
M. A. El-tayeb
Department of Botany, Faculty of Science, South Valley University, Qena 83523, Egypt
A. E. El-Enany
Department of Botany, Faculty of Science, Assiut University, Assiut, Egypt
N. l. Ahmed
Department of Botany, Faculty of Science, South Valley University, Qena 83523, Egypt
International Journal of Botany
Year: 2006 | Volume: 2 | Issue: 4 | Page No.: 372-379







ABSTRACT
The ameliorative effect of salicylic acid (SA: 0.5 mM) under Cu stress (5 mg L–1) in sunflower (Helianthus annuus L.) was studied. Excess Cu reduced the fresh and dry weights of different organs (roots, stems and leaves) and photosynthetic pigments (chlorophyll a, b and carotenoids) in four-week-old plants. There was a considerable increase in Chl a/b ratio and lipid peroxidation in both the roots and leaves of plants under excess Cu. Soluble sugars and free amino acids in the roots also decreased under Cu stress. However, soluble sugars in the leaves, free amino acids in the stems and leaves and proline content in all the plant organs increased in response to Cu toxicity. Salicylic Acid (SA) significantly reduced the chlorophyll a/b ratio and the level of lipid peroxidation in Cu-stressed plants. Under excess Cu, a higher accumulation of soluble sugars, soluble proteins and free amino acids including proline occurred in plants treated with SA. Exogenous application of SA appeared to induce an adaptive response to Cu toxicity including the accumulation of organic solutes leading to protective reactions to the photosynthetic pigments and a reduction in membranes damage in sunflower.
PDF Abstract XML References Citation
How to cite this article
M. A. El-tayeb, A. E. El-Enany and N. l. Ahmed, 2006. Salicylic Acid-induced Adaptive Response to Copper Stress in Sunflower (Helianthus annuus L.). International Journal of Botany, 2: 372-379.
DOI: 10.3923/ijb.2006.372.379
URL: https://scialert.net/abstract/?doi=ijb.2006.372.379
DOI: 10.3923/ijb.2006.372.379
URL: https://scialert.net/abstract/?doi=ijb.2006.372.379
INTRODUCTION
Complementary biochemical and genetical approaches are being used to dissect the signaling network that regulates the innate immune response in plants. Receptor-mediated recognition of abiotic and biotic stresses triggers a signal amplification loop that is based on synergistic interactions between nitric oxide, reactive oxygen intermediates and Salicylic Acid (SA). Alternative resistance mechanisms deployed against different types of stresses are mediated by either SA or growth regulators. SA (2-hydroxybenzoic acid), one of the plant hormones (Raskin, 1992), is present in all plants organs and its concentration rises when the cell, organs or whole plants are subjected to biotic and biotic stresses.
Environmental pollution by heavy metals has received increasing attention over the last few decades. Heavy metals in soil may come from various anthropogenic sources, such as atmospheric deposition from industry, phosphate fertilizers and wastewater sludges (Williams and David, 1976). Copper (Cu) loading of agricultural soils may originate from the application of sewage sludge or fungicidal sprays. Although Cu is an essential element for plant growth (Arnon and Stout 1939), its accumulation in soils may be toxic to biota, such as plants (Baryla et al., 2000) or microorganisms (Dumestre et al., 1999) and at toxic concentrations it interferes with numerous physiological processes (Fernandes and Henriques 1991). It is also known to damage cell membranes by binding to sulphydryl groups of membrane proteins and inducing lipid peroxidation (De Vos et al., 1989, 1992). Cu is extremely toxic and can catalyze the formation of active oxygen species in the cell in Haber-Weiss reaction (Kurepa et al., 1997). Since oxidative stress is produced in plants exposed to high metal concentrations, the implication of some antioxidant enzymes may complement the role of phytochelatins in the cellular response to metal toxicity (Mazhoudi et al., 1997).
Exogenous application of SA results in the activation of a range of plant defense genes. The SA level in plants increases in response to infection, exposure to ultraviolet light and ozone and is believed to be the part of a signaling process that results in systemic acquired resistance (Rasmussen et al., 1991; Delaney et al., 1994). It may sensitize the cells by acting as a signal in the transduction pathway for rapid activation of defense genes.
The present study was conducted to asses whether exogenous application of SA through the rooting medium could ameliorate the adverse effects of Cu toxicity on sunflower plants. Growth characteristics, lipid peroxidation, carbohydrates, proteins, total free amino acids including proline in the roots and shoots of sunflower plants were examined.
MATERIALS AND METHODS
Plant material: Seeds of Helianthus annuus L. obtained from the Agronomy Department, Faculty of Agriculture, Assiut University, Assiut, Egypt were germinated on moist filter papers in the dark at room temperature. Four-day-old seedlings were placed in plastic pots containing 1l nutrient solution. The germinated seeds were selected and transferred to half-strength Hoagland’s solution (Arnon and Hoagland, 1940) and the solution pH was maintained at 5.5±0.2. Four-day-old seedlings were planted in each plastic pot (five seedlings per pot), containing 1 l of freshly prepared nutrient solution.
Cu and salicylic acid treatments: Two-weeks old plants were treated with two concentrations of Cu (0 and 5 mg L–1) supplied as CuCl2 in the presence and absence of 0.5 mM salicylic acid. This was carried out by exposing the plant roots for two weeks to nutrient solution containing 5 mg L–1 Cu with Cu-free half- strength nutrient solution used as a control. All the solutions were renewed at an interval of every three days and the pH was adjusted to 5.5±0.2. There were three replications per treatment with five plants per replication. The harvested plants were divided into roots, stems and leaves and then stored at-20°C for biochemical analysis.
After four weeks, fresh and dry weights of different organs (roots, stems and leaves) were determined. The photosynthetic pigments (chlorophyll a, b and carotenoids) were determined according to Metzner et al. (1965). The results are presented as the mean of six measurements over three replications for each treatment.
Lipid peroxidation: Lipid peroxidation was determined as 2-thiobarbituric acid (TBA) reactive substances, i.e., malondialdehyde (MDA). Briefly, 0.25 g tissue sample was homogenized in 5 mL of 0.1% trichloroacetic acid (TCA) followed by centrifugation at 10,000 x g for 5 min. To 1 mL aliquot of the supernatant, 4 mL of 20% TCA containing 0.5% TBA was added. The mixture was heated at 95°C for 15 min and cooled immediately. The developed colour was extracted with 2 mL n-butanol and the absorbance was measured at 532 nm followed by the substraction of the value of non-specific absorption at 600 nm. The level of lipid peroxidation was expressed as nmol of MDA g–1 fresh weight at an extinction coefficient of 155 mM cm–1 (Heath and Packer, 1968). The results are presented as the mean of three replicate estimations for each treatment.
Estimation of soluble, insoluble and total sugars: The water-soluble sugars and total sugars were quantified by the anthrone sulfuric acid method (Fales, 1951). For estimation of total sugars, a known weight of dried tissue material was heated in a water bath at 100°C for 2 h in 10 mL of 2 N HCl. The solution was cooled and transferred to a 100 mL measuring flask after filtration, followed by making up to a known volume by distilled water. To 0.5 mL of the prepared solution, 4.5 mL of anthrone reagent was added and the total carbohydrate content was calculated as mg g–1 dry weight of the plant organ. To estimate water-soluble sugars, a known weight of the dried tissue material was boiled in 10 mL of distilled water for 2 h; after cooling the hydrolysate was filtered and then made up to 50 mL. The water-soluble sugars were estimated by the method of anthrone sulfuric reagent as above and calculated as the differences between the amount of the total and water-soluble sugars of the same sample. The results are presented as the mean of three replicate estimations for each treatment.
Estimation of protein and free amino acids: To estimate soluble proteins, tissue samples (50 mg) were boiled in 10 mL distilled water for 2 h. After cooling, the water extract was centrifuged at 6000 x g, the supernatant was decanted and made up to a definite volume by distilled water and the soluble proteins were determined according to Lowry et al. (1951). For water-insoluble proteins, the water-insoluble residue remaining after the extraction of soluble proteins was homogenized with 10 mL of 1 N NaOH for at least 30 min. Then, 1 mL of this homogenate was added to 5 mL of alkaline reagent solution without NaOH, mixed thoroughly and allowed to stand at room temperature for at least 10 min followed by the addition of 0.5 mL Folin-Ciocalteu reagent. After 30 min, the extinction was measured against an appropriate blank at 700 nm. Total protein content was calculated by summing the soluble and insoluble protein fractions of the same sample. Free amino acids were extracted from plant tissues and determined according to the method of Moore and Stein (1948). However, in this method traces of proline and hydroxyproline are encountered. A caliberation curve was constructed using glycine. The concentrations of free amino acids were calculated as mg g–1 dry weight. Free proline was determined according to Bates et al. (1973). Brifely, 100 mg of tissue powder was homogenized in 10 mL of 3% aqueous sulfosalicylic acid for 10 min followed by filtration. Two milliter of the filtrate were mixed with 2 mL of galcial acetic acid and 2 mL of acid ninhydrine for 1 h at 100°C. The developed colour was extracted in 4 mL toluene and measured colourimetrically at 520 nm against toluene. A standard curve with proline was used for the final calculations. The results are the mean of three replicates for each treatment.
Statistical analyses: The data were statistically analyzed by one-way analysis of variance (PC-STAT, Ver 1A, Pilot Edition, Georgia University, Georgia) and the Least Significant Difference (LSD) test was used to separate the means at p≤0.05.
RESULTS AND DISCUSSION
In higher plants, heavy metal toxicity is generally associated with growth inhibition and reduction in biomass production (Woolhouse, 1983). Inhibition of both cell elongation and division by heavy metals results in a decline in biomass production (Arduini et al., 1995). The present study indicated that Cu stress reduced the growth of roots, stems and leaves (Fig. 1). These results are in agreement with those reported by others (Chen et al., 2001; Wang et al., 2004). Chen et al. (2001) reported that Cu-induced inhibition of root growth was associated with H2O2-dependent peroxidase-catalyzed formation of cross-linking cell wall polymers. The growth inhibition in the presence of heavy metals might be due to some disturbances, such as the cellular water status (Burzynski and Grabowski, 1984), mitosis (Roderer, 1979), cell cycle (Wierzbicka, 1999) and stiffening of cell walls (Qureshi et al., 1996). We showed that SA application increased the growth of roots, stems and leaves of both the control and Cu-stressed plants. A similar increase in shoot and root growth of maize plants under salt stress in response to SA treatment has been reported earlier (Khodary, 2004). Dhaliwal et al. (1997) also reported that SA increased the leaf area in sugarcane plants.
The main site of attack in plant cells by a redox active metal is usually the cell membrane. Heavy metals induce severe lipid peroxidation due to the removal of hydrogen by Reactive Oxygen Species (ROS) from unsaturated fatty acids leading to the formation of lipid radicals. Lipid molecules in general and unsaturated ones in particular are sensitive to oxidation by ROS generated under stress conditions. As a result, the presence of elevated levels of lipid peroxides is generally accepted as an indicator of severe oxidative stress. The induction of oxidative stress in plants by heavy metals stress is well documented (Dietz et al., 1999). Cu, as one of the redox-active metals which catalyzes the Fenton reaction, can accelerate the generation of highly damaging OH• radical from O2• and H2O2 (Avery, 2001). Baryla et al. (2000) demonstrated that lipid peroxidation of membranes can be used as a toxicity bioassay for plants, because it is very sensitive to Cu. In the present study, Cu stress increased the malondialdehyde (MDA) content of both the roots and leaves (Fig. 2). An increase in the level of lipid peroxidation in plants treated with excess Cu was reported in earlier studies (Mazhoudi et al., 1997; Chen et al., 2001). These results concluded that lipid peroxidation might result in a cascade of cyclical reactions leading to the repetitive formation of short chain alkanes and lipid acid aldehydes, thus totally destroying the lipid structure. This further leads to dimerization and polymerization of proteins, which are considered to be the most damaging to the membranes.
We observed that SA increased the MDA content in the absence of Cu in both the roots and leaves of sunflower plants (Fig. 2), suggesting that it can induce oxidative stress in plants even in the absence of heavy metal stress. Similar results were also obtained by Rao et al. (1997) and Metwally et al. (2003). SA has the ability to be converted to oxidized SA inducing more free radicals (FRs); these FRs are capable of lipid peroxidation and modification of other macromolecules (Rao et al., 1997). In the present study, SA was found to decrease the lipid peroxidation in the roots and leaves of Cu-stressed sunflower (Fig. 2). The results indicate that SA could decrease the lipid peroxidation in sunflower plants by reducing the generation of OH• under Cu stress. In consistent with this, Kawano and Muto (2000) reported that the generation of OH• by Fenton-taype reaction in tobacco cell suspension was lowered by SA. It is suggested that SA could chelate the transition metals, thus reducing the formation of OH• and/or could act directly as a OH• scavenger. The role of SA as a OH• scavenger in plants has been suggested (Fry, 1998).
Further evidence for the role played by heavy metals in modifying the plant metabolism can be obtained from pigments analysis as obtained in this study. These results clearly demonstrated that Cu stress affected a reduction in photosynthetic pigments (Table 1). Excess Cu has been shown to cause a reduction in chlorophyll contents and inhibit some photosynthetic functions in the leaves (Prasad et al., 2001). Since Cu is an essential micronutrient and active component of the structure of different photosynthetic proteins and enzymes, it is particularly damaging to the chloroplasts when imported in excess (Fernandes and Henriques, 1991). Present results showed that SA caused a general increase in chlorophyll concentrations in both the control and Cu-stressed plants (Table 1).
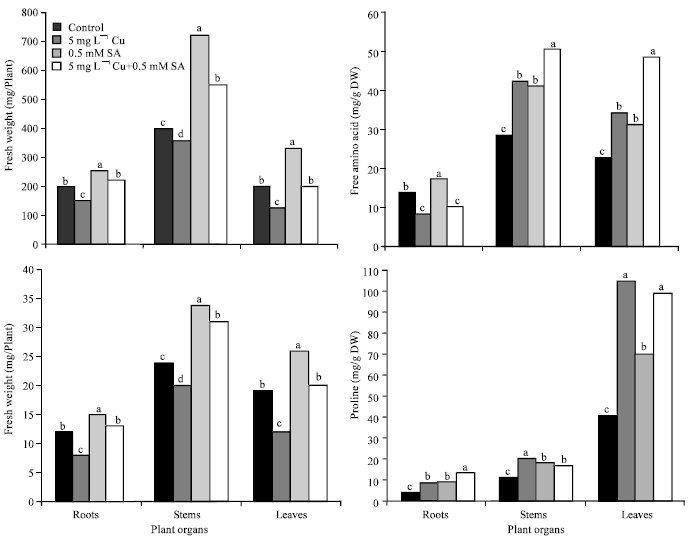 | |
| Fig. 1: | Fresh and dry weights, free amino acids and proline content in different organs of four-week-old sunflower plants under copper (Cu) stress and salicylic acid (SA) treatment. Means with the common letters are not significantly different at p≤0.05, according to Least Significant Difference (LSD) test |
| Table 1: | Effects of copper (Cu) stress and salicylic acid (SA) on photosynthetic pigmentsa in four-week-old sunflower plants |
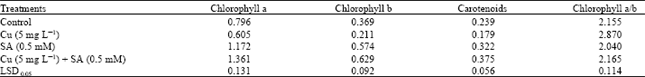 | |
| aThe values are expressed as mg g–1 dry weight | |
In agreement with this, SA was also reported to increase the chlorophyll content and stimulate the photosynthetic machinery in maize both under control and salinity stress (Khodary, 2004).
In higher plants, carotenoids protect the photosynthetic apparatus from excess photons and oxidative stress (Young, 1991). In the present study, SA significantly increased the total carotenoid contents in the leaves of sunflower plants under control as well as Cu-stress (Table 1). Rao et al. (1997) found that SA caused an increase in the carotenoids content in Arabidopsis thaliana leaves under normal conditions. In addition to carotenoid contents, the size of xanthophyll pool and de-epoxidation rate increased significantly with an increase in SA concentrations in both wheat and moong seedlings (Moharekar et al., 2003). In this study, exogenous SA application increased the endogenous SA levels (data not shown) in Cu-stressed plants. The data, presented in Table 1, also showed that Chl a/b ratio was higher in Cu-stressed plants than that in control ones. SA application lowered the Chl a/b ratio in both the control and Cu-stressed plants. It has also been reported that SA decreased the Chl a/b ratio in wheat plants (Moharekar et al., 2003). Present results indicate that SA-mediated counteraction of Cu toxicity in sunflower is associated with a reduction of chl a/b ratio as compared to control values.
| Table 2: | Effects of copper (Cu) stress and salicylic acid (SA) on soluble, insoluble and total sugarsa in various organs of four-week-old sunflower plants |
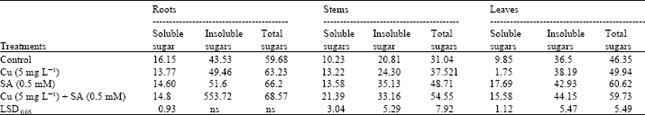 | |
| a The values are expressed as mg g–1 dry weight | |
| Table 3: | Effects of copper (Cu) stress and salicylic acid (SA) on soluble, insoluble and total proteinsa in various organs of four-week-old sunflower plants |
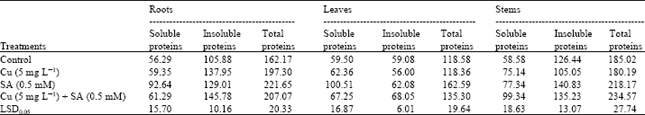 | |
| aThe values are expressed as mg g–1 dry weight | |
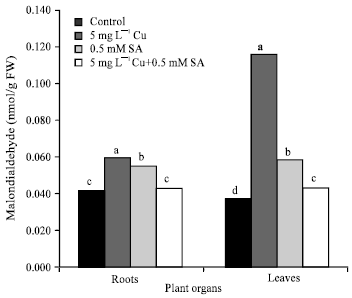 | |
| Fig. 2: | Lipid peroxidation (malondialdehyde content) in the roots and leaves of four-week-old sunflower plants under copper (Cu) stress and salicylic acid (SA) treatment. Means with the common letters are not significantly different at p = 0.05, according to Least Significant Difference (LSD) test |
Carbohydrates that represent one of the main organic constituents of the dry matter were found to be affected by excess of Cu. There was a significant increase in soluble sugars in the leaves when the plants were exposed to Cu stress (Table 1). Similar results were also obtained in cucumber in response to Cu stress (Alaoui-Sosse et al., 2004). On the other hand, a significant reduction in soluble sugars occurred in the roots in response to Cu stress. Cd has also been reported to cause an inhibition in the content of soluble sugars of sunflower plants (Gadallah, 1995). However, in the present study, insoluble and total sugars in various parts of sunflower plants under Cu stress were more or less unaffected. SA treatment induced an increase in soluble, insoluble and total sugars contents of the stems and leaves of both the control and Cu-stressed plants (Table 1). These results are in agreement with those obtained by Khodary (2004).
We showed that Cu stress caused an increase in insoluble and total protein contents concomitant with a pronounced decrease in the concentrations of free amino acids in sunflower roots (Table 2 and Fig. 1). However, the soluble, insoluble and total proteins in stem, soluble proteins in roots and total proteins in the leaves were more or less unaffected. The accumulation of proteins in plant organs due to heavy metals is well known (Demirevska-Kepova et al., 2004). SA treatment induced a considerable increase in the content of protein fractions in various organs of control and Cu-stressed plants, (Table 3).This may be due to the interactive effect of heavy metals and salicylic acid and such stimulation in protein accumulation under stress conditions is well known (McCue et al., 2000; Kang et al., 2003). The data also showed that free amino acids were accumulated in the stems and leaves of Cu-stressed plants (Fig. 1). In agreement with this, Hall (2002) reported that the accumulation of amino acids, such as histidine, is a potential ligand for heavy metals and, therefore, could play a role in their tolerance and detoxification. In the present study, SA induced an enhanced accumulation of amino acids in the stems and leaves of the Cu-stressed plants. Previously, SA was also reported to stimulate the accumulation of free amino acids in barley under NaCl stress (El-Tayeb, 2005).
In the present study, excess Cu was found to be associated with an increase in proline contents of the roots, stems and leaves of sunflower plants (Fig. 1). This is in agreement with the results obtained in other species (Gadallah and El-Enany, 1999; Panda et al., 2003). SA induced an accumulation of proline in all the three organs of control plants. Under Cu stress, when SA was applied there was a reduction of proline accumulation in both the stems and leaves, but an induction occurred in the roots. Our results are in agreement with those reported by Shakirova et al. (2003). Thus, proline can be considered to be one of the important factors involved in SA-induced protective reactions in sunflower plants in response to Cu toxicity. Many investigators have reported the accumulation of proline in response to Cu toxicity (Chen et al., 2004; Lombardi and Sebastiani, 2005). In addition, exogenously applied proline in the growth medium has also been reported to alleviate the toxic effect of Cu (Backor et al., 2004). The mechanisms of proline action are not fully understood, but it has been suggested that its increased accumulation permits osmotic adjustment as well as provides protection for some of the enzymes (Sharma et al., 1998). It is assumed that this protection is due to a reduction in the activity of free metal ions and the formation of metal-proline complexes. Besides, proline is also capable of detoxifying the free radicals by forming a stable complex, thereby maintaining the NADP+/ NADPH ratios under stress at values similar to normal conditions (Hare and Cress, 1997).
In general, high levels of photosynthetic pigments in SA-treated plants and the associated increase in insoluble and total sugars may be responsible for the improved dry matter accumulation in sunflower under Cu stress. Overall, the present study indicates that exogenous application of SA could counteract the adverse effects of Cu stress by increasing the concentrations of organic solutes (soluble sugars, free amino acids and proline) for osmoregulation and/or stabilization of essential proteins. This counteracting effect of SA is reflected in higher fresh and dry weights and lower lipid peroxidation of membranes in sunflower plants. In conclusion, the results suggest that SA application may be useful for improving the plant growth and yield in Cu-contaminated areas.
REFERENCES
- Alaoui-Sosse, B., P. Genet, F. Vinit-Dunand, M.L. Toussain, D. Epron and P.M. Badot, 2004. Effect of copper on growth in cucumber plants (Cucumis sativus) and its relationships with carbohydrate accumulation and changes in ion contents. Plant Sci., 166: 1213-1218.
CrossRefDirect Link - Arduini, I., D.L. Godbold and A. Onnis, 1995. Influence of copper on root growth and morphology of Pinus pinea L. and Pinus pinaster Ait. Seedlings. Tree Physiol., 15: 411-415.
CrossRefDirect Link - Arnon, D.I. and P.R. Stout, 1939. The essentiality of certain elements in minute quantity for plants with special reference to copper. Plant Physiol., 14: 371-375.
CrossRefDirect Link - Avery, S.V., 2001. Metal toxicity in yeasts and the role of oxidative stress. Adv. Applied Microbiol., 49: 111-142.
PubMedDirect Link - Backor, M., D. Fahselt and C.T. Wu, 2004. Free proline content is positively correlated with copper tolerance of the lichen photobiont Trebouxia erici (Chlorophyta). Plant Sci., 167: 151-157.
CrossRefDirect Link - Baryla, A., C. Laborde, J.L. Montillet, C. Trianaphylides and P. Chagvardieff, 2000. Evaluation of lipid peroxidation as a toxicity bioassay for plants exposed to copper. Environ. Pollut., 109: 131-135.
CrossRefDirect Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Chen, L.M., C.C. Lin and C.H. Kao, 2000. Copper toxicity in rice seedlings: Changes in antioxidative enzyme activities, H2O2 level and cell wall peroxidase activity in roots. Bot. Bull. Acad. Sin., 41: 99-103.
Direct Link - Chen, C.T., T.H. Chen, K.F. Lo and C.Y. Chiu, 2004. Effects of proline on copper transport in rice seedlings under excess copper stress. Plant Sci., 166: 103-111.
Direct Link - De Vos, C.H.R., H. Schat, R. Vooijs and W.A.O. Ernst, 1989. Copper induced damage to the permeability barrier in roots of Silene cucubalus. J. Plant Physiol., 135: 164-169.
Direct Link - De Vos, C.H.R., M.J. Vonk, R. Vooijs and H. Schat, 1992. Glutathione depletion due to copper induced phytocheation synthesis causes oxidative stress in Silene cucubalus. Plant Physoil., 98: 853-858.
Direct Link - Delaney, T.P., S. Ukness, B. Vernooij, L. Friedrich and K. Weymann et al., 1994. A central role for salicylic acid in plant disease resistance. Science, 266: 1247-1250.
Direct Link - Demirevska-Kepova, K., L. Simova-Stoilova, Z. Stoyanova, R. Holzer and U. Feller, 2004. Biochemical changes in barley plants after excessive supply of copper and manganese. Environ. Exp. Bot., 52: 253-266.
CrossRefDirect Link - Dietz, K.J., U. Kramer and M. Baier, 1999. Free Radicals and Reactive Oxygen Species as Mediators of Heavy Metal Toxicity. In: Heavy Metal Stress in Plants: From Molecules to Ecosystems, Prasad, M.N.V. and J. Hagemeyer (Eds.). Springer-Verlag, Berlin, pp: 73-97.
Direct Link - Dumestre, A., S. Sauve, M. McBride, P. Baveye and J. Berthelin, 1999. Copper speciation and microbial activity in long-term contaminated soils. Arch. Environ. Contam. Toxicol., 36: 124-131.
Direct Link - El-Tayeb, M.A., 2005. Response of barley grains to the interactive e.ect of salinity and salicylic acid. Plant Growth Regul., 45: 215-224.
CrossRefDirect Link - Fales, F.W., 1951. The assimilation and degradation of carbohydrates by yeast cells. J. Biol. Chem., 193: 113-124.
PubMedDirect Link - Fernandes, J.C. and F.S. Henriques, 1991. Biochemical, physiological and structural effects of excess copper in plants. Bot. Rev., 57: 246-273.
CrossRefDirect Link - Gadallah, M.A.A., 1995. Effects of cadmium and kinetin on chlorophyll content, saccharides and dry matter accumulation in sunflower plants. Biol. Planta., 37: 233-240.
CrossRefDirect Link - Gadallah, M.M.A. and A.E. El-Enany, 1999. Role of kinetin in alleviation of copper and zinc toxicity in Lupinus termis plants. Plant Growth Regul., 29: 151-160.
Direct Link - Hall, J.L., 2002. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot., 53: 1-11.
CrossRefDirect Link - Hare, P.D. and W.A. Cress, 1997. Metabolic implications of stress-induced proline accumulation in plants. Plant Growth Regul., 21: 79-102.
CrossRefDirect Link - Heath, R.L. and L. Packer, 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125: 189-198.
CrossRefPubMedDirect Link - Kang, G., C. Wang, G. Sun and Z. Wang, 2003. Salicylic acid changes activities of H2O2-metabolizing enzymes and increase the chilling tolerance of banana seedlings. Environ. Exp. Bot., 50: 9-15.
Direct Link - Kawano, T. and S. Muto, 2000. Mechanism of peroxidase actions for salicylic acid‐induced generation of active oxygen species and an increase in cytosolic calcium in tobacco cell suspension culture. J. Exp. Bot., 51: 685-693.
CrossRefDirect Link - Khodary, S.E.A., 2004. Effect of salicylic acid on the growth, photosynthesis and carbohydrate metabolism in salt stressed maize plants. Int. J. Agric. Biol., 6: 5-8.
Direct Link - Kurepa, J., M. Van Montagu and D. Inze, 1997. Expression of sodCp and sodB genes in Nicotiana tabacum: Effect of light and copper excess. J. Exp. Bot., 48: 2007-2014.
Direct Link - Lombardi, L. and L. Sebastiani, 2005. Copper toxicity in Prunus cerasifera: Growth and antioxidant enzymes responses of in vitro grown plants. Plant Sci., 168: 797-802.
CrossRefDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mazhoudi, S., A. Chaoui, M.H. Ghorbal and E. El Ferjani, 1997. Response of antioxidative enzymes to excess copper in tomato (Lycopersicum esculentum, Mill). Plant Sci., 127: 129-137.
Direct Link - McCue, P., Z. Zheng, J.L. Pinkham and K. Shetty, 2000. A model for enhanced pea seedling vigour following low pH and salicylic acid treatments. Process Biochem., 35: 603-613.
CrossRef - Metwally, A., I. Finkemeier, M. Georgi and K.J. Dietz, 2003. Salicylic acid alleviates the cadmium toxicity in barley seedlings. Plant Physiol., 132: 272-281.
CrossRefDirect Link - Metzner, H., H. Rau and H. Senger, 1965. Untersuchunge zur synchronisierbarkarkeit einzelner-pigmentmangle-mutanten von chlorella. Planta, 65: 186-194.
Direct Link - Panda, S.K., I. Chaudhury and M.H. Khan, 2003. Heavy metals induce lipid peroxidation and affect antioxidants in wheat leaves. Biol. Plant., 46: 289-294.
CrossRefDirect Link - Prasad, M.N.V., L.P. Malec, Waloszek, M. Bojko and K. Strzalka, 2001. Physiological responses of Lemna trisulca L. (duckweed) to cadmium and copper bioaccumulation. Plant Sci., 161: 881-889.
Direct Link - Qureshi, J.A., K. Hardwick and H.A. Collin, 1986. Intracellular localization of lead in a lead tolerant and sensitive clone Anthoxanthum odoratum. J. Plant Physiol., 122: 357-367.
Direct Link - Rao, M.V., G. Paliyath, P. Ormrod, D.P. Murr and C.B. Watkins, 1997. Influence of salicylic acid on H2O2 production, oxidative stress and H2O2-metabolizing enzymes (Salicylic acid-mediated oxidative damage requires H2O2). Plant Physiol., 115: 137-149.
CrossRefDirect Link - Raskin, I., 1992. Salicylate, a new plant hormone. J. Plant Physiol., 99: 799-803.
CrossRefDirect Link - Rasmussen, J.B., R. Hammerschmidt and M.N. Zook, 1991. Systemic induction of salicylic acid accumulation in cucumber after inoculation with Pseudomonas syringae pv syringae. Plant Physiol., 97: 1342-1347.
CrossRefDirect Link - Shakirova, F.M., A.R. Sakhabutdinova, M.V. Bezrukova, R.A. Fathudinova and D.R. Fathutdinova, 2003. Changes in the hormonal status of wheat seedlings induced by salicylic acid and salinity. Plant Sci., 164: 317-322.
CrossRefDirect Link - Sharma, S.S., H. Schat and R. Vooijs, 1998. In vitro alleviation of heavy metal-induced enzyme inhibition by proline. Phytochemistry, 49: 1531-1535.
Direct Link - Wang, S.H., Z.M. Yang, H. Yang, B. Lu, S.Q. Li and Y.P. Lu, 2004. Copper-induced stress and anti-oxidative responses in roots of Brassica juncea L. Bot. Bull. Acad Sin., 45: 203-212.
Direct Link - Wierzbicka, M., 1999. The effect of lead on the cell cycle in the root meristem of Allium cepa L. Protoplasma, 207: 186-194.
CrossRefDirect Link - Young, A.J., 1991. The photoprotective role of carotenoids in higher plants. Physiol. Plant, 83: 702-708.
Direct Link - Fry, S.C., 1998. Oxidative scission of plant cell wall polysaccharides by ascorbate-induced hydroxyl radicals. Biochem. J., 32: 507-515.
Direct Link