This study shows that nicotine reduces the RBC count by 26% in normal diet fed rat and 48.55% in protein restricted diets in comparison to their respective controls and curcumin minimizes the nicotine induced reduction of RBC count and increases the RBC counts (23% in normal diet and 64% in protein restricted diet fed rats). The reduction of RBC count by nicotine may be due to peroxidative membrane damages of erythrocytes via ROS as suggested by Tedesco et al. (2000). Furthermore, nicotine causes significant decrease in haemoglobin contents of the blood cells in both normal and protein restricted diet fed rats which supports the reduction of RBC counts. It has been already reported that binding of oxygen with haemoglobin is affected by carbon monoxide exerted from tobacco smoke due to its greater binding affinity (>200 times) than that of oxygen (Schwartzstein and Parker, 2005). In our experiment this may not be the issue; nicotine induced low RBC count may be responsible for decreased blood haemoglobin content and the compensatory effect of curcumin is due to the protective effect against RBC damage by nicotine.
We further report here that the average DNA yield per 300 μL of rat blood under normal dietary condition is 115.38 μg, in line with the value (105-158 μg) described earlier (Karissa et al., 2002). It has been reported that reduction of DNA content at different doses of nicotine (Abreu-Villac et al., 2003) is caused by either nicotine-induced suppression of cellular proliferation and/or an increase in DNA breakdown as a result of increased rate of cell death (Mohammed et al., 2004). Present results support the above mentioned studies because nicotine decreases significant DNA content of RBC under both normal- and protein restricted- diet fed rats. We have already reported that nicotine-induced toxicity in female rats is more in lipid profile (plasma) and lipid peroxidation (plasma and ovary tissue) under protein restricted diet as compared to that of the normal protein diet (Chattopadhyay and Chattopadhyay, 2008). Therefore it appears that the oxidative DNA damaging property of nicotine reduces the total DNA concentration in the blood cells and curcumin antagonize this process leading to increase DNA concentration. Protein malnutrition is a metabolic stress, which is aggravated by drug abuse and therefore severe toxic effect of nicotine in such condition is observed which is in agreement with the earlier findings (Angeli-Greaves and McLean, 1981).
The comet assay is a well-established genotoxicity test for in vitro and in vivo testing of chemicals (Brendler-Schwaab et al., 2005; Tice et al., 2000). We observed significant DNA damage in blood cells of normal diet fed rat and more pronounced damage in protein restricted diet fed rat by nicotine. This may be due to increased production of free radicals and Reactive Oxygen Species (ROS) that attack all types of macromolecules including DNA and damage it as reported in earlier studies (Yildiz et al., 1998; Comporti, 1989). Sudheer et al. (2007) have shown that nicotine could damage the DNA of rat peripheral blood lymphocytes in vitro through lipid peroxidation. Therefore high percentage of DNA damage and greater value of tail moment as obtained in our experiment suggest that nicotine aggravates DNA single strand break in protein restricted diet fed rat and reestablishes the nicotine induced DNA damage as seen previously. The high positive values (<1) of the correlation coefficients amongst the DNA damages or tail moment in both dietary conditions supports the significance of the data and also approves the validity of our experimental results. The photomicrographs of the comet assay (indicated color red) clearly indicate that nicotine plays a significant role for genotoxicity to blood cells of female rats, which is more prominent in protein restricted condition.
Curcumin has been widely studied for its antimutagenic and tumor-inhibiting properties (Ray et al., 2003). The protective effect of curcumin is due to its antioxidant action, modulation of mutagen metabolism by absorbing the xenobiotics (Premkumar et al., 2004) and inhibition of superactive oxygen species (SOS) functions (Oda, 1995). According to Basir (2004), curcumin resists the single strand breaks in DNA of yeast cells. These studies show that in addition to anti-initiating, detoxifying and antioxidant activities, curcumin has the ability to repair DNA damage also. The inhibition of nicotine induced DNA damage by curcumin observed by us further confirms the anti-initiating, detoxifying and antioxidant activities of curcumin. Xiaohong et al. (2000) have reported that nicotine affects the genes expression by adduct formation with DNA through its reaction with DNA-binding histones. Curcumin treatment causes a significant increase of histone hypoacetylation, which in turn regulates the gene transcription (Mukhopadhyay et al., 2002; Liu et al., 2009) by maintaining chromatin integrity. The regulatory role at transcriptional level and antioxidative role at cellular level of curcumin therefore justify the antagonistic role of curcumin against nicotine-induced effect in blood cells. Al-Suhaibani (2009) has seen the radioprotective effect of curcumin on γ-radiation-induced genetic toxicity in human blood lymphocytes which is again supplemented by out results. Further studies of curcumin during cellular proliferation and apoptosis may clear the exact role of it in protein-restricted condition.
In vitro interaction study clearly shows that nicotine binds to DNA and imposes a conformational change of it (Fig. 3B; 4B and 5A). The degree of damage is more for higher concentration of nicotine. Curcumin reduces the nicotine-induced genotoxic effect as revealed from Fig. 3C and 4C. In fact curcumin also binds to nicotine with a binding affinity 3.52x103 M-1 and quenches the absorbance of nicotine (Fig. 3A, 5C). The Hill coefficient for the nicotine-curcumin interaction is calculated as 1 which suggests that curcumin can bind to nicotine independently not cooperatively. Approximately 100 μM curcumin was sufficient to suppress the total absorbance intensity of 500 μM nicotine. It appears that nicotine binding to DNA is mainly on the surface of DNA, which increases the chiral nature of DNA molecule and enhances the CD peaks at 260-280 nm. When these sites becomes saturated for nicotine (>500 μM), the nicotine molecule probably inserts within the bases of DNA and perturb the DNA structure. This decreases the CD signal and increases UV absorbance of DNA (Fig. 4B, 3B). These results may help to understand the amelioration of curcumin against nicotine induced toxicity.
Wang et al. (2000) have shown that nicotine is capable of binding with BSA to form 1:1 complex and suggested that BSA plays a unique role in the detoxification and/or transportation of nicotine. In protein restriction condition, the concentration of unbound nicotine is increased as less protein is available for nicotine to bind (Table 6). In such condition more free nicotine molecules will be available to interact with DNA causing more damage as it is evident from our observations. The results of docking indicate that there are chances of nicotine and curcumin bonding on the same position (thymidine 6) with p53 (GGGCATGCCTAGGCATGCC) consensus sequence of human DNA (free energy changes -163.74 kcal for nicotine kcal and -272.75 kcal for curcumin, respectively) and even greater chances of nicotine and curcumin bonding with the specified residues of albumin (free energy changes-175.18 and -310.65 kcal, respectively). Docking results also indicate that binding of nicotine on albumin is more stable than its binding on DNA due to significant difference between the free energy changes. So nicotine causes greater damage on DNA in protein restricted condition. On the other hand, curcumin interacting with nicotine (free energy change -129 kcal) reduces the effect of nicotine on DNA in all conditions. Also curcumin has strong interaction with albumin (free energy change - 310.65 kcal), so in protein restricted condition, more unbound curcumin molecules will be available as compared to that of normal protein diet condition and hence will cause more amelioration of DNA against nicotine induced toxicity.
The metabolic stress occurred in protein restriction condition affects the immune systems. Nicotine thus induces severe damage at the cellular and genomic levels of living system under protein restricted condition. Smoking malnourished females may be at a risk to nicotine associated diseases like blood cancer and curcumin supplementation in their diet may benefit from nicotine induced blood cell damage.
ACKNOWLEDGMENTS
The supports for experimental facilities of Biophysics Laboratory (Physics Department, Jadavpur University) sponsored by Department of Biotechnology, New Delhi and also of UGC-DAE Consortium for Scientific Research, Sector 3, Salt lake, Kolkata, India, are gratefully acknowledged. Professor Soumen Basak, Head, Chemical Sciences Department, Saha Institute of Nuclear Physics, Kolkata and Dr. B.N Paul of Indian Institute of Toxicology Research, Lucknow were gratefully acknowledged for their kind help and advice. The authors are also thankful to Jadavpur University for giving legal approval of the work.
REFERENCES
- Abreu-Villac, Y., F.J. Seidler, C.A. Tate and T.A. Slotkin, 2003. Nicotine is a neurotoxin in the adolescent brain: Critical periods, patterns of exposure, regional selectivity and dose thresholds for macromolecular alterations. Brain Res., 979: 114-128.
PubMedDirect Link - Al-Suhaibani, E.S., 2009. Protective effect of curcumin on γ-radiation-induced sister chromatid exchanges in human blood lymphocytes. Int. J. Low Radiation, 6: 21-27.
CrossRefDirect Link - Bandyopadhyaya, G., S. Sinha, B.D. Chattopadhyay and A. Chakraborty, 2008. Protective role of curcumin against nicotine induced genotoxicity on rat liver under restricted dietary protein. Eur. J. Pharmacol., 588: 151-157.
CrossRef - Barcroft, J. and A.V. Hill, 1910. The nature of oxyhaemoglobin, with a note on its molecular weight. J. Physiol., 39: 411-428.
PubMedDirect Link - Mallo, P.R., G.R. Pinto and C. Botelho, 2001. The influence of smoking on fertility, pregnancy and lactation. J. Pediatr., 77: 257-264.
PubMedDirect Link - Bjartveit, K. and A. Tverda, 2005. Health consequences of smoking 1-4 cigarettes per day. Tobacco Control, 14: 315-320.
CrossRefDirect Link - Brendler-Schwaab, S., A. Hartmann, S. Pfuhler and G. Speit, 2005. The in vivo comet assay: Use and status in genotoxicity testing. Mutagenesis, 20: 245-254.
Direct Link - Chattopadhyay, K. and B.D. Chattopadhyay, 2008. Effect of nicotine on lipid profile, peroxidation and antioxidant enzymes in female rats with restricted dietary protein. Indian J. Med. Res., 127: 571-576.
PubMedDirect Link - Chun, K.S., Y.S. Keum, S.S. Han, Y.S. Song, S.H. Kim and Y.J. Surh, 2003. Curcumin inhibits phorbol esterinduced expression of cyclooxygenase-2 in mouse skin through suppression of extracellular signal regulated kinase activity and NF-kappa B activation. Carcinogenesis, 24: 1515-1524.
PubMedDirect Link - Chung, T.W. and C.P. Ho, 1999. Changes of viscosity of low shear rates and viscoelastic properties of oxidative erythrocyte suspensions. Clin. Hemorheol. Microcirc., 21: 99-103.
Direct Link - Comporti, M., 1989. Three models of free radical-induced cell injury. Chem. Biol. Interactions, 72: 1-56.
CrossRef - Cooke, J.P., 2007. Angiogenesis and the role of the endothelial nicotinic acetylcholine receptor. Life Sci., 80: 2347-2351.
CrossRef - Dani, J.A., D. Ji and F.M. Zhou, 2001. Synaptic plasticity and nicotine addiction. Neuron, 31: 349-352.
CrossRefDirect Link - El-Demerdash, F.M., M.I. Yousef and F.M.E. Radwan, 2009. Ameliorating effect of curcumin on sodium arsenite-induced oxidative damage and lipid peroxidation in different rat organs. Food Chem. Toxicol., 47: 249-254.
CrossRefDirect Link - Gajalakshmi, V., R. Peto, T.S. Kanaka and P. Jha, 2003. Smoking and mortality from tuberculosis and other diseases in India: Retrospective study of 43000 adult male deaths and 35000 controls. Lancet, 362: 507-515.
PubMedDirect Link - Helma, C. and M. Uhl, 2000. A public domain image-analysis program for the single-cell gel-electrophoresis (comet) assay. Mutat. Res., 466: 9-15.
CrossRefPubMedDirect Link - Kalpana, C., A.R. Sudheer, K.N. Rajasekharan and V.P. Menon, 2007. Comparative effects of curcumin and its synthetic analogue on tissue lipid peroxidation and antioxidant status during nicotine-induced toxicity. Singapore Med. J., 48: 124-130.
Direct Link - Karissa, K.A., A.S. Daniel, E.J. Thomas, R.S. Cara, P.O. Darin and M.H. Ellen, 2002. Utilizing genomic DNA purified from clotted blood samples for single nucleotide polymorphism genotyping. Arch. Pathol. Lab. Med., 126: 266-270.
PubMedDirect Link - Kido, R., I. Sato and S. Tsuda, 2006. Detection of in vivo DNA damage induced mice using the alkaline single cell gel electrophoresis (Comet) assay. J. Vet. Med. Sci., 68: 41-47.
Direct Link - Liu, Z., Z. Xie, W. Jones, R.E. Pavlovicz and S. Liu et al., 2009. Curcumin is a potent DNA hypomethylation agent. Bioorg. Med. Chem. Lett., 19: 706-709.
CrossRefPubMedDirect Link - Mandal, K., S. Dasgupta and B.D. Chattopadhyay, 2004. Nicotine induced alterations on reproductive organs of female rats under protein deficient condition. Indian J. Exp. Biol., 42: 330-332.
PubMedDirect Link - Akmal, M., A. Kesani, B. Anand, A. Singh, M. Wiseman and A. Goodship, 2004. Effect of nicotine on spinal disc cells: A cellular mechanism for disc degeneration. Spine, 29: 568-575.
PubMedDirect Link - Mukhopadhyay, A., S. Banerjee, L.J. Stafford, C. Xia, M. Liu and B.B. Aggarwal, 2002. Curcumin-induced suppression of cell proliferation correlates with down-regulation of cyclin D1 expression and CDK4-mediated retinoblastoma protein phosphorylation. Oncogene, 21: 8852-8861.
PubMedDirect Link - Oda, Y., 1995. Inhibitory effect of curcumin on SOS functions induced by UV irradiation. Mutat. Res. Lett., 348: 67-73.
CrossRefDirect Link - Premkumar, K., S. Kavitha, S.T. Santhiya, A.R. Ramesh and J. Suwanteerangkul, 2004. Interactive effects of saffron with garlic and curcumin against cyclophosphamide induced genotoxicity in mice. Asia Pacific J. Clin. Nutr., 13: 292-294.
PubMedDirect Link - Prescott, E., M. Hippe, P. Schnorr, H.O. Hein and L. Vestbo, 1998. Smoking and risk of myocardial infarction in women and men: Longitudinal population study. Br. Med. J., 316: 1043-1047.
Direct Link - Rahman, M. and T. Fukui, 2000. Bidi smoking and health. Public Health, 114: 123-127.
PubMedDirect Link - Ray, S., N. Chattopadhyay, A. Mitra, M. Siddiqi and A. Chatterjee, 2003. Curcumin exhibits antimetastatic properties by modulating integrin receptors, collagenase activity and expression of Nm23 and E-cadherin. J. Environ. Pathol. Toxicol. Oncol., 22: 49-58.
PubMedDirect Link - Sasikala, K., R.F. Rosalin, A.L.C. Jude, A.R. Kumar and S. Sudha et al., 2003. Active and passive smokers-a haematobiochemical and cytogenetic study. Int. J. Hum. Genet., 3: 29-32.
Direct Link - Sela, S., R. Shurtz-Swirski, J. Awad, G. Shapiro, L. Nasser, S.M. Shasha and B. Kristal, 2002. The involvement of peripheral polymorphonuclear leucocytes in the oxidative stress and inflammation among cigarette smokers. Isr. Med. Assoc. J., 4: 1015-1019.
PubMedDirect Link - Sudheer, A.R., S. Muthukumaran, C. Kalpana, M. Srinivasan and V.P. Menon, 2007. Protective effect of ferulic acid on nicotine-induced DNA damage and cellular changes in cultured rat peripheral blood lymphocytes: A comparison with N-acetylcysteine. Toxicol. In Vitro, 21: 576-585.
CrossRef - Tedesco, I., M. Russo, P. Russo, G. Iacomino and G.L. Russo et al., 2000. Antioxidant effect of red wine polyphenols on red blood cells. J. Nutr. Biochem., 11: 114-119.
CrossRefPubMedDirect Link - Tice, R.R., E. Agurell, D. Anderson, B. Burlinson and A. Hartmann et al., 2000. Single cell gel/comet assay: Guidelines for in vitro and in vivo genetic toxicology testing. Environ. Mol. Mutagen., 35: 206-221.
CrossRefPubMedDirect Link - Wang, Y., Y. Cheng and H.F. Sun, 2000. Interaction of nicotine and bovine serum albumin. Chinese Chem. Lett., 11: 247-250.
Direct Link - Xiaohong, W., S. Hongfang and L. Yuanfang, 2000. The conformation alteration of mouse hepatic histones after reacting with nicotine in vitro. Chinese Sci. Bull., 45: 825-830.
CrossRefDirect Link - Yildiz, D., N. Ercal and D.W. Armstrong, 1998. Nicotine enantiomers and oxidative stress. Toxicology, 130: 155-165.
CrossRef - Keloglu-Isler, E. and I. Erdogan, 2007. Opinions of Turkish university students on cigarette smoking at schools. J. Applied Sci., 7: 2762-2768.
CrossRefDirect Link - Tseng, P.J., 2009. Human exposure to perfluorinated compounds via smoking and second-hand smoke. J. Applied Sci., 9: 2475-2479.
CrossRefDirect Link