
H.H. Azzaz
Department of Dairy Sciences, National Research Centre, 33 Buhouth St, Dokki, Giza, Egypt
H.M. Ebeid
Department of Dairy Sciences, National Research Centre, 33 Buhouth St, Dokki, Giza, Egypt
T.A. Morsy
Department of Dairy Sciences, National Research Centre, 33 Buhouth St, Dokki, Giza, Egypt
S.M. Kholif
Department of Dairy Sciences, National Research Centre, 33 Buhouth St, Dokki, Giza, Egypt
International Journal of Dairy Science
Year: 2015 | Volume: 10 | Issue: 3 | Page No.: 107-116







ABSTRACT
This study was conducted to study the effect of yeast culture (Saccharomyces cerevisiae) supplementation either alone or in combination with Propionibacterium freudenreichii strain P169 on nutrient digestibility coefficients, blood metabolites, milk yield and milk composition of mild lactating buffaloes. Fifteen lactating buffaloes, 2 months after parturition, were randomly assigned into three groups, 5 animals each, using complete random design. The experiment lasted for two months, buffaloes were fed dry matter according to 3% of their mean body weight. The experimental groups were fed on: (1) Control ration (consisted of 50% concentrate feed mixture (CFM), 30% corn silage, 10% dried sugar beet pulp and 10% rice straw), (2) Control ration+50 g Yeast Culture (YC)/head/day and (3) Control ration+50 g YC+4 g propionibacteria, P169, (YC+P169)/head/day. The supplementation of YC or YC+P169 improved (p<0.05) all nutrients digestibility but significantly decreased (p<0.05) blood plasma urea nitrogen of treated buffaloes. Milk and 4% fat corrected milk yields were significantly increased (p<0.05) while milk fatty acids were unaffected by YC or YC+P169 supplementation. In conclusion, ration’s supplementation with YC or YC+P169 had beneficial effects on the buffaloes productivity with no deleterious effects on animals health.
PDF Abstract XML References Citation
Received: April 19, 2015;
Accepted: May 20, 2015;
Published: June 29, 2015
How to cite this article
H.H. Azzaz, H.M. Ebeid, T.A. Morsy and S.M. Kholif, 2015. Impact of Feeding Yeast Culture or Yeast Culture and Propionibacteria 169 on the Productive Performance of Lactating Buffaloes. International Journal of Dairy Science, 10: 107-116.
DOI: 10.3923/ijds.2015.107.116
URL: https://scialert.net/abstract/?doi=ijds.2015.107.116
DOI: 10.3923/ijds.2015.107.116
URL: https://scialert.net/abstract/?doi=ijds.2015.107.116
INTRODUCTION
Use of probiotics as feed additives for ruminants has been largely studied in the last two decades. Empirical observations suggest that incorporation of some types of live microorganisms such as propionibacteria and yeasts in feeds may beneficially influence animal performance in many types of production systems (Dawson, 2002).
Propionibacteria represent 1.4% of rumen total microbial population and propionate which was produced is a major precursor for glucose production through gluconeogenesis, spares glucogenic amino acids in hepatic gluconeogenesis, inhibits hepatic lipid oxidation and work as antiketogenic agent (Morsy et al., 2014). Effects of feeding each of propionibacteria, yeast culture or their combination to dairy animals have been evaluated but results were inconsistent. Stein et al. (2006) found that early lactating cows fed 6×1010 or 6×1011 CFU day-1 of Propionibacterium strain P169 produced about 8% more fat corrected milk than did the control cows, while, Morsy et al. (2014) found that supplementing dairy buffalo’s rations with Propionibacterium strain P169 did not affect nutrients digestibility, milk yield or milk components. Moreover, yeast based products have been shown to affect feed intake, rumen pH, nutrient digestibility and milk production of dairy animals in some studies (Campanile et al., 2008; Wang et al., 2009), but, in the others (Arambel and Kent,1990; Soder and Holden, 1999) yeast and yeast products did not cause any beneficial responses.
The effects of yeast culture and propionibacteria supplementation might differ, also the combination of these two feed additives might have an advantage over the use of each alone. Therefore, this study was conducted to study the effect of yeast culture (Saccharomyces cerevisiae) supplementation either alone or in combination with Propionibacterium freudenreichii strain P169 on nutrient digestibility coefficients, blood metabolites, milk yield and milk composition of mild lactating buffaloes.
MATERIALS AND METHODS
This study was carried out at the Agricultural Experimental Station, Cattle Research Unit, Faculty of Agriculture, Cairo University, Giza, Egypt. Laboratory analyses were carried out at the laboratories of Diary Sciences Department, National Research Centre, Dokki, Giza, Egypt.
Yeast and propionibacteria sources: Dairy ProP169® (a freeze-dried material of viable 3×1010 colony forming unit (CFU g-1) of Propionibacterium freudenreichii strain P169, Bio-Vet Inc. USA) and viable 2.6×104 CFU g-1 culture of yeast Saccharomyces cerevisiae (which maintained on Malt agar medium and obtained from Microbial Chemistry Lab., National Research Centre, Dokki, Giza, Egypt) were used as feed supplements in this study.
Feeding and management: Fifteen lactating buffaloes, after 2 months of calving, aged 4-6 years and weighting 615 kg on average were randomly assigned into three groups, 5 animals each, using complete random design. The entire experimental period was 2 months. Buffaloes were fed dry matter according to 3% of their mean body weight. The experimental groups were fed on: (1) Control ration (consisted of 50% concentrate feed mixture (CFM), 30% corn silage, 10% dried sugar beet pulp and 10% rice straw), (2) Control ration+50 g Yeast Culture (YC)/head/day and (3) Control ration+50 g YC+4 g Propionibacterium freudenreichii strain P169 (YC+P169)/head/day. The concentrate feed mixture was offered twice daily at 8.00 am and 8:00 pm, while corn silage and dried sugar beet pulp were offered at 9.00 and 11.00 am, respectively and rice straw was offered at 3:00 pm. Yeast culture or yeast culture+P169 were mixed with small amount of morning CFM and introduced to the animals once per day. Fresh water was available all the time for all experimental groups. The chemical composition of feed ingredients is shown in Table 1.
| Table 1: | Chemical composition (on DM basis %) of feed ingredients |
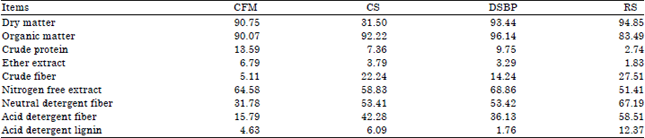 | |
| CFM: Concentrate feed mixture consisted 60% yellow corn, 15% wheat bran, 10% soybean meal ,10% distillers dried grains with soluble (DDGS) 3% limestone,1% minerals and1% of NaCl, CS: Corn silage, DSBP: Dried sugar beet pulp, RS: Rice straw and CTMR: Calculated total mixed ration | |
Apparent digestibility: At the end of each month of the experimental period, feces were collected from all animals (during the final two days) to determine the digestibility coefficients, where by silica was used as an internal marker. Fecal samples were collected from the rectum of each animal by hand at 12:00 pm (4 h after the distribution of morning CFM). The collected feces were dried at 60°C for 48 h and then ground for chemical analysis. Dry matter excreted in feces was calculated by dividing silica input in the feeds by silica output in the feces as described by Ferret et al. (1999). The nutrients digestibility coefficients were calculated according to the following equation (Ferret et al., 1999):
Feed and fecal analysis: Chemical analysis of feed stuffs and fecal samples were carried out to determine the percentage of Dry Matter (DM), Crude Protein (CP), Ether Extract (EE), Crude Fiber (CF) and ash content using the methods of the AOAC. (1995). The Nitrogen Free Extract (NFE) was calculated by difference. The Neutral Detergent Fiber (NDF), Acid Detergent Fiber (ADF) and Acid Detergent Lignin (ADL) contents were determined using the methods described by Van Soest et al. (1991).
Sampling and analysis of blood plasma: Blood samples were collected in glass tubes containing EDTA as an anticoagulant agent from the jugular vein of each animal at the last day of each month at 12:00 pm (4 h after the distribution of morning CFM) and then centrifuged at 4000 rpm/20 min to separate the plasma. The obtained plasma was stored at -18°C till analysis. Plasma total protein and albumin were determined as described by Armstrong and Carr (1964) and Doumas et al. (1971), respectively. Then, globulin and albumin/globulin ratio were calculated. Urea, glucose, Aspartate Aminotransferase (AST) and Alanin Aminotransferase (ALT) were determined as described by Fawcett and Soctt (1960) and Reitman and Frankel (1957), respectively.
Sampling and analysis of milk: Milk samples were taken every 2 weeks throughout the experimental period. Buffaloes were machinery milked twice a day at 7:00 am and 7:00 pm. Milk samples were collected immediately from each animal after morning and evening milking and milk yield was recorded. The sample of each animal represented a mixed sample of constant percentage of the evening and morning yield. Milk samples were analyzed for total solids, fat, total protein and lactose by Bentley150 infrared milk analyzer (Bentley Instruments, Chaska, MN, USA) according to AOAC (1995) procedures. Solids-Not-Fat (SNF) was calculated. Fatty acids profile of milk fat was determined as methylated according to Park et al. (2002) and separated by gas liquid chromatography. Fat corrected milk (4% fat) was calculated by using the following equation according to Azzaz et al. (2013):
Statistical analysis: Data obtained from this study were statistically analyzed by IBM SPSS (2008) according to the following model:
where, Yij is the parameter under analysis of the ij buffalo, μ is the overall mean, Ti is the effect due to treatment on the parameter under analysis and eij is the experimental error for ij on the observation. Duncan’s multiple range tests were used to test the significance among means (Duncan, 1955).
RESULTS
Digestibility and nutritive values: Data of Table 2 clearly showed significant (p<0.05) increase of all nutrients digestibility coefficients by buffaloes fed rations supplemented with YC or YC+P169 compared with those fed the control ration. No significant (p>0.05) differences were detected in all nutrients digestibility coefficients among YC or YC+P169 treated buffaloes. Also, the nutritive values of the experimental rations expressed as Total Digestible Nutrients (TDN) and Digestible Crude Protein (DCP) take the same trend of nutrients digestibility coefficients.
Blood plasma parameters: The effect of YC or YC+P169 supplementation on the metabolic blood profile of lactating buffaloes is presented in Table 3. There were insignificant differences (p>0.05) among all groups in the overall means of plasma ALT, AST, total lipids, total protein, albumin, globulin concentration and albumin/globulin ratio. Also, there were insignificant differences (p>0.05) among all groups in the overall means of plasma glucose concentration, but plasma glucose showed slight increase by buffaloes fed rations supplemented with YC or YC+P169 compared with buffaloes of the control group.
| Table 2: | Apparent nutrients digestibility coefficients and nutritive values of the experimental rations |
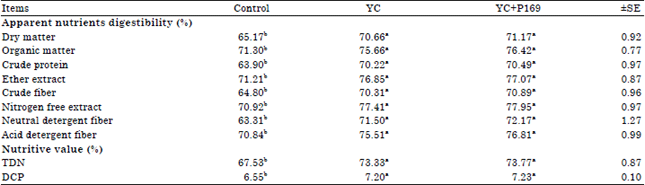 | |
| TDN: Total digestible nutrients, DCP: Digestible crude protein, a and b means at the same row with different superscript are significantly different at (p<0.05), YC: Control ration+50 g yeast, YC+P169: Control ration+50 g yeast+4 g Propionibacterium freudenreichii strain P169, ±SE: Standard error | |
| Table 3: | Blood plasma parameters of lactating buffaloes fed the different experimental rations |
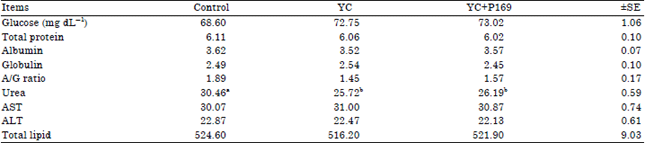 | |
| A/G: Albumin/Globulin ratio, AST: Aspartate aminotransferase, ALT: Alanin aminotransferase, a and b means at the same row with different superscript are significantly different at (p<0.05), YC: Control ration+50 g yeast, YC+P169: Control ration+50 g yeast+4 g Propionibacterium freudenreichii strain P169, ±SE: Standard error | |
| Table 4: | Milk yield and milk composition of lactating buffaloes fed the different experimental rations |
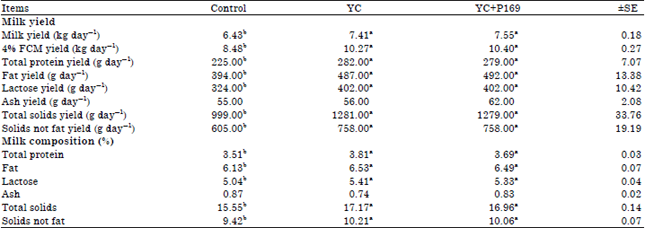 | |
| 4% FCM: 4% Fat corrected milk a and b means at the same row with different superscript are significantly (p<0.05) different, YC: Control ration+50 g yeast, YC+P169: Control ration+50 g yeast+4 g Propionibacterium freudenreichii strain P169, ±SE: Standard error | |
On the other hand, there were significant decrease (p<0.05) in plasma urea concentrations of YC and YC+P169 treated animals compared to those of the control group, while no significant (p>0.05) differences were detected in plasma urea concentrations among YC and YC+P169 treated buffaloes. Generally, plasma metabolites are frequently used to monitor the metabolic health status of dairy animals. In our study, the physiological limits of all blood parameters were within the normal range for healthy animals.
Milk yield and milk composition: Milk yield and 4% Fat Corrected Milk (FCM) yield were significantly higher (p<0.05) for YC or YC+P169 treated groups compared to the control group, but there were no significant differences among YC and YC+P169 treated groups (Table 4). Also, the percentages and yields of milk fat, protein, lactose, Total Solids (TS) and Solid Not Fat (SNF) take the same trend of milk productivity.
Milk fatty acids profile: The effects of YC and YC+P169 supplementation on milk methylated fatty acids are shown in Table 5. Yeast culture or YC+P169 supplementation had no effect on milk fatty acids profile of treated buffaloes compared to the control.
DISCUSSION
In the present study, the improvement in apparent digestibility coefficients of YC or YC+P169 treated buffaloes are consistent with the results obtained by Lehloenya et al. (2008a) who reported that total tract digestibility of OM, NDF and ADF tended to increase when YC or P169 were fed to steers. Moreover, Gaafar et al. (2009) demonstrated that digestibility coefficients of all nutrients and nutritive values of experimental rations increased significantly by buffaloes treated with baker’s yeast. These positive effects of treatments may be attributed to the effects of yeast culture in the rumen environment including the rise of ruminal pH, which stimulate proteolytic bacteria causing CP digestion to increase (Williams et al., 1991). It also, increase of microbial protein synthesis apart from altering the amino acid profile of the duodenal digesta (Erasmus et al., 1992). As well as, reduces lactate accumulation and reduce oxygen concentration in rumen fluid while improve utilization of starch supplied in the feeding ration (Girard, 1997). Moreover, decrease methane production, increase total number of microorganism’s especially cellulolytic bacteria and increase rate or extent of ruminal fiber digestion (Callaway and Martin, 1997). Additionally, rations composition may have contributed to the responses registered to YC supplementation.
| Table 5: | Milk fatty acids profile of lactating buffaloes fed the different experimental rations |
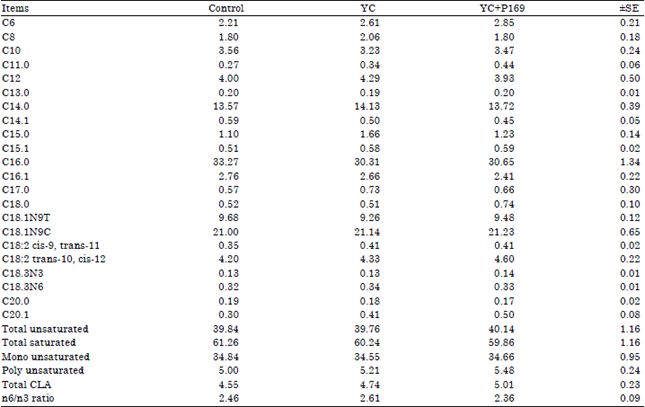 | |
| YC: Control ration+50 g yeast, YC+P169: Control ration+50 g yeast+4 g Propionibacterium freudenreichii strain P169, ±SE: Standard error | |
In our experiment, rations had high contents of corn silage and beet pulp, which are quickly fermented in the rumen causing an increase in acidity of the rumen environment. Therefore, this composition may improve the potential response to yeast supplementation (Williams et al., 1991).
The buffalo’s blood plasma glucose, ALT, AST, total lipids, total protein, albumin, globulin concentrations and albumin/globulin ratio were similar between all groups in the currant study. These finding had confirmed the results of Yalcin et al. (2011) who reported that cow’s blood plasma glucose, ALT, AST, cholesterol, triglyceride, total protein and albumin concentrations were not affected by yeast culture supplementation, also (Morsy et al., 2014) observed that buffalo’s blood plasma glucose and total lipids concentrations were not affected by propionibacteria P169 supplementation. Moreover, Lehloenya et al. (2008a, b) found that feeding P169 or P169+yeast did not affect plasma glucose concentrations in steers and multiparous Holstein cows. Lehloenya et al. (2008b) reviewed that lack of a detectable change in plasma glucose observed in dairy cows fed rations treated only with P169 or P169+yeast may be due partly to the fact that plasma insulin concentrations increased faster in response to treatment than the control. On the other hand, the significant lower of plasma urea concentrations of YC and YC+P169 treated animals may be attributed to the improvements occurred in metabolic process as a response to the YC or YC+P169 supplementation which reflect better utilization of protein in treated groups compared to the control group. Our results are in line with those obtained by Bruno et al. (2009) who reported that feeding YC reduced concentrations of urea nitrogen in blood plasma of dairy Holstein cows.
The increased synthesis of milk and 4% FCM yield of treated animals (YC or YC+P169) agreed with Lehloenya et al. (2008b) who found that feeding P169+yeast to cows increased actual milk and 4% FCM yield by 8.5-16.6% above the control cows and this overall increase in milk yield was due to increased milk production during mid lactation (9-30 weeks). Stimulatory factors for rumen bacteria such as Vitamins B, amino acids and organic acids are present in yeast culture and their absence may have contributed to the lack of milk production response as reported by Francisco et al. (2002). Also, propionibacteria likely increases milk production because of an increased supply of glucogenic precursors caused by changes in rumen fermentation (Stein et al., 2006).
Moreover, our results are concordant with those of Alshaikh et al. (2002) and Gaafar et al. (2009). Where they found that the percentages and yields of milk fat, protein, lactose, Total Solids (TS) and Solid Not Fat (SNF) were significantly higher in cows fed diets supplemented with YC or baker’s yeast, respectively, than those of the control. In addition, it was observed that both high and low dose of P169 tended to increase milk protein and SNF percentage of treated cows above control during 25 weeks study (Stein et al., 2006). However, Lehloenya et al. (2008b) stated that cows fed P169+yeast had significantly greater milk lactose and SNF percentage than cows of control or cows fed on yeast alone, while milk protein and fat percentage in P169+yeast and yeast groups did not differ.
The milk fatty acids from C4:0-C14:0, as well as about 50% of C16, arise from de novo synthesis within the mammary gland. In contrast, the longer chain fatty acids are supplied from circulating lipids and arise from either dietary sources or from depot lipids (Yalcin et al., 2011). Although, there were enhancements in milk fat percentage and yield of treated buffaloes, the YC or YC+P169 supplementation had no effect on milk fatty acids profile. Similarly, Yalcin et al. (2011) observed that milk fatty acids were unaffected by yeast culture supplementation. Also, Morsy et al. (2014) found that no significant effect of P169 supplementation at low or high dose on buffalo’s total milk fatty acid classes (saturated fatty acids, unsaturated fatty acids, monosaturated fatty acids, polyunsaturated fatty acids, n-6 polyunsaturated fatty acids, n-3 polyunsaturated fatty acids and conjugated linoleic acid). In our study, ration supplementation with YC or YC+P169 increased dry matter and organic matter digestibility (Table 2) thus allowing higher energy availability for milk production in treated buffaloes. The enhancement of milk fat percentage and yield might be attributed to increased milk production, improved degradation of structural carbohydrates and increased production of acetic acid in the rumen of YC or YC+P169 treated buffaloes.
Increased percentage and yield of milk protein might indicate that changes in rumen fermentation as a result of feeding YC or YC+P169 increased supply of glucogenic and aminogenic substrates. Moreover, Saccharomyces cerevisiae supplementation had been associated with increased flow of microbial protein leaving the rumen and enhanced supply of amino acids entering the small intestine (Paryad and Rashidi, 2009). Yalcin et al. (2011) suggested that the mammary gland has the capacity to alter the uptake of substrates from the arterial supply in response to changes in arterial amino acid concentrations, mammary blood flow and metabolic activity to improve milk protein yield.
The elevating of percentage and yield of milk lactose in YC or YC+P169 treated buffaloes could be attributed to the improvement in feed digestibility, specifically OM digestibility which may cause a numerical downward shift in the ratio of acetate to propionate and thereby increased delivery of glucogenic precursors to the mammary gland. Stein et al. (2006) thought that P169 supplementation with increased ruminal propionate should increase blood glucose concentration via gluconeogenesis and subsequently increase milk lactose. Increased percentage and yield of milk TS and SNF in treated buffaloes were likely due to increased milk components (fat, protein and lactose) that had been used in their calculation.
CONCLUSION
It can be concluded that lactating buffalo’s rations supplemented with yeast (Saccharomyces cerevisiae) either alone or in combination with Propionibacterium freudenreichii strain P169 showed significant improvement in nutrients digestibility, milk production and milk composition, but their supplementation had no effect on milk fatty acids profile. Also, no deleterious effects on general health of the treated animals were detected.
ACKNOWLEDGMENTS
The authors express appreciation to Bio-Vet, Inc (Blue Mounds, WI), especially of gratefully acknowledge for Mr. Bill Helwig, Marketing Director of Bio-Vet, Inc for helping and for providing the bacterial inoculants for carrying out this study.
REFERENCES
- AOAC., 1995. Official Methods of Analysis. 16th Edn., Association of Official Analytical Chemists, Washington, DC., USA.
Direct Link - Alshaikh, M.A., M.Y. Alsiadi, S.M. Zahran, H.H. Mogawer and T.A. Alshowime, 2002. Effect of feeding yeast culture from different sources on the performance of lactating Holstein cows in Saudi Arabia. Asian Aust. J. Anim. Sci., 15: 352-356.
Direct Link - Arambel, M.J. and B.A. Kent, 1990. Effect of yeast culture on nutrient digestibility and milk yield response in early- to midlactation dairy cows. J. Dairy Sci., 73: 1560-1563.
CrossRefDirect Link - Bruno R.G.S., H.M. Rutigliano, R.L. Cerri, P.H. Robinson and J.E.P. Santos, 2009. Effect of feeding Saccharomyces cerevisiae on performance of dairy cows during summer heat stress. Anim. Feed Sci. Technol., 150: 175-186.
CrossRefDirect Link - Callaway, E.S. and S.A. Martin, 1997. Effects of a Saccharomyces cerevisiae culture on ruminal bacteria that utilize lactate and digest cellulose. J. Dairy Sci., 80: 2035-2044.
CrossRefPubMedDirect Link - Campanile, G., F. Zicarelli, D. Vecchio, C. Pacelli and G. Neglia et al., 2008. Effects of Saccharomyces cerevisiae on in vivo organic matter digestibility and milk yield in buffalo cows. Livest. Sci., 114: 358-361.
Direct Link - Doumas, B.T., W.A. Watson and H.G. Biggs, 1971. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta, 31: 87-96.
CrossRefPubMedDirect Link - Erasmus, L.J., P.M. Botha and A. Kistner, 1992. Effect of yeast culture supplement on production, rumen fermentation and duodenal nitrogen flow in dairy cows. J. Dairy Sci., 75: 3056-3065.
CrossRefDirect Link - Patton, C.J. and S.R. Crouch, 1977. Spectrophotometric and kinetics investigation of the Berthelot reaction for the determination of ammonia. Anal. Chem., 49: 464-469.
CrossRefDirect Link - Ferret, A., J. Plaixats, G. Caja, J. Gasa and P. Prio, 1999. Using markers to estimate apparent dry matter digestibility, faecal output and dry matter intake in dairy ewes fed Italian ryegrass hay or alfalfa hay. Small Rumin. Res., 33: 145-152.
CrossRefDirect Link - Francisco, C.C., C.S. Chamberlain, D.N. Waldner, R.P. Wettemann and L.J. Spicer, 2002. Propionibacteria fed to dairy cows: Effects on energy balance, plasma metabolites and hormones and reproduction. J. Dairy Sci., 85: 1738-1751.
CrossRefDirect Link - Gaafar, H.M.A., A.M.A. Mohi El-Din, M.I. Basiuoni and K.F.A. El-Riedy, 2009. Effect of concentrate to roughage ratio and baker's yeast supplementation during hot season on performance of lactating buffaloes. Slovak J. Anim. Sci., 42: 188-195.
Direct Link - Lehloenya, K.V., C.R. Krehbiel, K.J. Mertz, T.G. Rehberger and L.J. Spicer, 2008. Effects of propionibacteria and yeast culture fed to steers on nutrient intake and site and extent of digestion. J. Dairy Sci., 91: 653-662.
CrossRefDirect Link - Lehloenya, K.V., D.R. Stein, D.T. Allen, G.E. Selk and D.A. Jones et al., 2008. Effects of feeding yeast and propionibacteria to dairy cows on milk yield and components and reproduction. J. Anim. Physiol. Anim. Nutr., 92: 190-202.
CrossRefDirect Link - Morsy, T.A., H.M. Ebeid, A.E.M. Kholif, H.A. Murad, A.E.M. Abd El-Gawad and T.M. El-Bedawy, 2014. Influence of propionibacteria supplementation to rations on intake, milk yield, compositionand plasma metabolites of lactating buffaloes during early lactation. Sci. Int., 2: 13-19.
Direct Link - Park, S.J., C.W. Park, S.J. Kim, J.K. Kim and Y.R. Kim et al., 2002. Methylation methods for the quantitative analysis of Conjugated Linoleic Acid (CLA) isomers in various lipid samples. J. Agric. Food Chem., 50: 989-996.
CrossRefDirect Link - Paryad, A. and M. Rashidi, 2009. Effect of yeast (Saccharomyces cerevisiae) on apparent digestibility and nitrogen retention of tomato pomace in sheep. Pak. J. Nutr., 8: 273-278.
CrossRefDirect Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Soder, K.J. and L.A. Holden, 1999. Dry matter intake and milk yield and composition of cows fed yeast prepartum and postpartum. J. Dairy Sci., 82: 605-610.
CrossRefPubMedDirect Link - Stein, D.R., D.T. Allen, E.B. Perry, J.C. Bruner and K.W. Gates et al., 2006. Effects of feeding propionibacteria to dairy cows on milk yield, milk components and reproduction. J. Dairy Sci., 89: 111-125.
CrossRefPubMedDirect Link - van Soest, P.J., J.B. Robertson and B.A. Lewis, 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci., 74: 3583-3597.
CrossRefPubMedDirect Link - Williams, P.E., C.A. Tait, G.M. Innes and C.J. Newbold, 1991. Effects of the inclusion of yeast culture (Saccharomyces cerevisiae plus growth medium) in the diet of dairy cows on milk yield and forage degradation and fermentation patterns in the rumen of steers. J. Anim. Sci., 69: 3016-3026.
PubMedDirect Link - Yalcin, S., S. Yalcin, P. Can, A.O. Gurdal, C. Bagci and O. Eltan, 2011. The nutritive value of live yeast culture (Saccharomyces cerevisiae) and its effect on milk yield, milk composition and some blood parameters of dairy cows. Asian-Aust. J. Anim. Sci., 24: 1377-1385.
Direct Link - Azzaz, H.H., H.A. Murad, A.M. Kholif, T.A. Morsy, A.M. Mansour and H.M. El-Sayed, 2013. Increasing nutrients bioavailability by using fibrolytic enzymes in dairy buffaloes feeding. J. Biol. Sci., 13: 234-241.
CrossRefDirect Link