
J.M. Al-Khayri
Department of Agricultural Biotechnology, College of Agricultural and Food Sciences, King Faisal University, P.O. Box 420, Al-Hassa 31982, Saudi Arabia
LiveDNA: 966.11152
ORCID: 0000-0001-9507-0201
Biotechnology
Year: 2010 | Volume: 9 | Issue: 4 | Page No.: 477-484







ABSTRACT
Micropropagation systems often require optimization for different genotypes by modifying culture medium. Improved regeneration in numerous plant species was attained using Coconut Water (CW) as a medium additive. The objective of the study was to evaluate callus growth and somatic embryogenesis of two commercial cultivars of date palm (Phoenix dactylifera L.) in response to CW concentrations and sterilization methods, coautoclaving or microfiltration. Calli induced from shoot tip explants, were grown on multiplication medium supplemented with CW at 0 to 20% (v/v). To induce somatic embryogenesis, callus was transferred to a hormone-free regeneration medium containing the corresponding CW concentrations. Results have shown improved callus growth and enhanced somatic embryogenesis in response to CW. Callus fresh weight and numbers of resultant embryos were directly proportional to CW concentration reaching optimum levels at 10 and 15% CW for cv. Khasab and cv. Nabout Saif, respectively. Both coautoclaving and microfiltering CW were equally effective in promoting culture growth and differentiation; however, coautoclaved CW generally gave increased responses. Furthermore, CW enrichment expedited the induction somatic embryos and increased germination percentage. Potentially this may contribute to reducing the cost of commercial micropropagation of date palm.
PDF Abstract XML References Citation
Received: May 13, 2010;
Accepted: July 23, 2010;
Published: October 21, 2010
How to cite this article
J.M. Al-Khayri, 2010. Somatic Embryogenesis of Date Palm (Phoenix dactylifera L.) Improved by Coconut Water. Biotechnology, 9: 477-484.
DOI: 10.3923/biotech.2010.477.484
URL: https://scialert.net/abstract/?doi=biotech.2010.477.484
DOI: 10.3923/biotech.2010.477.484
URL: https://scialert.net/abstract/?doi=biotech.2010.477.484
INTRODUCTION
Date palm (Phoenix dactylifera L.) is a member of the monocot family Palmae (Arecaceae) grown mainly in tropical and subtropical arid regions where it represents an important income for local farmers and dates industries (Zohary and Hopf, 2000). Because of inherent limitations associated with conventional date palm propagation using offshoots, tissue culture has become an attractive alternative for mass propagation of commercial cultivars (Al-Khayri, 2005, 2007).
Since, the earliest successful demonstrations of date palm micropropagation significant progress has been made to improve plant regeneration through adventitious organogenesis and somatic embryogenesis. Several studies were conducted to optimize the regeneration efficiency by examining the components of culture medium including physical status (Veramendi and Navarro, 1996; Taha et al., 2001; Fki et al., 2003), sucrose (Veramendi and Navarro, 1996; Taha et al., 2001), silver nitrate (Al-Khayri and Al-Bahrany, 2001, 2004), biotin and thiamine (Al-Khayri, 2001), solidifying agents (Taha et al., 2001), salt strength (Taha et al., 2001; Al-Khayri, 2003) and partial desiccation (Othmani et al., 2009). Recent studies involving cell suspension cultures have improved date palm somatic embryogenesis through manipulating the content of Abscisic Acid (ABA), Benzylaminopurine (BAP) and glutamine (Zouine and El-Hadrami, 2007). These studies have contributed to understanding the effects of a variety of culture medium additives and enhanced plant regeneration frequency. Therefore, it is imperative to investigate other culture medium components to further enhance and expedite regeneration capacity, especially for recalcitrant date palm genotypes.
In a number of plant species, regeneration improvement was achieved by augmenting culture medium with coconut water (Maddock et al., 1983; Mathias and Simpson, 1986; Al-Khayri et al., 1992; Boase et al., 1993; Jayasih and Wattimena, 1994; Mandal et al., 2002; Krug et al., 2005; Aktar et al., 2008; Nasib et al., 2008; Baskaran et al., 2008; Cheong et al., 2009; Pena-Ramirez et al., 2010; Bhattacharya et al., 2010). Coconut water contains mainly water (94%) and growth promoting substances that can influence in vitro cultures including inorganic ions, amino acids, organic acids, vitamins, sugars, sugar alcohols, lipids, nitrogenous compounds and phytohormones (Yong et al., 2009). In an effort to expedite and improve the frequency regeneration of date palm, this study was conducted to examine the effect of enriching the culture medium with coconut water on the behavior of in vitro cultures. The specific objective of the study was to evaluate callus growth and somatic embryogenesis of two date palm commercial cultivars in response to increasing concentrations of CW (0-20% v/v) added either before or after autoclaving the culture medium.
MATERIALS AND METHODS
Explant preparation and culture establishment: This research project was conducted from Jan. 1, 2008 to Dec. 30, 2009. Date palm, cvs. Khasab and Nabout Saif, offshoots of 2 to 4 years of age, were separated from mother trees, outer leaves were removed exposing the shoot tip region that were excised and immediately placed in a chilled antioxidant solution consisting of ascorbic acid and citric acid, 150 mg L-1 each, to prevent browning. The shoot tip tissues, about 8 cm in length, were surface sterilized in 70% ethanol for 1 min followed by 15 min in 1.6% w/v sodium hypochlorite (30%, v/v, Clorox, commercial bleach) containing 0.1 mL of Tween 20 (Sigma Chem Co, St. Louis, MO) per 100 mL disinfectant solution. The tissue was then rinsed with sterile distilled water four times, 30 sec each time and placed again in sterile antioxidant solution in preparation for explant excision. The tissue surrounding the shoot tips was removed until the leaf primordia were exposed and detached at the base. The shoot tip terminals, about 1 cm in length, were sectioned longitudinally into four sections. Each offshoot yielded 10 explants, 6 leaf primordia plus 4 terminal tip sections, which were used for culture initiation.
Callus initiation and maintenance: The explants were cultured on a medium consisting of MS salts (Murashige and Skoog, 1962) supplemented with (per L) 170 mg NaH2PO4, 125 mg myo-inositol, 200 mg glutamine, 5 mg thiamine-HCl, 1 mg nicotinic acid, 1 mg pyridoxine-HCl, 30 g sucrose and 7 g agar (purified Agar-agar/Gum agar) (Sigma). This basal medium was used throughout the system with some modifications made according to each stage. The callus initiation medium contained (per L) 100 mg 2,4-dichlorophenoxyacetic acid (2,4-D) (452.5 μM) and 3 mg 2-isopentenyladenine (2iP) (14.7 μM) and 1.5 g activated charcoal (acid-washed, neutralized) (Sigma). These cultures were maintained in darkness at 24±3°C for 12 weeks during which they were transferred at a 3 weeks interval. At the end of this period, resultant calli were separated and transferred to callus proliferation medium that contained (per L) 10 mg Naphthalene Acetic Acid (NAA) (53.7 μM), 30 mg 2iP (147 μM) and 1.5 g activated charcoal. These cultures were maintained for an additional 3 weeks at 24±3°C and a 16 h day-1 photoperiod with 50 μmol/m/sec Photosynthtic Photon Flux (PPF) provided by cool-white fluorescent lamps. These light and temperature regimes were used in all subsequent stages of the system. To proliferate embryogenic callus, the callus cultures were transferred to a medium containing (per L) 10 mg NAA (53.7 μM), 6 mg 2iP (29.6 μM) and 1.5 g activated charcoal. These cultures were maintained for 9 weeks during which they were transferred at a 3 weeks interval. Embryogenic calli were kept for 12 weeks on callus maintenance medium containing (per L) 10 mg NAA (53.7 μM) and 1.5 mg 2iP (7.4 μM). These callus cultures were used as a source of callus for testing the effect of coconut water. All media were adjusted to pH 5.7 with 1 N KOH and autoclaved for 15 min at 121°C and 1x105 Pa (1.1 kg cm-2).
Evaluation of the effects of coconut water: Liquid was extracted from mature coconuts, purchased from a local supermarket and heated to 70°C for 15 min, then allowed to cool and filtered by suction through buchner funnel fitted with whatman filter paper No. 1. The liquid was frozen overnight, thawed and filtered again. To test the response to Coconut Water (CW), callus maintenance medium was enriched with 0, 5, 10, 15 and 20% (v/v) of deproteinized CW. Two methods of CW sterilization were tested: the first involved adding CW to the medium and then coautoclaved, whereas the second involved adding filter-sterilized (Millipore nitrocellulose microfilter, 0.2 μm) CW to a cooled (60°C) previously-autoclaved medium. The medium was dispensed in 100 mm diameter Petri dishes at 30 mL each and inoculated with 0.5 g embryogenic callus. The cultures were maintained for 12 weeks during which they were subcultured at a 4 weeks interval. The calli were weighed to determine the effect of the treatments on callus growth. Subsequently, the callus was transferred to a hormone-free regeneration medium containing the corresponding concentrations of CW. After embryo development occurred, the resultant embryos were counted and transferred to germination.
Embryo germination and plant acclimatization: Embryos were cultured on hormone-free MS medium (without CW) and incubated for 8 weeks after which germinated embryos were transferred to rooting medium consisting of half-strength MS basal medium containing 0.1 mg L-1 (0.54 μM) NAA. For acclimatization, the plantlets were placed in beakers containing enough water to submerge the roots, kept in the culture room covered with a transparent plastic bag for 7 days and uncovered gradually during the next 7 days. Plantlets were treated with 500 mg L-1 Benlate fungicide and planted in 5 cm plastic pots containing potting mix (1 soil: 1 peat moss: 1 vermiculite). They were kept for 4 weeks in the culture room, misted three times a day and watered as needed with 100 mg L-1 of peters (20-20-20) fertilizer (Grace-Sierra). The effect on germination of somatic embryos in relation to CW treatments used during previous stages, callus growth and somatic embryo induction, was determined based on percentage of embryos that developed green shoots on the germination medium. Acclimatized potted plantlets were transferred to a greenhouse and the number of plants survived was noted.
Experimental design: To test the influence of coconut water on callus growth and subsequent somatic embryogenesis, the experiments were set up as a 2x5x2 factorial in Completely Randomized Design (CRD) comprising three main factors: genotype at 2 levels (cvs. Khasab and Nabout Saif), coconut water concentration at 5 levels (0, 5, 10, 15 and 20%, v/v) and coconut water sterilization method at 2 levels (microfiltered and coautoclaved). Each treatment consisted of 10 replications (n = 10). Data included callus weight, time of embryo initiation, number of resultant somatic embryos and percentage of embryo germination. Data were subject to Analysis of Variance (ANOVA) and the means were separated, where appropriate, using the Least Significant Difference (LSD) at 5% significance.
RESULTS
Effect of coconut water on callus growth: Callus growth was significantly affected by CW concentration and genotype as indicated by ANOVA (Table 1). In both genotypes, callus growth was promoted by the addition of as low as 5% CW. In response to increasing concentrations of CW, callus growth increased reaching maximum growth at 10 to 15% CW for cv. Khasab and cv. Nabout Saif, respectively (Fig. 1a, b). Callus growth of cv. Khasab reached maximum growth at 10% CW; whereas, cv. Nabout Saif reached maximum growth at 15% CW (Fig. 1).
| Table 1: | Analysis of variance summary of p-values of the effect of coconut water concentration and sterilization method on callus growth and embryogenesis of two date palm cultivars |
 | |
| *Significant at 5% | |
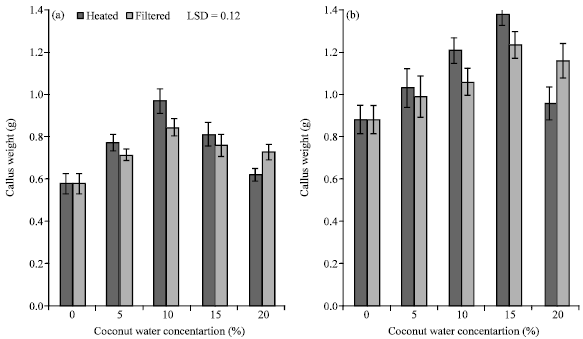 | |
| Fig. 1: | The response of callus growth, expressed in fresh weight, of two date palm cultivars, (a) Khasab and (b) Nabout Saif, to different concentrations of coconut water ranging from 0 to 20% (v/v) which was sterilized by either coautoclaving or microfiltering |
 | |
| Fig. 2: | The response of embryogenesis expressed in number of somatic embryos of two date palm cultivars, (a) Khasab and (b) Nabout Saif, to different concentrations of coconut water ranging from 0 to 20% (v/v) which was sterilized by either coautoclaving or microfiltering |
In both cultivars, 20% CW appeared to inhibit callus growth. Generally, the sterilization method of CW using coautoclaving resulted in enhanced callus proliferation (Fig. 1). An exception to this relationship was observed at the highest concentration, 20% CW, where callus growth was higher with microfiltered CW as compared to coautoclaving, especially in cv. Nabout Saif. The ANOVA, however, indicated a borderline significance of callus growth response to sterilization method (Table 1).
Effect of coconut water on somatic embryogenesis: The efficiency of somatic embryogenesis was determined based on the average number of somatic embryos produced per culture. The numbers of embryos were significantly influence by CW concentration and genotype as indicated by ANOVA (Table 1). Embryo formation was promoted in response to CW even at the lowest concentration tested, 5%. As the concentrations of CW increased, the number of resultant embryos increased, reaching a maximum 10 to 15% CW for cv. Khasab and cv. Nabout Saif, respectively (Fig. 2a, b). This relationship corresponds to that observed for callus growth. Increasing CW concentration to 20% resulted in inhibition of somatic embryogenesis. The sterilization method of coconut water showed a borderline significant effect on number of embryos (Table 1). However, like callus response, in most treatments higher embryo number formed on coautoclaved CW; except at the highest concentration, 20%, where microfiltration resulted in higher embryo number (Fig. 2).
Cultures augmented with CW commenced somatic embryo formation prior to the control cultures. Somatic embryos were visible within 8 weeks after callus was transferred to regeneration medium while the control required 12 weeks to develop embryos. Adding CW to the culture medium reduced about 30% of the time required for the regeneration stage. This improvement was exhibited by both cultivars tested regardless of CW concentration; therefore, the numbers of resultant embryos were pooled and plotted over time (Fig. 3). The pooled accumulative number of somatic embryos produced in response to CW-enrichment, were counted weekly after somatic embryogenesis occurred. The final embryo yield was determined based on the number of embryos produced 14 weeks after embryo induction. Within 12 weeks, treatments containing 10 and 15% CW produced approximately 90% of their final embryo yield; whereas, the control treatment produced only about 28% of its final embryo yield.
Embryo germination and plant survival: Medium used for somatic embryo germination was devoid of CW. However, percentage germination of somatic embryos was affected by the CW treatments used during previous stages, callus multiplication and somatic embryo induction (Fig. 4). Germination of somatic embryos obtained from treatments 0 and 5% CW showed reduced germination rate; whereas, those produced from 10 and 15% CW exhibited higher germination rate reaching above 80% in both cultivars tested.
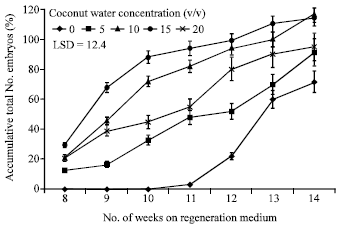 | |
| Fig. 3: | Accumulative number of somatic embryos, pooled over cultivars and coconut water sterilization methods, in response to medium enrichment with 0 to 20% (v/v) coconut water at 8 to 14 weeks after transfer of date palm callus to regeneration medium |
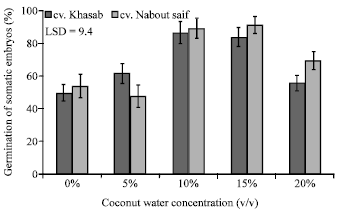 | |
| Fig. 4: | Percentage of somatic embryos germination, pooled over coconut water sterilization method, in response adding 0 to 20% (v/v) coconut water to somatic embryogenesis induction medium enrichment with counted 14 weeks after transfer of date palm callus to regeneration medium |
Upon rooting complete plants with well-developed shoot and root systems were obtained. Acclimatization conditions yielded 72 to 84% survivable in soil irrespective of the treatment. The plantlets grew normally in soil and appeared phenotypically normal.
DISCUSSION
Determination of optimum CW concentration: To stimulate in vitro responses, CW is usually added to the culture medium at concentrations ranging from 5 to 30% (v/v); however, the optimum concentration varies among plant species and must be empirically determined. For instance, optimum response in watermelon Citrullus lanatus occurred at a medium supplemented with 10% CW (Krug et al., 2005). Similarly, the best multiplication in pineapple Ananas comosus was obtained on a medium supplemented with 10% CW (Mandal et al., 2002). The addition of 10 % CW improved the number of shoots per explant in Adenia hondala (Aruna et al., 2009). This concentration of CW is routinely added to the culture medium of various plant micropropagation systems; for instance, Vanda teres (Sinha and Roy, 2004), sorghum Sorghum bicolor (Baskaran et al., 2006), blueberry Vaccinium corymbosum (Fira et al., 2008) and leaf-tea Melothria maderaspatana (Baskaran et al., 2008). In lemon grass Cymbopogon pendulus the addition of CW promoted callus induction, growth and somatic embryogenesis (Bhattacharya et al., 2010). In other plants like spinach Spinacia oleracea (Al-Khayri et al., 1992) the optimum concentration for callus growth and plant regeneration was observed at 15% CW. Whereas, adding 20% CW to the culture medium increased the number of adventitious shoots from hypocotyl segments in Spanish red cedar Cedrela odorata (Pena-Ramirez et al., 2010).
The current study has determined the optimum concentration of CW suitable for date palm in vitro cultures at 10 to 15% depending on cultivar. These levels resulted in, depending on cultivar, approximately 30 to 60% increase in callus fresh weight (Fig. 1) and about 50 to 80% increase in number of resultant somatic embryos (Fig. 2). An incremental enhanced response was observed as the concentration of CW increased to an optimum level above which an inhibitory effect was noticed in both cultivars.
In a similar study, Gnasekaran et al. (2010) evaluated the potential of CW (0, 10, 20, 30%) for micropropagation of orchid Phalenopsis violacea protocorm. They found that 10% CW stimulated proliferation of protocorm-like bodies; however, higher concentrations, 20 and 30%, tended to be inhibitory. To assess the effects of CW on micropropagation of the protocorm-like bodies of Dendrobium orchids, Puchooa (2004) supplemented the culture medium with 0-20% CW. They obtained maximum multiplication on a medium enriched with 15% CW whereas 20% was inhibitory.
In another study, Nasib et al. (2008) evaluated the effect of adding 0 to 25% CW to the culture medium of kiwifruit (Actinidia deliciosa var. deliciosa). They found that the numbers of shoots, shoot lengths and numbers of nodes were proportional to increased concentrations reaching the best responses at 20% CW whereas 25% was inhibitory. Prior to determining the optimum concentration, 10% CW was arbitrarily added to kiwifruit culture medium. Despite this using suboptimal level, Boase et al. (1993) obtained significantly improved nodes number, shoot length, petiole length and leaf area. Based on empirical experimentation involving a range of concentrations, these studies have determined the optimum concentration of CW for specific species; 10, 15 and 20% CW were reported. The present study has shown that CW concentration used in culture stages prior to the germination stage can influence the ability of resultant somatic embryos to germinate and form plantlets (Fig. 4).
Genotype-dependent response to coconut water: Variable responses of different plant genotypes to the manipulations of tissue culture factors have been well documented. For instance, the growth response to CW was found to vary among Saccharum species (Cheong et al., 2009). Date palm cultivars have been shown to respond differently to different concentrations of silver nitrate (Al-Khayri and Al-Bahrany, 2004). In the present study, callus growth (Fig. 1) and somatic embryogenesis (Fig. 2) in both date palm cultivars tested were stimulated on CW-enriched medium. However, the degree of stimulation in response to a given CW concentration and the optimum concentration varied between the two cultivars; 10 and 15% CW for cv. Khasab and cv. Nabout Saif, respectively. The results suggest that the response to CW concentrations in date palm is dependent upon cultivars. Similarly, a significant interaction between genotype and CW was also observed in wheat Triticum aestivum (Mathias and Simpson, 1986), spinach Spinacia oleracea (Al-Khayri et al., 1992) and rice Oryza sativa (Roy and Mandal, 2005).
Effect of autoclaving on coconut water activity: As the case in the current study, researchers often extract CW from mature coconuts but some reported using green coconuts. Mature coconuts usually contain higher concentrations of some active substances than green coconuts (Yong et al., 2009). Coconut water can be added either before or after autoclaving the culture medium. In the latter method, cooled autoclaved medium is augmented with microfilter-sterilized CW to avoid degradation of heat-labile active substances. However, adding CW to the medium before autoclaving eliminates the cumbersome process of microfiltration and reduces the risk of introducing contaminants during mixing and dispensing.
Various researchers routinely use either sterilization method but seldom compared in vitro culture responses to CW processed by both methods. The current study has shown that both methods were effective in enhancing callus growth (Fig. 1) and embryo formation (Fig. 2). However, the response to heated-CW generally exceeded that of filter-sterilized CW, except at 20%. This increase in response to coautoclaved CW may be due the tendency of some active substances to increase upon autoclaving CW like some amino acids and total sugars, albeit degradation of others such as glutamine, lysine and proline (Yong et al., 2009).
The exception noted at 20%, where the response to filter-sterilized exceeded that to heated-CW, may be due to reaching an inhibitory level because heated CW contains more of some active substances. Moreover, filtration may have caused the removal of more active substance rendering 20% CW of filtered sterilized CW less potent than the same concentration of coautoclaved CW. This conclusion was based on an observation made by Reddy et al. (2007) indicating that filtration reduced different nutrients including some elements, sugars and proteins. The filtration process used in their study was similar to that used in the present study, a two-stage filtration process involving whatman filter paper followed by cellulose nitrate membrane with 0.2 μm pore size. However, the CW used in their study was not deproteinized; whereas, the CW used in the present study was deproteinized twice, first by heating and second by freezing and thawing. More research is needed to fully understand the effect of heating and filtration on CW action; in particular comparative studies involving chemical analysis of CW as well as tissues of regenerated plants are necessary. More research is also needed to determine the effect of sterilization processes on individual growth factors comprising CW.
In conclusion, this study has shown that callus growth, somatic embryogenesis and embryo germination in date palm can be enhanced by supplementing the culture medium with 10 to 15% CW, depending on cultivar. Moreover, results have shown that using CW as a natural complex organic additive reduce one third of the normal time required for somatic embryo induction. This finding could contribute to shortening the lengthy duration of currently available regeneration systems which is considered a major limitation of date palm micropropagation particularly for commercial production. Moreover, the use of such a natural organic additive to replace expensive chemical sources of growth factors could contribute to reducing micropropagation cost and in turn reduces the market price per plantlet. However, more research is required to demonstrate these applications.
ACKNOWLEDGMENT
This project was supported by a research grant No. 90110 provided by the Deanship of Scientific Research, King Faisal University.
REFERENCES
- Aktar, S., K.M. Nasiruddin and K. Hossain, 2008. Effects of different media and organic additives interaction on in vitro regeneration of Dendrobium orchid. J. Agric. Rural Dev., 6: 69-74.
CrossRefDirect Link - Al-Khayri, J.M., 2001. Optimization of biotin and thiamine requirements for somatic embryogenesis of date palm (Phoenix dactylifera L.). In vitro Cell. Dev. Biol.-Plant, 37: 453-456.
CrossRefDirect Link - Al-Khayri, J.M., 2003. In vitro germination of somatic embryos in date palm: Effect of auxin concentration and strength of MS salts. Curr. Sci., 84: 680-683.
Direct Link - Al-Khayri, J.M. and A.M. Al-Bahrany, 2001. Silver nitrate and 2-isopentyladenine promote somatic embryogenesis in date palm (Phoenix dactylifera L.). Scientia Horticulturae, 89: 291-298.
CrossRefDirect Link - Al-Khayri, J.M. and A.M. Al-Bahrany, 2004. Genotype-dependent in vitro response of date palm (Phoenix dactylifera L.) cultivars to silver nitrate. Scientia Horticulturae, 99: 153-162.
CrossRefDirect Link - Al-Khayri, J.M., F.H. Huang, T.E. Morelock and T.A. Busharar, 1992. Spinach tissue culture improved with coconut water. HortScience, 27: 357-358.
Direct Link - Baskaran, P., B.R. Rajeswari and N. Jayabalan, 2006. Development of an in vitro regeneration system in sorghum (Sorghum bicolor (L.) Moench) using root transverse Thin Cell Layers (tTCLs). Turk. J. Bot., 30: 1-9.
Direct Link - Baskaran, P., P. Velayutham and N. Jayabalan, 2009. In vitro regeneration of Melothria maderaspatana via indirect organogenesis. In vitro Cell. Dev. Biol.-Plant, 45: 407-413.
CrossRefDirect Link - Boase, M.R., S. Wright and P.L. McLeay, 1993. Coconut milk enhancement of axillary shoot growth in vitro of kiwifruit. N. Z. J. Crop Hortic. Sci., 21: 171-176.
CrossRefDirect Link - Cheong, E.J., R. Mock and R. Li, 2009. Optimizing culture medium for meristem tissue culture of several Saccharum species and commercial hybrids. J. Am. Soc. Sugar Cane Technol., 29: 149-165.
Direct Link - Fki, L., R. Masmoudi, N. Drira and A. Rival, 2003. An optimised protocol for plant regeneration from embryogenic suspension cultures of date palm, Phoenix dactylifera L. cv. Deglet Nour. Plant Cell Rep., 21: 517-524.
CrossRef - Gnasekaran, P., X. Rathinam, U.R. Sinniah and S. Subramaniam, 2010. A study on the use of organic additives on the Protocorm-Like Bodies (PLBS) growth of Phalaenopsis violacea orchid. J. Phytol., 2: 29-33.
Direct Link - Jayasih, Y. and D.I.G. Wattimena, 1994. The effect of coconut water and zeolite on micropropagation of potato shoot. Acta Horticult., 369: 451-454.
Direct Link - Krug, M.G.Z., L.C.L. Stipp, A.P.M. Rodriguez and B.M.J. Mendes, 2005. In vitro organogenesis in watermelon cotyledons. Pesquisa Agropecuaria Brasileira, Brasilia, 40: 861-865.
CrossRefDirect Link - Maddock, S.E., V.A. Lancaster, R. Risiott and J. Franklin, 1983. Plant regeneration from cultured immature embryos and inflorescences of 25 cultivars of wheat (Triticum aestivum). J. Exp. Bot., 34: 915-926.
Direct Link - Mandal, A.B., M. Aparna and R. Elanchezhian, 2002. In vitro micropropagation of Ananas comosus L. (Merr.) var. Queen. J. Applied Hortic., 4: 107-112.
Direct Link - Mathias, R.J. and E.S. Simpson, 1986. The interaction of genotype and culture medium on the tissue culture responses of wheat (Triticum aestivum L. em. thell) callus. Plant Cell Tissue Organ Cult., 7: 31-37.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nasib, A., K. Ali and S. Khan, 2008. An optimized and improved method for the in vitro propagation of kiwifruit (Actinidia deliciosa) using coconut water. Pak. J. Bot., 40: 2355-2360.
Direct Link - Othmani, A., C. Bayoudh, N. Drira, M. Marrakchi and M. Trifi, 2009. Somatic embryogenesis and plant regeneration in date palm Phoenix dactylifera L., cv. Boufeggous is significantly improved by fine chopping and partial desiccation of embryogenic callus. Plant Cell Tissue Organ Culture, 97: 71-79.
CrossRefDirect Link - Pena-Ramirez, Y.J., J. Juarez-Gomez, L. Gomez-Lopez, J.L. Jeronimo-Perez, I. Garcia-Shesena, J.A. Gonzalez-Rodriguez and M.L. Robert, 2010. Multiple adventitious shoot formation in Spanish Red Cedar (Cedrela odorata L.) cultured in vitro using juvenile and mature tissues: An improved micropropagation protocol for a highly valuable tropical tree species. In vitro Cell. Dev. Biol.-Plant, 46: 149-160.
CrossRef - Puchooa, D., 2004. Comparison of different culture media for the in vitro culture of Dendrobium (Orchidaceae). Int. J. Agric. Biol., 6: 884-888.
Direct Link - Roy, B. and A.B. Mandal, 2005. Anther culture response in indica rice and variations in major agronomic characters among the androclones of a scented cultivars, kernal local. Afr. J. Biotechnol., 4: 235-240.
Direct Link - Taha, H.S., S.A. Bekheet and M.M. Saker, 2001. Factors affecting in vitro multiplication of date palm. Biol. Plant, 44: 431-433.
Direct Link - Veramendi, J. and L. Navarro, 1996. Influence of physical conditions of nutrient medium and sucrose on somatic embryogenesis of date palm. Plant Cell, Tissue Organ Culture, 45: 159-164.
CrossRef - Yong, J.W.H., L. Ge, Y.F. Ng and S.N. Tan, 2009. The chemical composition and biological properties of coconut (Cocos nucifera L.) water. Molecules, 14: 5144-5164.
CrossRefDirect Link - Zouine, J. and I. El-Hadrami, 2007. Effect of 2,4-D, glutamine and BAP on embryogenic suspension culture of date palm (Phoenix dactylifera L.). Sci. Hortic., 112: 221-226.
CrossRef