
Intisar Islam
Little Rock Central High School, 1500 Park Street, Little Rock, AR 72204, USA
A.U. Shaikh
Department of Chemistry, University of Arkansas at Little Rock, Little Rock, AR 72202, USA
I.M. Shahidul
University of Arkansas at Pine Bluff, Mail Slot 4913, 153 Woodward Hall, 1200 North University Drive, Pine Bluff, AR 71601, USA
International Journal of Cancer Research
Year: 2009 | Volume: 5 | Issue: 3 | Page No.: 83-94







ABSTRACT
The present study investigated the physiological functions of sweetpotato leaves and as a resource for products with these functions. The polyphenolic compositions and the potential chemo-preventative properties such as radical scavenging activity and antimutagenicity of leaf extracts were compared in six selected sweetpotato (Ipomoea batatas L.) genotypes. The total phenolic content ranged from 12.18 to 16.17 g/100 g Dry Weight (DW). The Radical Scavenging Activity (RSA) ranged from 1.09 to 1.85 μmol Trolox mg-1. In the case of RSA, the cultivar V-5 showed highest activity (1.85 μmole Trolox equivalence mg-1 DM) and V-3 was the lowest (1.09 μmol Trolox equivalence mg-1 DM). The significant positive correlation between the radical scavenging activity and the level of total phenolic (r = 0.62; n = 90; p<0.01) suggests that phenolic compounds are important antioxidant components of sweet potato leaves. The antimutagenicity of the six sweetpotato genotypes studied ranges from 77 to 96% inhibition. The polyphenolic compositions of sweetpotato leaves were identified by using the Reverse Phase-High Performance Liquid Chromatography (RP-HPLC). All the RP-HPLC profiles of the genotypes tested showed peaks at the same retention times but peak areas of individual phenolic compounds differed. Six caffeoylquinic acid derivatives of sweetpotato leaves were identified and quantified. They include: caffeic acid, chlorogenic acid, 4,5-di-O-caffeoylquinic acid, 3,5-di-O-caffeoylquinic acid, 3,4-di-O-caffeoylquinic acid and 3,4,5-tri-O-caffeoylquinic acid. The caffeoylquinic acid derivatives showed higher DPPH-radical scavenging activity (%) and antimutagenicity by effectively inhibiting the reverse mutation induced by Trp-P-1 on Salmonella typhimurium TA 98. Thus, these phenolic components have potential value as chemo-preventative materials for human health.
PDF Abstract XML References Citation
How to cite this article
Intisar Islam, A.U. Shaikh and I.M. Shahidul, 2009. Antioxidative and Antimutagenic Potentials of Phytochemicals from Ipomoea batatas (L.) Lam. International Journal of Cancer Research, 5: 83-94.
DOI: 10.3923/ijcr.2009.83.94
URL: https://scialert.net/abstract/?doi=ijcr.2009.83.94
DOI: 10.3923/ijcr.2009.83.94
URL: https://scialert.net/abstract/?doi=ijcr.2009.83.94
INTRODUCTION
Sweet potato (Ipomoea batatas (L.) Lam.) is an important food crop. The consumption of sweetpotato tops as a fresh vegetable in many parts of the world (Villareal et al., 1982; Nwinyi, 1993; Yoshimoto et al., 2007) indicates that they are acceptable as edible vegetables.
They are rich in vitamin B, iron, calcium, zinc and protein and are more tolerant to diseases, pests and high moisture than many other leafy vegetables grown in the tropics (Pace et al., 1985; Woolfe, 1992; Yoshimoto et al., 2001; Islam, 2006; Yoshimoto et al.,2007). Sweetpotato is among the crops planned by the National Aeronautics and Space Administration of the United States to be grown in a controlled ecological life support system as a primary food source (Hoff et al., 1982).
Earlier experiments revealed that sweetpotato leaves are an excellent source of antioxidative compounds, namely, polyphenolics (Islam et al., 2002). We have recently isolated the 6-caffeic acid derivatives in sweetpotato leaves: Caffeic Acid (CA); 3-mono-o-caffeoylquinic acid (Chlorogenic acid; ChA); 3,4-di-o-caffeoylquinic acid (3,4-diCQA); 3,5-di-o-caffeoylquinic acid (3,5-diCQA); 4,5-di-o-caffeoylquinic acid (4,5-diCQA) and 3,4,5-tri-o-caffeoylquinic acid (3,4,5-triCQA) (Islam et al., 2002). ChA, di- and tri-CQA are esters of QA and one-, two- and three-caffeic acids, respectively. The structural feature responsible for the antioxidative and free Radical Scavenging Activity (RSA) of caffeic acid is the ortho-dihydroxyl functionality in the catechol ring (Son and Lewis, 2002). The recent development of screening methods for environmental carcinogens by determining their mutagenicity has enabled various types of mutagens and carcinogens to be detected and identified in daily foods (Ames et al., 1975; Yoshimoto et al.,2007). It is also now known that various types of inhibitors that act against mutagens and carcinogens exist in our daily food and that play important roles in reducing the risks of mutagenesis and carcinogenesis (Shinohara et al., 1988; Kurata et al., 2007).
Phenolic acids are bioactive compounds and a diverse group of secondary metabolites universally present in higher plants where they play important roles (Ho, 1992; Harborne, 1994). Phenolics also have attracted special attention because they may protect the human body from oxidative stress, which in turn is associated with many diseases including cancer and cardiovascular diseases, as well as aging (Peluso et al., 1995; Shimozono et al., 1996; Yoshimoto et al., 1999; Islam, 2006; Bovell-Benjami , 2007; Rowena et al., 2009; Nagy et al., 2009). Physiological functions of sweetpotato leaves and their derivatives have not been studied since all their derivatives have not been isolated from one specific plant. Recently, caffeoylquinic acid derivatives with mono-, di- and tri-caffeoyl residues were isolated from sweetpotato leaves the first time (Islam et al., 2002). However, little is known about the relationship between the physiological functions of caffeoylquinic acid derivatives of sweetpotato leaves. Therefore, working on this area is important for human health. The objective of this study was to characterize the foliar caffeoylquinic acid derivatives of sweet potatoes and investigate their chemo-preventive potentials, specifically RSA and antimutagenicity.
MATERIALS AND METHODS
Plant Materials and Cultural Methods
Sweetpotato tips of the 6 advanced lines which were selected from 60 genotypes according to total leaf polyphenol contents such as V-1, V-2, V-3, V-4, V-5 and V-6. The experiment was conducted during 2006 to 2008 at the Department of Agriculture of University of Arkansas, USA. Cuttings were planted in sterilized soil in 12 cm vinyl pots. Five pots were planted and replicated and arranged in a randomized block design. After 45 days, all the leaves (40-50 leaves) from each plant were harvested, washed gently, put into pre-labeled vinyl bags and then it was immediately frozen at -85°C. The following day the frozen samples were freeze-dried for 48 h and powdered using a blender prior to analysis.
Extraction and Measurement of Total Phenolics
Total phenolics were measured by the procedure described by Cosetang and Lee (1987) with a slight modification. The lyophilized powdered sweet potato leaf tissues (10 mg) were vigorously mixed with 1:10 (mg mL-1) 80% aqueous ethanol solution. The mixture were then boiled for 5 min under a hood, centrifuged at 5000 x g for 10 min and the supernatant was collected. The residue was mixed with 5 mL of 80% aqueous ethanol, boiled for 5 min to re-extract the phenolics and centrifuged under the same conditions. The extracts were combined, made up to 10 mL and used for the measurement of total phenolics. The alcohol extract was diluted to obtain an absorbance reading within the range of the standards (40-800 μg chlorogenic acid mL-1). The absorbance was then measured at 600 nm with a Microplate reader. The results were expressed as g/100 g Dry Mass (DM) (Padda and Picha, 2008).
Quantification of Phenolic Acids by High Pressure Liquid Chromatography (HPLC)
The lyophilized, powdered sweetpotato leaf tissue (50 mg) was vigorously mixed with 4 mL of 80% ethanol in a capped centrifuge tube (Islam et al., 2002; Rowena et al., 2009). The mixture was then boiled for 5 min in a hood and centrifuged at 3000 x g for 10 min. The supernatant was filtered through a cellulose acetate membrane filter (0.20 μm). A 5 μL portion of the filtrate was injected into the HPLC system and eluted as described below. The HPLC system consisted of two Model LC-10AT pumps, a Model SIL-10AXL auto injector, a Model CTO-10AC column oven and a Model SPD-M10AVP photodiode array UV-VIS detector. The column was a YMC-Pack ODS-AM-302 (150x4.6 mm i.d., 5 μm particles). The column oven temperature was set at 40°C. The mobile phase consisted of water containing 0.2% (v/v) formic acid (A) and methanol (B). Elution was performed with a linear gradient as follows: 2% B from 0 to 15 min, 2 to 45% B from 15 to 50 min and 45% B from 50 to 65 min. The flow rate was 1 mL min-1. The phenolics were detected at 326 nm. The retention times (Rt) of the phenolics compounds were compared with those of purified phenolics from sweetpotato leaves used as authentic standard (purity >97% estimated by the HPLC analysis (Islam et al., 2002). The chemical structures of the isolated caffeic acid derivatives are shown in Fig. 1.
Measurement of Radical Scavenging Activity Using the DPPH Method
The Radical Scavenging Activity (RSA) was measured according to Brand-Williams et al. (1995) with slight modifications (Padda and Picha, 2008; Nagy et al., 2009). 1, 1-Diphenyl-2-picrylhydrazyl (DPPH) was used as a stable radical. All reactions were run in a 96 well Microplate with a total volume of 300 μL. A 75 μL sample (80% ethanol extract) was combined with 150 μL, 0.1 M 2-(N-morpholino) ethanesulfonic acid (MES) (pH 6.0) and 75 μL DPPH in 50% ethanol in the microplate well and mixed.
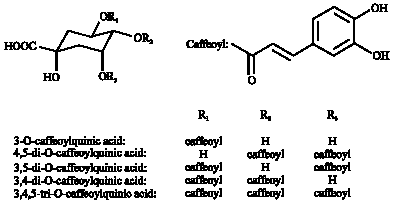 | |
| Fig. 1: | Chemical structures of caffeic acid derivatives in Ipomoea batatas L. leaves |
For the control, 75 μL of 80% ethanol was used in place of the sample. The reaction mixtures were shaken and held for 2 min at room temperature in the dark. Trolox (6-Hydroxy-2, 5, 7, 8-tetramethyl-chroman-2-carboxylic acid) was used as the reference antioxidant compound and 80% ethanol was used as the blank solution (without DPPH). The decrease in absorbance of DPPH at 520 nm was measured within 2 min. All samples were analyzed in triplicate. The RSA of samples (antioxidants) was expressed in terms of IC50 (concentration in μmole Trolox mg-1 DM required for a 50% decrease in absorbance of the DPPH radical). A plot of absorbance vs. concentration was made to calculate IC50. The results were expressed as μmole Trolox mg-1 DM. For the polyphenolic compositions (i.e., caffeic acid and derivatives) the DPPH-RSA was calculated according to the equation:
Assay of Antimutagenicity
The antimutagenicity assay was performed by a slight modification of the method of Yahagi et al. (1977) with slight modifications (Yoshimoto et al.,2007; Nagy et al., 2009). The antimutagenic activity was evaluated for Salmonella typhimurium TA 98 using a mutagen, Trp-P-1. These mutagens required metabolic activation to induce mutation in TA 98. S-9 mix contained 50 μmol of sodium phosphate buffer (pH 7.4), 4 μmol of MgCl2, 16.5 μmol of KCl, 2.5 μmol of glucose-6-phosphate, 2 μmole of NADH, 2 μmol of NADPH and 50 μL of S-9 fraction in a total volume of 0.5 mL. For the inhibition test, 0.1 mL of mutagen, 0.1 mL DMSO-dissolved polyphenolics solution and 0.5 mL of S-9 mix or phosphate buffer were simultaneously incubated with 0.1 mL of bacterial suspension at 37°C for 20 min and then poured on minimal-glucose-agar plates with 2 mL of soft agar. The colony number of each plate was counted after 48 h cultivation at 37°C.
Statistics
A randomized complete block design with five replications was used. Data for the different parameters were analyzed by analysis of variance (ANOVA) procedure and the level of significance was calculated from the F-value of ANOVA. The relation between total polyphenols and RSA was described with correlation analysis using the Excel program (Islam et al., 2002).
RESULTS
Total Polyphenol and Radical Scavenging Activity (RSA) of the Sweetpotato Genotypes
The total leaf polyphenol content of 6 advanced lines, which were selected based on their higher leaf polyphenol contents from 60 genotypes, were analyzed and characterized. Of all the genotypes studied, the total polyphenol ranges from 12.18 to 16.17 g/100 g DW (Fig. 2). The highest polyphenol content was found in 89SB-38 (V-5) (16, 17 g/100 g leaf powder) and the lowest in the B64-3 (V-3) (12.18 g/100 g leaf powder). RSA ranged from 1.09 to 1.85 μmol Trolox mg-1 (Fig. 3). The cultivar V-5 showed highest activity (1.85 μmole Trolox mg-1 DM) and V-3 was the lowest (1.07 μmol Trolox equivalence mg-1 DM). Among the 6 advanced lines studied, V-5 was designated as having highest polyphenol content. The correlation among RSA, total polyphenol and different caffeoylquininc acid derivatives is shown in Fig. 4.
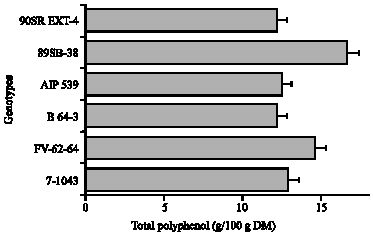 | |
| Fig. 2: | Total polyphenol contents (g/100 g DM) of the 6 genotypes of sweetpotato leaves. Bars indicate the standard error of the mean of five replications, when absent falls under the symbol. DM: Dry Matter |
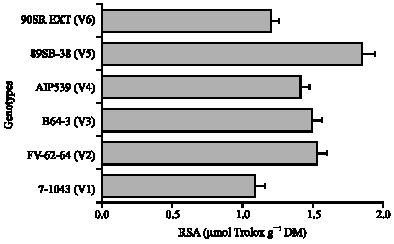 | |
| Fig. 3: | Radical Scavenging Activity (RSA; μmol Trolox g-1 DM) of the 6 genotypes of sweetpotato leaves. Bars indicate the standard error of the mean of five replications, when absent falls under the symbol. DM: Dry Matter |
The data demonstrate that there is a significant (p<0.001) positive linear correlation between total polyphenol contents and radical scavenging activity in sweetpotato leaves (r = 0.62; n = 90).
Antimutagenicity of Sweetpotato Genotypes
The antimutagenicity (% inhibition) of the six sweetpotato genotypes range from 77 to 96% (Fig. 5). The cultivar V-5 has highest activity (96%) followed by V2 (93%) and V-1 is the lowest (76%) followed by V6 (77%). Among the 6 advanced lines studied, V-5 is designated as having highest polyphenol content, RSA and antimutagenicity.
Polyphenolic Compositions of Sweetpotato Leaves
Six different caffeic acid derivatives were identified and quantified by HPLC analysis procedures (Fig. 6). All the RP-HPLC profiles of the genotypes tested showed peaks at the same retention times but peak areas of individual phenolic compounds differed. Six caffeoylquinic acid derivatives of sweetpotato leaves were identified and quantified.
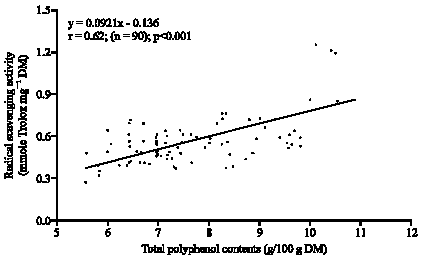 | |
| Fig. 4: | Linear correlations between the total polyphenol contents (g/100 g dry matter) and Radical Scavenging Activities (RSA; μmol Trolox g-1 DM) of sweetpotato leaves |
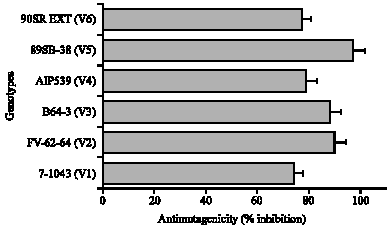 | |
| Fig. 5: | Antimutagenicity (% inhibition) of the 6 genotypes of sweetpotato leaves. Bars indicate the standard error of the mean of five replications, when absent falls under the symbol |
| Table 1: | Distribution of polyphenolic composition (caffeoylquinic acid derivatives; mg g-1 dry weight) of 6 genotypes of sweetpotatoes |
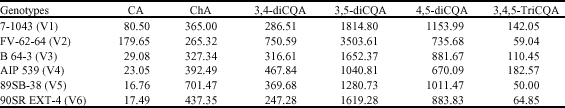 | |
| CA: Caffeic acid; ChA: Chlorogenic acid; 3,4-diCQA: 3,4-di-o-caffeoyl quinic acid; 3,5-diCQA: 3,5-di-o-caffeoyl quinic acid; 4,5-diCQA: 4,5-di-o-caffeoyl quinic acid; 3,4,5-triCQA: 3,4,5-tri-o-caffeoyl quinic acid | |
They include: caffeic acid; chlorogenic acid; 4, 5-di-O-caffeoylquinic acid; 3,5-di-O-caffeoylquinic acid; 3,4-di-O-caffeoylquinic acid and 3,4,5-tri-O-caffeoylquinic acid. Large variation was observed in relation to individual caffeic acid in sweetpotato leaves (Table 1).
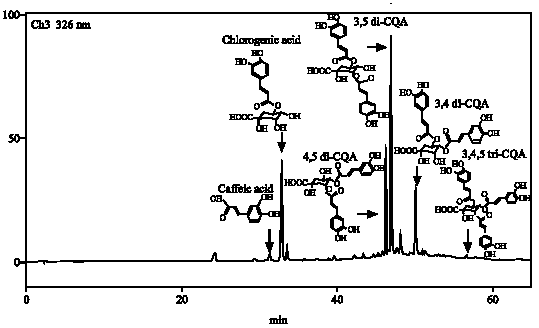 | |
| Fig. 6: | C18-HPLC separation chromatograms for caffeic acid derivatives in sweetpotato leaf extracts. Caffeic acid (Rt = 32.27 min); Chlorogenic acid (Rt = 33.42 min); 4,5-di-O-caffeoylquinic acid (Rt = 46.72 min); 3,5-di-O-caffeoylquinic acid (Rt = 47.34 min); 3,4-di-O-caffeoylquinic acid (Rt = 50.55 min); 3,4,5-tri-O-caffeoylquinic acid (Rt = 57.25 min); Rt: Retention time |
Radical Scavenging Activity (%) and Antimutagenicity (% Inhibition) of Caffeoylquinic Acid Derivatives of Sweetpotato Leaf Constituents
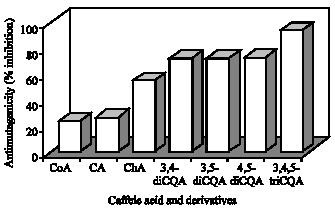
Coumaric Acid (CoA) has not been identified in the sweetpotato leaves but this component is one part of CQA derivatives. Therefore, CoA was used in this experiment to clarify whether binding of CoA to caffeoyl groups affects the RSA or not. The RSA (%) of CoA; CA; ChA; 3,4-dicQA; 3,5-diCQA; 4,5-diCQA and 3,4,5-triCQA were, 60, 61, 81, 43, 49, 48 and 71%, respectively (Fig. 7). The results indicate that differences between the di-CQA derivatives in relation to RSA were not significant. ChA showed the highest activity followed by 3,4,5-triCQA. There was no difference between CoA and CA and 3,4,5-triCQA was significantly higher activity (p<0.05) than that of the CoA and CA.
The antimutagenicity activity has been examined with three different concentrations: 1.0, 0.5 and 0.1 mM sample per plate. The effect was found to be a dose-dependent and the lowest dose of 0.1 mM showed the lower inhibition (Fig. 8). There were no differences between 1.0 and 0.5 mM concentrations (then, it is not dose dependent) and we used the values of 0.5 mM concentration. There was less difference between CoA and CA and the inhibition is negligible. ChA inhibited the reverse mutation by 41% in a dose range of 0.50 mM, while 3,4-diCQA; 3,5-diCQA and 4,5-diCQA, respectively inhibited the reverse mutation by 59, 59 and 61%. The 3,4,5-triCQA derivative inhibited the reverse mutation by 84% in the same dose of 0.50 mM. The di-CQAs exhibited almost similar antimutagenic activity in the same dose. Antimutagenicity of di-CQA derivatives and 3,4,5-triCQA was about 1.5 to 2 times higher than ChA, respectively. The result suggests that all the CQA derivatives effectively depressed the reverse mutation induced by Trp-P-1.
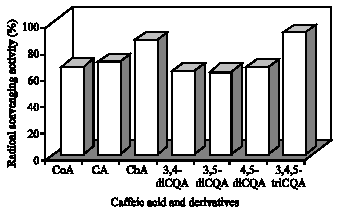 | |
| Fig. 7: | Radical scavenging activity (%) of caffeoylquinic acid derivatives isolated from sweetpotato leaves. Values are expressed as Mean±SE of three separate experiments done in triplicate. CoA: p-coumaric acid, CA: Caffeic acid; ChA: Chlorogenic acid; 3,4-diCQA: 3,4-di-O-Caffeoyl quinic acid; 3,5-diCQA: 3,5-di-O-Caffeoyl quinic acid; 4,5-diCQA: 4,5-di-O-Caffeoyl quinic acid; 3,4,5-triCQA: 3,4,5-tri-O-caffeoyl quinic acid |
 | |
| Fig. 8: | Effect of caffeoylquinic acid derivatives isolated from sweetpotato leaves on the antimutagenicity (% inhibition) of Trp-P-1 against Salmonella typhimurium TA 98. Trp-P-1 was added at a dose of 0.075 μg plate-1. The mutagenicity was tested with S-9 mix. Values are expressed as Mean±SE (standard error) of three separate experiments were done. The values shown have had the spontaneous mutation frequency subtracted. The His+ revertant values of the controls for the caffeoylquinic acid derivatives were 518±49 plate-1. Co: p-coumaric acid, CA: Caffeic acid; ChA: Chlorogenic acid; 3,4-diCQA: 3,4-di-O-caffeoyl quinic acid; 3,5-diCQA: 3,5-di-O-caffeoylquinic acid; 4,5-diCQA: 4,5-di-O-caffeoyl quinic acid; 3,4,5-triCQA: 3,4,5-tri-o-caffeoyl quinic acid |
DISCUSSION
Recent studies reveal that sweet potato leaves contain higher amounts of total polyphenols than certain other commercial vegetables, including sweet potato storage root and potato tubers (Walter et al., 1979; Yoshimoto et al., 2001, 2007). Therefore, the total leaf polyphenol content of 6 advanced lines, which were selected based on their higher leaf polyphenol contents from 60 genotypes, were analyzed and characterized. There were strong positive correlation among RSA, total polyphenol and different caffeoylquininc acid derivatives. The genotypes studied also showed high antimutagenicity. The phenolic acids are ubiquitous bioactive compounds found in plant foods and beverages. These phenolic compounds have received increased attention because of their potential antioxidant activities that may exert cardioprotective effects in humans (Islam, 2006; Yoshimoto et al.,2007; Nagy et al., 2009) and it has been shown that intake of these compounds is inversely related to coronary heart disease mortality (Yagasaki et al., 2000). The antioxidative and antimutagenic components in the storage root of sweet potato cultivars are primarily associated with phenolic compounds (Furuta et al., 1998; Yoshimoto et al., 1999, 2007). Since, polyphenol compounds show various physiological functions, sweetpotato leaves be expected to have physiologically active properties because they contain higher contents of polyphenolic compounds. These results maybe useful for various chemical breeding programs improving desirable organoleptic, nutritional quality characteristics of and potential chemo-preventative properties crop plants.
Polyphenolic Compositions of Sweetpotato Leaves
Six caffeoylquinic acid derivatives of sweetpotato leaves were identified and quantified. They include: caffeic acid, chlorogenic acid, 4,5-di-O-caffeoylquinic acid; 3,5-di-O-caffeoylquinic acid; 3,4-di-O-caffeoylquinic acid and 3,4,5-tri-O-caffeoylquinic acid. Large variation was observed in relation to individual caffeic acid in sweetpotato leaves. The result suggests that the main phenolic compound in sweetpotato leaves is 3,5-diCQA followed by 4,5-diCQA. The results of this study revealed that the sweetpotato leaf caffeoylquinic acid derivatives did vary according to genotype, but proportions were relatively similar in all the genotypes studied. It has been shown that intake of these compounds was inversely related to coronary heart disease mortality (Hertog et al., 1995; Knekt et al., 1996; Yoshimoto et al., 2007; Kurata et al., 2007). Since caffeic acid derivatives show various physiological functions, sweetpotato leaves might also be expected to have physiologically active properties because they contain higher contents of polyphenolics.
Wide variation was observed in relation to individual phenolic compound in sweetpotato leaves. The result suggests that the main phenolic compound in sweetpotato leaves is 3,5-di-O-caffeoylquinic acid followed by 4,5-di-O-caffeoylquinic acid. The average level of polyphenolic compounds in leaf of 60 genotypes in the following order: 3,5-di-O-caffeoylquinic acid >4,5-di-O-caffeoylquinic acid >3-O-caffeoylquinic acid >3,4-di-O-caffeoylquinic acid >3,4,5-tri-O-caffeoylquinic acid>caffeic acid. 3-O-caffeoylquinic acid of sweetpotato leaves was much higher than that of sweetpotato storage root, potato tubers and apple fruit (Islam, 2006; Yoshomoto et al., 2007) and it has various physiological functions (Kaul and Khanduja, 1998; Shimozono et al., 1996; Yoshomoto et al., 2007; Kurata et al., 2007; Padda and Picha, 2008). Among other polyphenolic compounds caffeic acid was the most effective inhibitor of tumor promotion in mice skin and the 3-O-caffeoylquinic acid, 3,4-di-O-caffeoylquinic acid, 3,5-di-O-caffeoylquinic acid and 4,5-di-O-caffeoylquinic acid, which were extracted from steamed sweetpotato root suppressed the melanogenesis equally (Kaul and Khanduja, 1998; Shimozono et al., 1996). Mahmood et al. (1993) reported that the 3,4,5-triCQA exhibited a greater selective inhibition of HIV replication than 4,5-di-O-caffeoylquinic acid and caffeic acid had only slight anti HIV activity.
Radical Scavenging Activity (%) and Antimutagenicity (% Inhibition) of Caffeoylquinic Acid Derivatives of Sweetpotato Leaf Constituents
The results of the study indicate that differences between the di-CQA derivatives in relation to RSA were not significant. ChA showed the highest activity followed by 3,4,5- triCQA.
| Table 2: | 3,4,5-tri-caffecoylquinic acid contents in sweetpotato leaves and other plant materials |
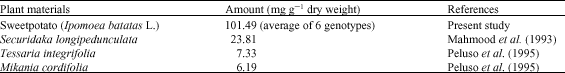 | |
There was no difference between CoA and CA and 3,4,5-triCQA was significantly higher activity (p<0.05) than that of the CoA and CA. This result is not consistent with the report of Murata et al. (1995) that esterfication with CoA lowered the activity of ChA on the antioxidative activity. This inconsistency in results may be due to genotype differences or age of leaves used for analysis. The effect of antimutagenicity activity was found to be a dose-dependent and the lowest dose of 0.1 mM showed the lower inhibition. The result suggests that all the CQA derivatives effectively depressed the reverse mutation induced by Trp-P-1. Yoshimoto et al. (2001) reported that the catechol structure plays an important role in the strong antimutagenicity. Furthermore, Yagasaki et al. (2000) suggested the possible involvement of the 3,4-dihydroxyl group of CA in the suppression of hepatoma invasion in vitro from an experiment using CA, cinnamic acid and p-coumaric acid (4-hydroxycinnamic acid). ChA, di- and tricaffeoylquinic acid are esters of quinic acid and one-, two- and three-caffeic acids and it has various physiological functions (Kapil et al., 1995; Tsuchiya et al., 1996; Yagasaki et al., 2000; Kurata et el., 2007; Yoshomoto et al., 2007; Padda and Picha, 2008). These results suggest that an increased caffeoyl group promotes antimutagenicity. Among other polyphenolic compounds, CA has been shown to be the most effective inhibitor of tumor promotion in the skin of mice and ChA, 3,4-diCQA; 3,5-diCQA and 4,5-diCQA, which were extracted from steamed sweetpotato, suppressed melanogenesis equally effectively (Shimozono et al., 1996; Kaul and Khanduja, 1998). The 3,5-diCQA and 4, 5-diCQA from Tessaria integrifolia and Mikania cordifolia exhibited an appreciable anti-inflammatory activity in vitro, while the tricaffeoyl derivative was inactive (Peluso et al., 1995). The compounds, CA, ChA and 3,5-diCQA exhibited over 50% inhibition of the histamine secretion induced by concanavalin plus phosphatidylserine from mast cells (Kimura et al., 1985). Furthermore, 3,4,5-triCQA exhibited a greater selective inhibition of HIV replication than 4,5-diCQA and CA had only slight anti HIV activity (Mahmood et al., 1993). The present data demonstrate that sweetpotato leaf accumulates higher concentration of 3,4,5-triCQA than other plant materials reported in the literature (Table 2) (Mahmood et al., 1993; Peluso et al., 1995). Furthermore, present study indicate that the RSA (except ChA) and antimutagenicity of 3,4,5-triCQA are more effective than mono- or di-caffeoylquinic acids. The importance of di- or tri-caffeoyl derivatives on physiological activities and the number of catechol structures in the caffeoyl derivatives play a significant role for these physiological functions (Kurata et el., 2007; Yoshomoto et al., 2001, 2007).
CONCLUSION
Results from this study shows that sweetpotato leaves contain a very high concentration of biologically active caffeoylquinic acid derivatives that exhibit enhanced chemo-preventative activities such as antimutagenic and antioxidative properties, which might have values in the prevention of certain human conditions like cancer and cardiovascular disease. These results may help in future programs of breeding for specific constituents and to enhance recent progress in molecular biology aimed at increasing antioxidant food supplement components available for consumption, which play an important dietary role in human health.
ACKNOWLEDGMENTS
This research was a part of the Science project done by Intisar Islam. These results presented in different local, regional and national science fairs in United State of America and received several awards of excellences. The author acknowledges his mentors for their generous help and cooperation. The first author is also thankful to his science teachers Mrs. Melissa Donham, Annice Steadman and Dr. Beth Maris.
REFERENCES
- Ames, B.N., J. McCann and E. Yamasaki, 1975. Methods for detecting carcinogens and mutagens with Salmonella/mammalian microsome mutagenicity test. Mut. Res., 31: 347-364.
PubMedDirect Link - Bovell-Benjamin, A.C., 2007. Sweet potato: A review of its past, present and future role in human nutrition. Adv. Food Nutr. Res., 52: 1-59.
CrossRef - Brand-Williams, W., M.E. Cuvelier and C. Berset, 1995. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol., 28: 25-30.
CrossRefDirect Link - Coseteng, M.Y. and C.Y. Lee, 1987. Changes in apple polyphenoloxidase and polyphenol concentrations in relation to degree of browning. J. Food Sci., 52: 985-989.
CrossRefDirect Link - Hertog, M.G., D. Kromhout, C. Aravanis, H. Blackburn, R. Buzina and F. Fidonza et al., 1995. Flavonoid intake and long-term disk of coronary heart disease and cancer in the seven countries study. Arch. Int. Med., 155: 381-386.
PubMed - Islam, S., M. Yoshimoto, S. Yahara, S. Okuno, K. Ishiguro and O. Yamakawa, 2002. Identification and characterization of foliar polyphenolic composition in sweetpotato (Ipomoea batatas L.) genotypes. J. Agric. Food Chem., 50: 3718-3722.
CrossRefDirect Link - Islam, S., 2006. Sweetpotato (Ipomoea batatas L.) Leaf: Its potential effect on human health and nutrition. J. Food Sci., 71: R13-R21.
CrossRefDirect Link - Kapil, A., O.P. Suri and I.B. Koul, 1995. Antihepatotoxic effects of chlorogenic acid from Anthrocephalus cadamba. Phytother. Res., 9: 189-193.
CrossRef - Kaul, A. and K.L. Khanduja, 1998. Polyphenols inhibit promotional phase of tumorigenesis: Relevance of superoxide radicals. Nutr. Cancer, 32: 81-85.
Direct Link - Knekt, P., R. Jarvinen, A. Reunanen and J. Maatela, 1996. Flavonoid intake and coronary mortality in Finland: A cohort study. Br. Med. J., 312: 478-481.
Direct Link - Kurata, R., A. Masaru, O. Yamakawa and M. Yoshimoto, 2007. Growth suppression of human cancer cells by polyphenolics from sweetpotato (Ipomoea batatas L.) leaves. J. Agric. Food Chem., 55: 185-190.
CrossRefDirect Link - Nwinyi, S.C., 1993. Effect of age at shoot removal on tuber and shoot yields at harvest of five sweetpotato (Ipomoea batatas (L.) Lam) cultivars. Field Crops Res., 29: 47-54.
CrossRef - Padda, M.S. and D.H. Picha, 2008. Quantification of phenolic acids and antioxidant activity in sweetpotato genotypes. Sci. Hort., 119: 17-20.
CrossRef - Rumbaoa, R.G.O., D.F. Cornago and I.M. Geronimo, 2009. Phenolic content and antioxidant capacity of Philippine sweet potato (Ipomoea batatas) varieties. Food Chem., 113: 1133-1138.
CrossRefDirect Link - Son, S. and B.A. Lewis, 2002. Free radical scavenging and antioxidative activity of caffeic acid amide and ester analogues: Structure-activity relationship. J. Agric. Food Chem., 50: 468-472.
CrossRefDirect Link - Walter, W.M., A.E. Purcell and G.K. McCollum, 1979. Use of high-pressure liquid chromatography for analysis of sweetpotato phenolics. J. Agric. Food Chem., 27: 938-941.
Direct Link - Yagasaki, K., Y. Miura, R. Okauchi and T. Furuse, 2000. Inhibitory effects of chlorogenic acids and its related compounds on the invasion of hepatoma cells in culture. Cytotechnology, 33: 229-235.
CrossRefPubMedDirect Link - Yoshimoto, M., S. Okuno, T. Kumagai, M. Yoshinaga and O. Yamakawa, 1999. Distribution of antimutagenic components in colored sweetpotatoes. Jap. Agric. Res. Q., 33: 143-148.
Direct Link - Yoshimoto, M., S. Okuno, M. Yamaguchi and O. Yamakawa, 2001. Antimutagenicity of deacylated anthocyanins in purple-fleshed sweetpotato. Biosci. Biotechnol. Biochem., 65: 1652-1655.
CrossRefPubMedDirect Link - Yoshimoto, M., R. Kurata, S. Okuno, K. Ishiguro and O. Yamakawa et al., 2007. Nutritional value and physiological functions of sweetpotato leaves. Acta Hort., 703: 107-116.
Direct Link - Furuta, S., I. Suda, Y. Nishiba and O. Yamakawa, 1998. High tert-butylperoxyl radical scavenging activities of sweet potato cultivars with purple flesh. Food Sci. Technol. Int. Tokyo, 4: 33-35.
CrossRefDirect Link