
N. Bagheri
Department of Plant Breeding, College of Agriculture, University of Mazandaran, P.O. Box 578, Sari, Iran
N.B. Jelodar
Department of Plant Breeding, College of Agriculture, University of Mazandaran, P.O. Box 578, Sari, Iran
A. Ghanbari
Department of Plant Breeding, College of Agriculture, University of Mazandaran, P.O. Box 578, Sari, Iran
International Journal of Agricultural Research
Year: 2008 | Volume: 3 | Issue: 6 | Page No.: 386-396







ABSTRACT
In a diallel cross rice involving five varieties (Dailamani, Sange-tarom, Hasani, Shastak-Mohammadi and Daei-shastak), combining ability and gene action for yield and yield-related traits were estimated. The study indicated that both general and specific combining ability effects were significant and important for the more traits, except for primary branching of panicles and paddy width traits. This indicates the role of additive and non-additive gene effects in inheritance of the traits. Variances due to GCA were greater than those due to SCA for plant height, flag leaf width, primary and secondary branching of panicles and filled grain per panicle traits, suggesting the role of additive gene effects were predominant in inheritance of the traits. These characters have high heritability and selection for them may well succeed. But, low heritability of tiller number, flag leaf length, days to maturity, panicle length, paddy length and width, 1000-grain weight and grain yield characters, were because of a high amount of non-additive variance in the genetic variance. Thus these characters were important for production of hybrid variety and exploitation of heterosis. The GCA effects of each parent for these traits showed that the Shastak-Mohammadi and Daei-shastak are good general combiner for days to maturity and grain yield, respectively.
PDF Abstract XML References Citation
How to cite this article
N. Bagheri, N.B. Jelodar and A. Ghanbari, 2008. Diallel Analysis Study of Yield and Yield-Related Traits in Rice Genotypes. International Journal of Agricultural Research, 3: 386-396.
DOI: 10.3923/ijar.2008.386.396
URL: https://scialert.net/abstract/?doi=ijar.2008.386.396
DOI: 10.3923/ijar.2008.386.396
URL: https://scialert.net/abstract/?doi=ijar.2008.386.396
INTRODUCTION
New rice (Oryza sativa L.) varieties with early maturity and high yield potential are an important breeding challenge for Iran. Quantitatively genetic parameters such as heritability and variance components are useful for designing new breeding programmes. Predicting response to selection, allocating resource in field performance trials and constructing selection indexes.
Information and exact study of combining ability can be useful in regard to selection of breeding methods and selection of lines for hybrid combination (Can et al., 1997). Among different methods to assess the nature of gene action in the parents, the diallel cross technique is a systematic method (Verma, 2003). Many studies show that rice yield related characters (tiller No., filled grains per panicle and grain weight) and agronomic characters (plant height and days to flowering) are inherited quantitatively, related genetically to one another and influenced by growing environments (Kobayashi et al., 2003). The value amount of heterosis as well as the GCA and SCA effects is important considerations for hybrid breeding (Herrmann, 2007). Exploiting from diallel analysis in rice breeding has generally conducted to evaluate combining ability for quantitative traits and useful results are available. Despite the advances of molecular breeding classical quantitative genetics remains useful in practical rice improvement. Thus estimation of genetic parameters such as heritability and combining ability give inferences about the predominant action of the genes, indicates the appropriate selection strategy to be applied in the breeding program and allows the identification of the best parents (Torres and Geraldi, 2007).
This study used partial diallel analysis to determine the combining ability of Iranian rice cultivars and to study the genetic basis of important agronomic and grain yield related traits. Increasing yield in surface unit by using breeding and introducing of high yield varieties is one of the main goals of breeders. Most parts of rice cultivation farms in North districts of Iran devoted to cultivate of qualitative varieties and mid or late maturity varieties, which they have a low grain yield. Therefore breeding in order to early maturity and more yield are an important goal in breeding programs. Because successful conducting of each breeding program, it is necessary to aware of genetic structure, for this purpose.
MATERIALS AND METHODS
Plant Materials
Five rice genotypes (Hasani (long paddy width and high 1000-grain weight), Dailamani (long fag leaf and long paddy length), Shastak-Mohammadi (long flag leaf width and short days to maturity), Sange-tarom (low tiller No. and tall plant height) and Daei-shastak (high tiller No., high primary branching of panicles and high grain yield) and their 10 f1s hybrids obtained through half diallel set were evaluated for combining ability and gene effects. Single seedlings of each entry were transplanted at 20x20 cm spacing in 3x5 m2 plots in a randomized complete block design with three replications. This study was conducted in 2006-2007 years at Research Station of College of Agriculture, University of Mazandaran. General Combining Ability (GCA) and Specific Combining Ability (SCA) were calculated for all agronomic traits. In this study 13 traits includes tiller No., plant height, length and width of flag leaf, days to maturity, panicle length, primary and secondary branching of panicles, filled grain per panicle, length and width of paddy, 1000-grain weight and grain yield were evaluated based on standard evaluation system rice (Scshu, 1988).
Statistical Analysis
Data were recorded on ten randomly selected plant from parents and F1s plant samples. The Hayman, s (1954a, b) diallel analysis was used to compute the variance (Vr), covariance (Wr) and to construct the Wr, Vr graph. The analysis of variance for general and specific combining abilities was carried out according to Griffing (1956a, b) using method 2 (parents and F1s), Model 1 (fixed effects for genotypes). Statistical analysis of data was conducted by using of GENETIC ANALYSIS software (Deck, 1988).
RESULTS
Means of the Estimated Traits
In Table 1 means of traits classified using Duncan`s multiple range test (p = 0.05). Daei-shastak, Sange-tarom, Dailamani and Shastak-Mohammadi produced the highest respectively tiller No., plant height, flag leaf length and flag leaf width. The longest days to maturity were given by Daei-shastak but it was not significantly different from Sange-tarom, also Shastak-Mohammadi was given the least days to maturity. Tiller No. was least in Hasani but it was not significantly different from that of Sange-tarom. The least flag leaf width were given by Sange-tarom, Dailamani and Hasani in that order, also Shastak-Mohammadi was given the longest flag leaf width. The longest panicle length was given by Shastak-Mohammadi but it was not significantly different from Hasani and Dailamani. Primary branching of panicles was highest in Daei-shastak but it was not significantly in Hasani, Dailamani, Shastak-Mohammadi and Sange-tarom cultivars. Shastak-Mohammadi and Hasani produced the highest and the least secondary branching of panicles, respectively. Hasani produced the least filled grains per panicle but it was the highest in Daei-shastak. Paddy length was least in Hasani but it was not significantly different from that of Daei-shastak and Shastak-Mohammadi. Also Sange-tarom and Dailamani produced the least paddy width, but it was the longest in Shastak-Mohammadi. Hasani produced the highest 1000-grain weight, but it was the least in Sange-tarom and Dailamani cultivars. Grain yield was least in hasani but it was not significantly different from of Shastak-Mohammadi and Dailamani, but it was the highest in Daei-shastak (Table 1). These results showed that different genetic systems involved in controlling traits, which emphasized on important of study of these traits.
| Table 1: | Means of the estimated traits in 5 rice genotypes |
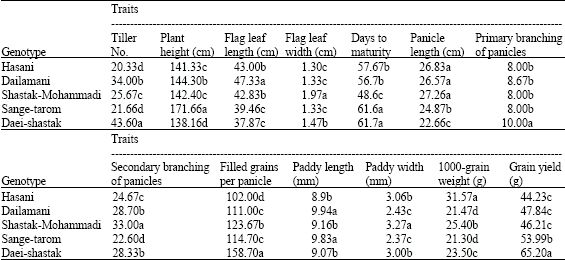 | |
| In each column, any two means having a common letter are not significantly at p = 0.05 based on Duncan`s multiple range test | |
Combining Ability Analysis
There were significant differences among the genotypes for characters, that lead to the combining ability analysis (Table 2). Thus were partitioned genetipic effects between genotypes into General Combining Ability (GCA) and Specific Combining Ability (SCA). The results showed significant differences between GCA and SCA for the investigated characters except for primary branching of panicles and paddy width traits. This indicates the role of additive and non-additive gene effects in inheritance of the traits. For total of traits, the values of MS (GCA) are more than MS (SCA) which indicated that there is more difference between parents for these traits and the role of additive variance inheritance of these traits is more than dominance variance (Verma, 2003; Torres and Geraldi, 2007). Regarding to the significance of gi in two directions in traits, we can declare that parents have potential of transfer of high and low values for each trait. Hence, in cases which increasing and decreasing the value of traits are desired, we should consider positive and negative values of gi, respectively.
In Table 3, it is observed that the most value of gi for trait of tiller No. is observed in Daei-shastak and Dailamani varieties therefore can use these for increasing tiller No. In case of plant height, it is observed that the most and the least value of gi is in Sange-tarom and Daei-shastak varieties, respectively. Regarding to dwarf in rice, it is accompanied by yield increasing, thus can use Daei-shastak variety in breeding programs for decreasing plant height. The effects of gi for traits of flag leaf length and width in Shastk-mahammadi variety is the most value and in Daei-shastak variety are the least value. Thus regarding to breeding programs, we can use these varieties for increasing or decreasing the desired traits in varieties production. Low and negative values of gi for days to maturity showed that earlier maturity desired in breeding programs in rice cultivars, thus can use Shastak-Mohammadi for decreasing days to maturity in hybrid production programs. The effects of gi for panicle length which is one of the yield components, in Shastak-Mohammadi variety, it has the highest positive and significant value. Hence for increasing this important component of yield, can use this variety in breeding programs. Also for traits of primary and secondary branching of panicles (of important components of the yield), Daei-Shastak and Shastak-Mohammadi have the most significant and positive value of gi, respectively. Therefore can use these varieties in breeding programs for increasing these traits. For trait of filled grains per panicle which is one of important components of the yield, Daei-shastak variety can be regarded as the best combiner in terms of increasing this trait in positive direction. The effect of gi for the trait of paddy length in Dailamani variety is the most value and in Hasani variety is the least value. Also the effects of gi for paddy width in Hasani and Shastak-mohammadi varieties are the most values and in Sange-tarom variety is the least value. Since in Iran country, most people prefer tall and thin seed rice or slender rice, therefore, for this purpose, can use Dailamani and Sange-tarom varieties in breeding programs. For the trait of 1000-grain weight (one of the other important components of the yield), Hasani variety can be considered as the best combiner in terms of increasing this trait in positive direction. For the trait of grain yield, Daei-shastak variety has the most value of gi.
| Table 2: | Analysis of variance for combining ability effects |
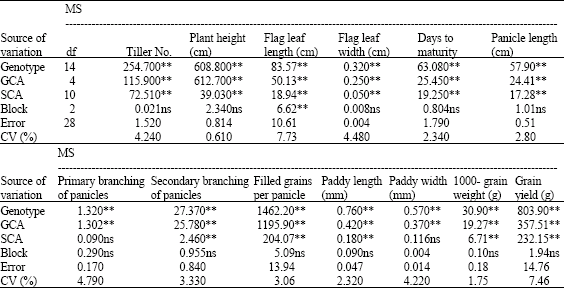 | |
| *, **Significant at p = 0.05 and 0.01 levels, based on an f-test, respectively, ns: Non-Significant | |
| Table 3: | General combining ability (Sgi) effects for traits in rice parents |
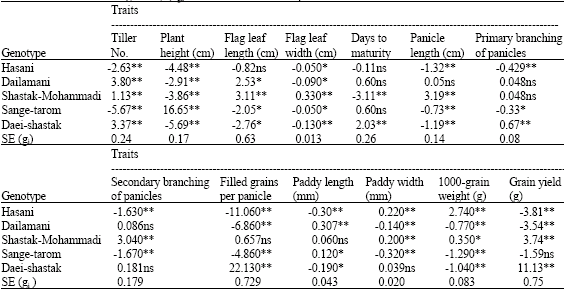 | |
| *, **General combining ability estimate significantly different from zero at p = 0.05 and 0.01, respectively, based on Duncan`s multiple range test | |
| Table 4: | Specific combining ability (sij) effects for traits in rice crosses |
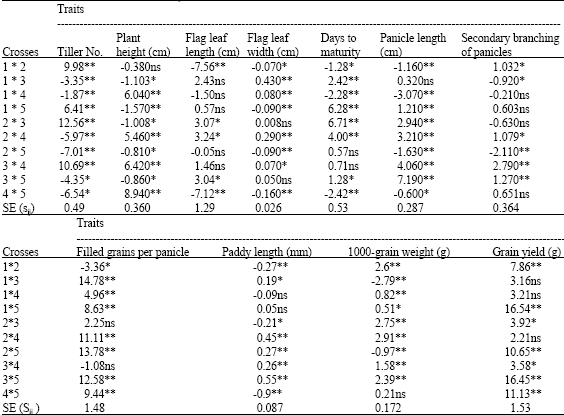 | |
| *, **Specific combining ability estimate significantly different from zero at p = 0.05 and 0.01, respectively, based on Duncan`s multiple range test | |
The value of specific combining ability of hybrids (Sij) for each of the studied traits, except of traits of primary branching of panicles and paddy width due to non-significant of its MS (SCA) has been presented in Table 4. For each of the studied traits, significant positive and negative SCA is observed. Positive and negative Sij represent high and low values SCA for that trait, respectively.
Sange-taromxDaei-shastak hybrid has the least value of Sij (-2.42**) for days to maturity, in result this hybrid determined as the best hybrid in terms of early maturity. Shastak-MohammadixDaei-shastak hybrid has the most value of Sij for grain yield, regarding to this that SCA of traits such as panicle length, filled grains per panicle and 1000-grain weight in Shastak-MohammadixDaei-shastak hybrid has almost higher values, this hybrid was recognized as the best hybrid in terms of grain yield. Also, this hybrid has the most paddy length (Table 4).
Regarding to the correctness of pre-hypothesis of Griffing model (1956b), on the basis of being diploid of parents, additive and dominance variances were calculated by formula of ![]() and by them and regarding to
and by them and regarding to ![]() formula, narrow sense heritability were estimated and along with MS (GCA)/MS (SCA), the importance of genetic variance components, the best general combiner and the best hybrid combination for each of the studied traits has given in Table 5. Regarding to this table, significant being of proportion of MS (GCA)/MS (SCA), being more of the proportion of additive effects of genes are determined for traits such as plant height, flag leaf width, primary and secondary branching of panicles and filled grains per panicle. Which due to has been caused being-high of the value of specific heritability of these traits. Therefore regarding to these obtained results, selection for these traits can be successful and/or namely, response to selection for these traits can be seen (Gravois and McNew, 1993). In Table 5, the best combiner for each of the traits and also the best hybrid have been introduced which by using of these information and other results of this research, rice breeders will be able to perform designing of breeding programs.
formula, narrow sense heritability were estimated and along with MS (GCA)/MS (SCA), the importance of genetic variance components, the best general combiner and the best hybrid combination for each of the studied traits has given in Table 5. Regarding to this table, significant being of proportion of MS (GCA)/MS (SCA), being more of the proportion of additive effects of genes are determined for traits such as plant height, flag leaf width, primary and secondary branching of panicles and filled grains per panicle. Which due to has been caused being-high of the value of specific heritability of these traits. Therefore regarding to these obtained results, selection for these traits can be successful and/or namely, response to selection for these traits can be seen (Gravois and McNew, 1993). In Table 5, the best combiner for each of the traits and also the best hybrid have been introduced which by using of these information and other results of this research, rice breeders will be able to perform designing of breeding programs.
| Table 5: | Proportion of squares means of GCA to SCA, gene action, additive variance, dominance variance, general and specific heritability, the best general combiner and the best combination for the studied traits |
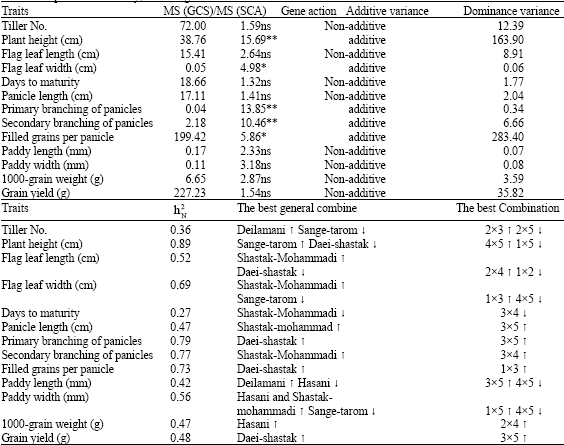 | |
| * and ** Significant in probability levels of 0.05 and 0.01, respectively, ns: Non Significant | |
Parameters of D, F, H1 and H2 estimated for total traits except of primary branching of panicles and paddy width due to correctness of H0: b = 1 hypothesis (Table 6). It is observed that the value of additive variance component (D) to two components of dominance variance (H1 and H2) for traits such as plant height, secondary branching of panicles is more and represents more proportion of additive effects of genes for controlling these traits. For other traits, proportions of additive effects of genes were lower which represents dominance and/or over dominance effects of genes in their genetic control.
Mean degree of dominance ![]() for traits such as plant height, primary and secondary branching of panicles is estimated lower than one and for trait such as filled grains per panicle is estimated about one and for the other traits, it is estimated more than one. Being more and lower of this index from one represents the action of over dominance and partial dominance of genes, respectively. Therefore it was concluded that traits such as tiller No., flag leaf width and length, days to maturity, panicle length, paddy length, 1000-grain weight and plant yield are influence by the effect of over dominance of genes and filled grains per panicle are influenced by the effect of complete dominance of genes and the other traits are influenced by the effect of partial dominance.
for traits such as plant height, primary and secondary branching of panicles is estimated lower than one and for trait such as filled grains per panicle is estimated about one and for the other traits, it is estimated more than one. Being more and lower of this index from one represents the action of over dominance and partial dominance of genes, respectively. Therefore it was concluded that traits such as tiller No., flag leaf width and length, days to maturity, panicle length, paddy length, 1000-grain weight and plant yield are influence by the effect of over dominance of genes and filled grains per panicle are influenced by the effect of complete dominance of genes and the other traits are influenced by the effect of partial dominance.
| Table 6: | Components of variation and stability estimates for different traits in rice |
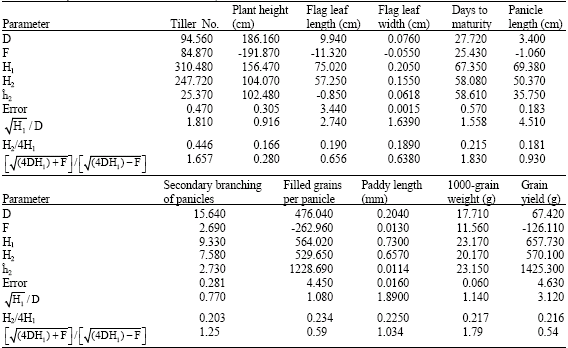 | |
In this study, index F for traits of plant height, flag leaf length and width, panicle length, primary branching of panicles, filled grains per panicle and grain yield were negative. This represents that for these traits in the studied parents, frequency of recessive alleles is more than the frequency of dominant alleles. Also for the other traits, index F was positive which represents this that in these traits, frequency of dominant alleles is more than the frequency of recessive alleles, ignoring this that dominant alleles have increasing or decreasing effects (Table 6).
Index H2/4H1 (proportion of genes with positive and negative effects in parents), for trait of tiller No. is more than 0.25 which represents that the frequency of dominant gene is more than 0.5 and for trait of filled grains per panicle, this proportion is about 0.25 which represents this that positive and negative genes are almost the equal in parents. It means their frequency is about 0.5. But for the other traits, Index H2/4H1 is lower than 0.25.
Proportion of dominant and recessive genes in the parents ![]() for trait of paddy length is about one. It means dominant and recessive genes for this trait have the same frequency and for traits such as tiller No., days to maturity, secondary branching of panicles and 1000-grain weight, this proportion is more than one. It means dominant genes are more and for the other traits, this proportion is lower than one and represents that recessive genes frequency in this traits are more than dominant genes.
for trait of paddy length is about one. It means dominant and recessive genes for this trait have the same frequency and for traits such as tiller No., days to maturity, secondary branching of panicles and 1000-grain weight, this proportion is more than one. It means dominant genes are more and for the other traits, this proportion is lower than one and represents that recessive genes frequency in this traits are more than dominant genes.
Variance and Covariance Analysis
The variance (Vr) and covariance (Wr) values on days to maturity (Table 7, Fig. 1), were related by a straight regression line with slope b. Since b = 0.80 and was not significantly (p>0.05) different from b = 1.0, the absence of non-allelic interaction and independent distribution of genes were indicated (Fig. 1).
Likewise, the intercept is negative and the actual regression line was slightly below the line of unit slope through the origin, indicating over dominance in the control of the trait.
The position of points on the regression line depicts the dominance order of the parents. Parents with most dominant genes have their points nearest the origin while the patents with most recessive genes fall farthest from origin. This graph together with Wr+Vr values (Table 7) indicated increased frequency of recessive alleles. The order of dominance was thus Sange-tarom>Daei-shastak>Dailamani>Hasani>Shastak-Mohammadi. Daei-Shastak, which gave the most days to maturity, had the most dominant genes while Shastak-Mohammadi, which gave the least, had the most recessive genes. The correlation coefficient between parental values (Yr) and parental order of dominance (Wr+Vr) was negative (r = -0.93) and significant (p<0.05), indicating that the parents containing most negative genes for the trait had the highest values of Wr and Vr. Since the value was nearly one, it confirmed the result that the dominant genes must be mostly negative, i.e., they are the beneficial genes contributing to days to maturity.
| Table 7: | Array variances (Vr) and covariances (Wr) for days to maturity and grain yield |
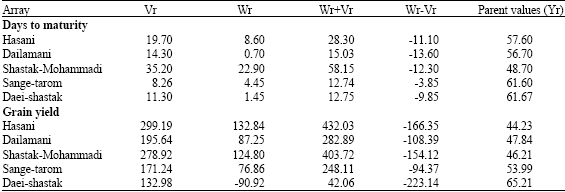 | |
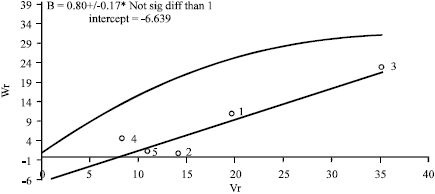 | |
| Fig. 1: | Variance (Vr), Covariance (Wr) graph on days to maturity in a five-parent incomplete diallel cross (1-Hasani, 2-Deilamani, 3-Shastak-Mohammadi, 4-Sange-tarom, 5-Daei-shastak) |
Values of Wr+Vr for grain yield indicated that increased frequency of dominant alleles. The order of dominance was thus Sange-tarom>Dailamani>Hasani>Daei-shastak>Shastak-Mohammadi (Table 7, Fig. 2). Daei-shastak, which gave the most grain yield, had the most recessive genes while Hasani, which gave the least, had the most dominant genes. The correlation coefficient between parental values (Yr) and parental order of dominance (Wr+Vr) was negative (r = -0.96) and significant (p<0.05), indicating that the parents containing most dominant genes for the trait had the highest values of Wr and Vr. Since the value was nearly one, it confirmed the result that the dominant genes must be mostly negative, i, e., they are the beneficial genes contributing to grain yield.
Results of this study imply that for days to maturity and grain yield gene action were non-additive, also parental order of dominance and the combining ability effects were almost identical for both the traits. Therefore, these results can be help in choosing suitable breeding method.
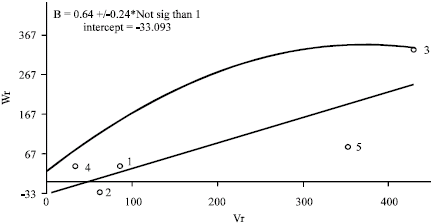 | |
| Fig. 2: | Variance (Vr), Covariance (Wr) graph on grain yield in a five-parent incomplete diallel cross (1-Hasani, 2-Deilamani, 3-Shastak-Mohammadi, 4-Sange-tarom, 5-Daei-shastak) |
DISCUSSION
Analysis of variance of traits for the studied rice varieties represents the existence of obvious difference between varieties, significant GCA for parental varieties and obvious SCA for F1 hybrids. These results show that for successful selection of genotypes in most evaluated traits in this study, there is increasingly a good genetic basis. This is a very good in traits which the proportion of additive genetic variance is high, because in this case, traits heritability for an effective selection is enough high. This case is when genetic control of influenced traits shows partial dominance effect which of additive gene effect is more than dominance effect (Singh and Chaudhary, 1985; Gravois and McNew, 1993). This study demonstrated that genetic factors were involved in traits. Present results also indicated that additive gene effects controlled plant height, primary and secondary branching of panicles. This was similar to earlier reports (Hoang and Long, 1991; Gravois and McNew, 1993). In case of grain yield and the other traits, different results were reported by different researchers. Some earlier reports revealed the predominance of additive gene effects in the control of the characters (Bui and Tuan, 1991; Singh and Kumar, 2004), while present results differed from them. On the contrary, the results presented here confirmed that non-additive gene effects was more important that additive gene effects. The different results could be attributed to different materials used. The parental lines chosen for crosses in this study were from five type lines of Iranian local cultivars.
Regarding to 89% heritability for plant height, it is a satisfactory sign which this trait will be easily transferred to next generation. Therefore regarding to this that in rice, being short-stem has been along with more yield and it is desired for breeders, to this same reason, Daei-shastak variety which shows the shortest height among the studied varieties (![]() ) and has a definite and negative GCA for plant height, when it is crossed with Hasani, Dailamani and Shastak-Mohammadi varieties (varieties which have negative GCA), there is a high probability for selection of lines with shorter height by crosses.
) and has a definite and negative GCA for plant height, when it is crossed with Hasani, Dailamani and Shastak-Mohammadi varieties (varieties which have negative GCA), there is a high probability for selection of lines with shorter height by crosses.
Selection for filled grains per panicles cannot be very successful, because this trait is controlled by dominance gene effects. But in this trait, heritability is relatively high (![]() ) and it seems that kind of gene action is additive (Table 5), on the other hand, HasanixDaei-shastak, DailamanixDaei-shastak, Shastak-MohammadixDaei-shastak and Sange-taromxDaei-shastak hybrids show a significant SCA for filled grains per panicle (Table 4). Generally for trait of filled grains per panicle, hybrids have more seed number in panicle than parental varieties which according to SCA, it is significant and positive; this can be resulted from heterosis effect which has been caused dominance gene effects.
) and it seems that kind of gene action is additive (Table 5), on the other hand, HasanixDaei-shastak, DailamanixDaei-shastak, Shastak-MohammadixDaei-shastak and Sange-taromxDaei-shastak hybrids show a significant SCA for filled grains per panicle (Table 4). Generally for trait of filled grains per panicle, hybrids have more seed number in panicle than parental varieties which according to SCA, it is significant and positive; this can be resulted from heterosis effect which has been caused dominance gene effects.
The existence of over dominance effect in genetic control of traits such as tiller No., flag leaf length and width, days to maturity, panicle length, paddy length and width, 1000-grain weight and grain yield, selection success has made difficult and ambiguous, particularly when heritability of these traits is low (Bui and Tuan, 1991; Verma and Srivastava, 2004). For trait of panicle length, positive and significant GCA of Shastak-Mohammadi variety (Table 3) is hoped that in crossing this variety with other varieties, we can obtain hybrids with a longer panicle length (Table 4). Positive and significant SCA of some hybrids such as Shastak-MohammadixDaei-Shastak, Shastak-MohammadixSange-tarom, DailamanixSange-tarom, DailamanixShastak-Mohammadi and HasanixDaei-shastak confirms that they have a longer panicle length to their parents.
The variation observed in the breeding values of parents show that in Iranian rice germplasm there exist considerable additive genetic variations to be exploited in breeding to produce inbred lines for cultural conditions. Because of the importance of additive effects, the dispersion of positive alleles for all characters between parents and the difficulty in using immediately exotic germplasm, breeding methods such as recurrent selection that emphasize GCA and is an efficient method of systematically recombining many genes could be used in rice breeding programs in order to combine high yield potential and maturity for use in Iran regions.
Quantitative inheritance of traits could complicate the selection process, analysis of combining abilities help to select suitable parents and the best hybrid combinations. Evaluating a good combination depends on both the value of GCA and SCA. A hybrid combination which has SCA value and good parental GCA values may be considered as a suitable hybrid combination. For instance, hybrid combination of Shastak-Mohammadi * Daei-shastak for grain yield (Table 5), had the positive value of SCA and at least one of the parents had the highest GCA value. Therefore, GCA and SCA effects should be taken into account when developing the strategy of the selection of genotypes for obtaining hybrid combination with high callus induction and high grain yield.
ACKNOWLEDGMENT
The researchers would like to express their appreciation to the University of Mazandaran for funding this research.
REFERENCES
- Can, N.D., S. Nakamura and T. Yoshid, 1997. Combining ability and genotype x environment interaction in early maturing grain sorghum for summer seeding. Jan. J. Crop Sci., 66: 698-705.
Direct Link - Gravois, K.A. and R.W. McNew, 1993. Combining ability and heterosis in US Southern long-grain rice. Crop Sci., 33: 83-86.
Direct Link - Griffing, B., 1956. A generalised treatment of the use of diallel crosses in quantitative inheritance. Heredity, 10: 31-50.
CrossRefDirect Link - Griffing, B., 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci., 9: 463-493.
Direct Link - Herrmann, M., 2007. A diallel analysis of various traits in winter triticale. Plant Breed., 126: 19-23.
CrossRef - Kobayashi, S., Y. Fukuta, T. Sato, M. Osaka and G.S. Khush, 2003. Molecular marker dissection of rice (Oryza sativa L.) plant architecture under temperate and tropical climates. Theor. Applied Genet., 107: 1350-1356.
CrossRef - Singh, R.K. and B.D. Chaudhary, 1985. Biometrical Methods in Quantitative Genetic Analysis. Kalyani Publishers, New Delhi, India, Pages: 318.
Direct Link - Singh, N.K. and A. Kumar, 2004. Combining ability analysis to identify suitable parents for heterotic rice hybrid breeding. Int. Rice Res. Notes, 29: 21-22.
Direct Link - Torres, E.A. and I.O. Geraldi, 2007. Partial diallel analysis of agronomic characters in rice (Oryza sativa L.). Genet. Mol. Biol., 30: 605-613.
Direct Link - Verma, O.P., 2003. Diallel analysis in rice (Oryza sativa L.) for physiological traits. Madras Agric. J., 90: 637-642.
Direct Link - Srivastava, H.K. and O.P. Verma, 2004. Genetic component and combining ability analyses in relation to heterosis for yield and associated traits using three diverse rice growing ecosystems. Field Crops Res., 88: 91-102.
CrossRef