
Razia Riaz
Not Available
Muhammad Aslam Chowdhry
Not Available
Asian Journal of Plant Sciences
Year: 2003 | Volume: 2 | Issue: 10 | Page No.: 748-755







ABSTRACT
Six wheat genotypes were evaluated following a 6x6 diallel. cross to determine the heritability and variation for some physio-morphic traits under drought condition. Parental genotypes and their hybrids differed significantly for all the characters studied including leaf venation, stomatal frequency, stomata size, epidermal cell size, flag leaf area, grains per spike, 1000-kernel weight and grain yield per plant. High estimates of both broad and narrow sense heritability estimates indicated that leaf venation, stomata size, epidermal cell size, grains per spike, 1000-grain weight and grain yield per plant were highly heritable in the breeding material used in this study. A fairly high amount of genetic variation for these traits was transmitted to the offsprings and almost whole of that variation was additive in nature. Thus, single plant selection for these traits can be practiced during early generations of offsprings with high efficiency.
PDF Abstract XML References Citation
How to cite this article
Razia Riaz and Muhammad Aslam Chowdhry, 2003. Estimation of Variation and Heritability of Some Physio-morphic Traits of Wheat under Drought Condition. Asian Journal of Plant Sciences, 2: 748-755.
DOI: 10.3923/ajps.2003.748.755
URL: https://scialert.net/abstract/?doi=ajps.2003.748.755
DOI: 10.3923/ajps.2003.748.755
URL: https://scialert.net/abstract/?doi=ajps.2003.748.755
INTRODUCTION
Drought is an environmental stress of sufficient duration to produce a plant water deficit or stress which in turn causes disturbance of physiological processes. As a result crop growth rate is reduced and yield is lowered. In spite of great difficulties in breeding for high yield under drought stress genetic improvement for yield is possible and has been accomplished in many drought prone areas of the world (Smith, 1987). However, it still need further improvement. Thus, it is desirable to screen the genotypes under stress conditions to identify better adaptive ones to utilize them in future breeding programmes.
Sufficient genetic information regarding important economic characters of wheat under drought stress is not available without which the breeding strategies for drought prone areas may not prove fruitful. From breeding point of view usefulness of a character is related to its onward transmission from parents to the progeny i.e., heritability. Heritability in broad sense estimates the ratio of total genetic variance, including additive, dominance, epistatic variance, to the phenotypic variance while heritability in narrow sense estimates only the additive portion of the total phenotypic variance. Thus, narrow sense heritability is more useful because it measures the relative importance of additive portion of genetic variance, that can be transmitted to the offsprings. Characters with high heritability are easy to select during earlier segregating populations. However, most of the economic characters like grain yield and drought resistance are polygenic in nature and often are influenced by the environment and thus have low heritability. Selection for such polygenic traits becomes more difficult and requires to be delayed. Earlier studies pertaining to heritability have yielded sufficient knowledge abut the inheritance of many polygenic traits. Alexander et al. (1984) reported low heritability values (0.13 to 0.34%) for number of tillers per plant and number of grains per spike (0.335 to 0.23%) and intermediate heritability estimates (0.35 to 0.85%) for grain yield per plant.
Abid (1987) observed high heritability estimates for leaf venation and epidermal cell size. Similarly, Shahid (1987) also found medium to high heritability estimates for flag leaf venation, epidermal cell size and stomatal frequency while studying heritability of drought related morpho-physiological characters in wheat. Ahmad (1990) reported high heritability estimates for flag leaf area (72.04 to 86.5 7%), 1000-grain weight (75.10 to 94.47%) and grain yield per plant (87.11 to 97.04%), whereas moderate to high heritability estimates were found for number of grains per spike (39.57 to 80.84%).
He also observed high heritability estimates for epidermal cell size and stomatal frequency. Nayeem and Garskin (1990) reported high heritability estimates for stomatal frequency. Subliani (1997) found high narrow sense heritability for flag leaf area and 1000-kemel weight. He observed moderate estimates for stomatal frequency, leaf venation and grain yield per plant and low for grains per spike. Mahmood and Chowdhry (1999) reported high estimates of narrow sense heritability for grain and grain yield per plant. Khaliq (2000) recorded low heritability for grains per spike plant and moderate for 1000-kernel weight. Mahmood and Chowdhry (2000) reported high estimates of narrow sense heritability for grains per spike. The present studies were thus designed to study the variation for some morpho-physiological traits of wheat and to determine their heritability pattern under drought condition.
MATERIALS AND METHODS
The studies were conducted in the Department of Plant Breeding and Genetics, University of Agriculture, Faisalabad, during 1997-99. The experimental material consisted of 6 diverse wheat genotypes viz., 90-R-34, 96-R-37, Rawal 87, Rohtas 90, Chakwal 86 (Chak.86) and Kohsar 95 (Koh. 95) collected from the gene pool maintained in the department. These wheat genotypes were crossed in a 6x6 full diallel fashion during 1997-98. F1 seed along with their parents was space planted in the field using a triplicated Randomized Complete Block Design during 1998-99. Inter-row and inter-plant spacings were kept 30 and 15 cm, respectively. Seeds were sown in holes (made with the help of a dibble) at the rate of 2 seeds p!r site which were later thinned to single healthy seedling after germination. Each treatment in each replication was a single row of 5 m length comprising of approximately 30 plants. After planting the experiment, no surface irrigation was applied to maintain drought condition. All the other cultural operations including hoeing, weeding, fertilizers, etc. were carried out uniformly to reduce experimental error. Data for various physio-morphic plant traits viz., leaf venation, stomatal frequency, stomata size, epidermal cell size., flag leaf area, grains per spike, 1000-kernel weight and grain yield per plant were collected from randomly selected competitive plants. The data collected were subjected to analysis of variance according to Steel and Torrie (1984) to sort out significant differences among genotype for whole the data collected. Broad sense heritability was calculated using the variance components derived from analysis of variance table. Heritability in narrow sense was estimated by using parent-offspring regression method according to the procedures of Reeve and Robertson (1953).
Results and Discussion
A. variation
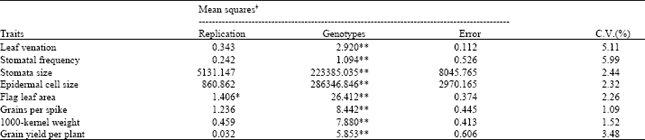
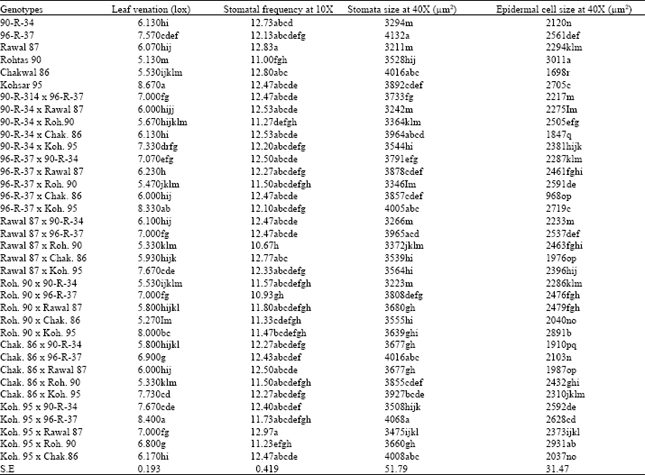
Leaf venation: The number of veins per microscopic field (10 x magnification) in the flag leaf blades of all the genotypes showed a highly significant difference (Table 1) with a moderate coefficient of variability (5.11%). Maximum number of veins (8.67) per microscopic field were recorded in Koh. 9 5 among the parental genotypes (Table 2) which differed significantly from the other five parental genotypes. Two genotypes, Roh.90 and Chak.86 produced the lowest number of veins and were statistically similar to each other.
In case of hybrids, it was observed that crosses between genotypes with higher number of veins also produced hybrids with greater leaf veins. Similarly, the number of, leaf veins also increased when a genotype with low number of veins was crossed with a genotype with higher number of veins. Therefore, maximum number of veins per microscopic field was recorded in direct and reciprocal crosses of 96-R-37 and Koh.95 (8.33 and 8.40, respectively). Both of these genotypes had highest leaf venation among parents.
Similarly, crosses showing lowest number of veins per microscopic field (viz., Roh.90 x Chak.86, Rawal.87 x Roh.90, Chak.86 x Roh.90, 96-R-37 x Rohtas 90 and Rohtas 90 x 90-R-34 involved either one or both of the parents with lower leaf venation. These results indicated the prevalence of additive genetic effects for the inheritance of leaf venation. Overall results indicated that Koh.95 and 96-R-337 were the potential genotypes to increase the leaf venation in the hybrids. Subhani (1997) also indicated Roh. 90 as a potential parent to be used in breeding for increased leaf venation in wheat.
Stomatal frequency: Stomatal frequency (number of stomata per microscopic field at 10 x magnifications) differed highly significantly among genotypes (Table 1) with a moderate (5.99%) coefficient of variability. Comparison of genotypic means for stomatal frequency (Table 2) indicated that it ranged form 10,7 to 13.0. Among the parental genotypes, Rohtas 90 had the lowest (11.0) frequency of stomata and it differed significantly from the other 5 parental genotypes whose stomatal frequency was higher and were statistically similar to each other. Highest frequency of stomata (12.8) was recorded in Rawal 87 followed by Chak.86 (12.8) and 90-R-34 (12.7).
In case of hybrids it became evident that hybrids involving Rohtas 90 showed lower stomatal frequency as compared to all other hybrids. Stomatal frequency was seemed to be under additive control which was, however, greater in some crosses and less in others. All crosses showing lowest stomatal frequency include Rohtas 90 viz., Rawal 87 x Rohtas 90 (10.7), Rohtas 90 x 96-R-37 (10.9), Koh-95 x Rohtas 90 (11.2), 90-R-34 x Rohtas 90 (11.3) and Rohtas 90 x Chak.86 (11.3). These crosses differed significantly from all the other crosses which showed higher stomatal frequency. Highest stomatal frequency (13.0) was recorded in Koh-95 x Rawal 87 hybrid. Low frequency of stomata in Roh.90 was also highlighted by Subhan (1997) and was shown to be controlled by additive gene action.
| Table 1: | Analysis of variance for some drought and yield related traits of wheat sown under drought conditions |
 | |
| Table 2: | Mean values of parental genotypes, their crosses, and statistical significance for leaf venation, stomatal frequency, stomata size and epidermal cell size |
 | |
| Means sharing common letters do not differ using DMR test at 5% P | |
Stomata size: Analysis of variance for stomata size indicated highly significant differences among genotypes with a smaller (2.44%) coefficient of variability (Table 1). Statistical comparison of genotypic means (Table 2) indicated that the parental genotypes 96-R-37 and Chak.86 had the maximum stomata size and differed significantly from the other four parental genotypes. These two genotypes were, however, statistically similar to each other. The lowest stomata size (3211 and 3294 μm2, respectively) was recorded in Rawal 87 and 90-R-34 which were statistically similar to each other but different from their parental genotypes.
A study of hybrid performance revealed that crosses between parents with higher stomata size showed larger stomata while crosses of genotypes with shorter stomata produced relatively shorter stomata. Crosses of genotypes having higher stomata size with genotypes having smaller stomata produced hybrids with intermediate stomata size or they showed an increase in stomata size towards the better parent.
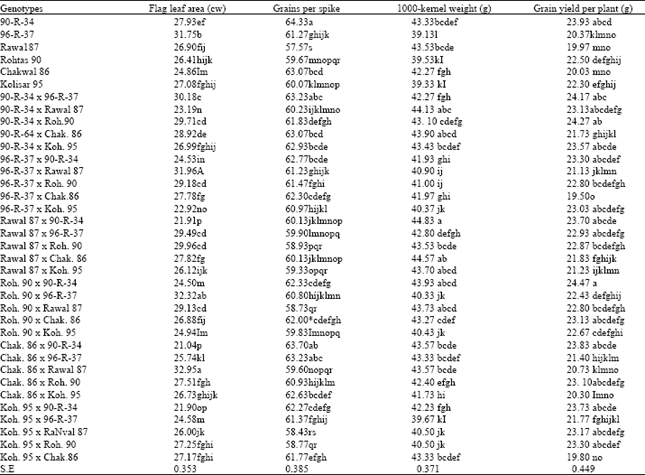
| Table 3: | Mean values of parental genotypes, their crosses and statistical significance for flag leaf area, grains per spike, 1000-kernel weight and grain yield per plant |
 | |
| Means sharing common letters do not differ using DMR test at 5% P | |
| Table 4: | Estimates ofcomponents ofvariation and heritability (h2) in broad sense (BS) and in narrow sense (NS) for some drought and yield related traits in wheat under drought |
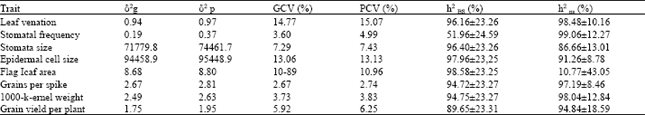 | |
| (δ2g genoty phenotypic variance, δ2p = genotypic coefficient of variability, GCV = Genotypie coefficient of variability, PGV= phenotypic coefficient of variability | |
Highest stomata size (4068 μm2) was recorded in Koh-95 x 96-R-37 hybrid closely followed by Chak.86 x 96-R-37 (4016 μm2), Koh-95 x Chak.86 (4008 μm2) and 96R-37 x Koh-95 (4005 μm2). These crosses, however, were statistically similar to each other but differed significantly from the crosses having smallest stomata size viz., Rohtas 90 x 90-R-34 (3223 μm2), 90-R-34 x Rawal 87 (3242 μm2) and Rawal 87 x 90-R-34 (3266 μm2) which were statistically similar to each other.
Epidermal cell size: Analysis of the data regarding epidermal cell size (Table 1) also revealed highly significant difference among genotypes with a lower percentage of variation (2.32%).
Statistical comparison of means (table) revealed that among the parental genotypes Rohtas 90 had'the maximum (3011 μm2) epidermal cell size which was significantly different from the other 5 parental genotypes. Similarly, lowest epidermal cell size (1698 μm2 ) was recorded in Chak.86.
Hybrids showing higher epidermal cell size involved one or.both parents with larger epidermal cell size, while hybrids between parents having smaller epidermal cell size were also having smaller epidermal cell size. Therefore, the maximum epidermal cell size (2931 μm2) was recorded in Koh-95 x Rolitas 90 hybrid followed by its direct cross i.e., Rohtas 90 x Koh-95 (2891 μm2). Both Rolitas 90 and Koh-95 were the parents having large epidermal cell size. Similarly, crosses between Chak.86 and 90-R-34 (parents with shorter epidermal cell size) produced hybrids with the shortest epidermal cell size viz., 90-R-34 x Chak.86 (1847 μm2) and Chak.86 x 90-R-34 (1910 μm2).
Flag leaf area: Comparison of genotype means after obtaining highly significant differences among them (Table 3) indicated that maximum flag leaf area among parental genotypes was recorded in 96-R-33 7 (31.75 cm2) which was significantly higher from all other parental genotypes. Similarly, the lowest flag leaf area. (24.9 cm2) was recorded in Chak.86.
Comparison of the hybrids for flag leaf area indicated the involvement of both additive and non-additive genetic effects. Flag leaf area of some hybrids showed increase towards the better parent, and in some hybrids it decreased towards the lower parent and showed intermediate values in some cases. The maximum flag leaf area (3 2.9 cm2) was recorded in Chak. 86 x Rawal 87 hybrid where both parental genotypes had smaller flag leaf area but their hybrid displayed an increase over the better parent. Similar was the case in Rohtas 90 x 96-R-37, 96-R-37 x Rawal 87, 90-R34 x Rohtas 90, 90-R-34 x Chak.86, Rawal 87 x Rohtas 90, Rawal 87 x Chak.86 and Rohtas 90 x Rawal 87 hybrids. The lowest flag leaf area (21.0 cm2) was recorded in the Chak.86 x 90-R-34 hybrid which showed over dominance towards its lower parent. This type of behaviour was found in crosses like 90-R-34 x Rawal 87, 90-R-34 x Koh-95,96-R37 x 90-R-34,90-R-34 x Koh-95, Chak.86 x 90R-34, Koh-95 x 90-R-34, Koh-95 x 96-R-37 etc. Crosses like 96-R-37 x Rohtas 90, 96-R-37 x Chak. 8 6 showed flag leaf area which was near to mid-parent value.
The genotypes differed significantly for flag leaf area but with small genotypic and phenotypic variances (Table 4). These differences arose due to only the genetic differences among the genotypes as indicated by nearly equal values of genotypic and phenotypic coefficients of variability (10.89 and 10.96%, respectively). Importance of genetic variance was also highlighted by a high and significant broad sense heritability estimate for flag leaf area (98.6%) which indicated that almost all of the genetic variance was transmitted to the offsprings for this trait. However, low heritability estimates in narrow sense pointed out that out of that genetic portion additive part was too small while a large portion included non-additive genetic variation. This suggested that selection for this trait may not be effective during early generations and must be delayed till later generations. Low heritability for flag leaf area was also observed by Borojevic (1983) while Ahmad (1990) and Subhani (1997) reported high heritability for flag leaf area.
Grains per spike: Significant differences prevailed for number of grains per spike among the 36 genotypes compared (Table 1). However, its coefficient of variation was smaller (1.09%). Statistical comparison of genotype means (Table 3) revealed that maximum number of grains per spike (64.3) was recorded in 90-R-34 followed by Chak. 86 (63.1). The lowest number (57.6) was recorded in Rawal 87.
Comparison of hybrids indicated that hybrids between 90-R-34 and 96-R-37 and other hybrids involving these genotypes produced relatively more number of grains per spike as compared to hybrids synthesizes from other parental genotypes with a few exceptions. Thus, maximum number of grains per spike was recorded in Chak.86 x 90-R-34 (63.7), 90-R-34 x 96-R-37 (63.2), Chak.86 x 96-R-37 (63.2), 90-R-34 x Chak.86 (63.1), 90-R-34 x Koh-95 (62.9) hybrids. Most of the hybrids synthesized with the genotype Rawal 87 (lowest parent) were having lower number of grains per spike.
1000-kernel weight: Comparison among genotype (Table 3) revealed that maximum 1000-kernel weight (43.5 and 4-3.3 g) was produced by Rawal 87 and 90-R-34, respectively. These two genotypes differed significantly from the other four parents but not from each other. The genotypes Rohtas 90, Koh-9 5 and 96-R-37 produced 1000-kernel weight of 39.5, 39.3 and 31.2 g, respectively and were statistically similar to each other.
Comparison of hybrids indicated that maximum 1000-kernel weight (44.8 g) was recorded in Rawal 87 x 90-R-34 hybrid. It was also noted that all hybrids involving Rawal 87 or 90-R-34 produced higher 1000-kernel weight. Similarly, all hybrids of 96-R-37 produced lower 1000-kernel weight and the lowest figure (3 9.7 g) was recorded in Koh-95 x 96-R3 7 hybrid.
Grain yield per plant: Statistical analysis of the data regarding grain yield per plant revealed significant differences among genotype Table 1 with 3.48% coefficient of variation. It was further revealed (Table 3) that the genotype 90-R-34 produced the highest (23.9 g) grain yield per plant while Rawal 87, Chak.86 and 96-R-37 produced the lowest grainyield per plant (20.0, 20.0 and 20.4 g, respectively).
In case of hybrids maximum grain yield per plant (24.5 g) was obtained in the cross Rolitas 90 x 90-R-34 followed by 90-R-34 x Rohtas 90 (24.3 g), 90-R-34 x 96-R-37 (24.2 g), Chak.86 x 90-R-3-14 (23.8 g), Koh-95 x 90-R-34 (23.7 g) and Rawal 87 x 90-R-34 (23.7 g). It was noted that all crosses producing high grain yield per plant involved 90-R-34 as one of the parents. This fact indicated the usefulness of this genotype for synthesizing high yielding genotypes under drought.
B. Heritability: It should be noted that heritability is not a constant value. The method of estimation and the procedures used by the breeder may influence its magnitude and genetic improvement obtained through selection. Furthermore, environment may also interact with the genotypic constitution to influence heritability. Therefore, the results discussed here pertains only to the genotypes under study and the environmental conditions prevailing at the experimental site. Heritability estimates for all the characters studied are presented in Table 4.
Leaf venation: Both estimates of genotypic and phenotypic variation for leaf venation were lower in magnitude with almost equal GCV and PCV percentage indicating that variation for leaf venation among genotypes was of low magnitude. Heritability estimates in broad sense and narrow sense were very high and significant depicting that all the genetic variation for leaf venation transmitted to the offsprings had a genetic base with predominant role of additive genetic causes. Thus, selection of genotypes for leaf venation can effectively be made during early generations. These results are in agreement with those of Shahid (1987) and Shahid (1993) who also reported high heritability estimates for leaf venation but are different from those of Subhani (1997) who observed low to moderate estimate of heritability for leaf venation.
Stomatal frequency: Phenotypic variance (0.37) for stomatal frequency was higher than genotypic variance (0.19) indicated the possible role of environmental influences for the variation in stomatal frequency among genotypes. Similarly genotypic coefficient of variability (3.60%) was smaller than phenotypic coefficient of variability (4.99%) for stomatal frequency. A moderate but significant estimate of broad sense heritability (51.96%) for this character also indicated that about half of the variation transmitted to the offsprings may be ascribed due to genetic causes. However, a high and significant narrow sense heritability estimate for the trait indicated that although contribution of genetic factors towards variation in stomatal frequency was less but those were all additive in nature. Thus, single plant selection may prove useful for stomatal frequency during early generations. These results are similar to those of Shahid (1-987), Ahmad (1990), Nayeem and Garskin (1990) and Shahid (1993) who also observed high heritability for stomatal frequency. Subhani (1997), however, observed a moderate narrow sense heritability estimate for stomatal frequency.
Stomata size: Genotypic variance for stomata size was smaller than the phenotypic variance. However, nearly equal values of genotypic and phenotypic coefficients of variability (7.29 and 7.43% respectively) for stomata size indicated that all of the variation for the trait was due to genetic causes. High estimates of broad sense heritability (Table 4) indicated that stomata size was a highly heritable trait and more than 96% of the genetic variability for the trait was transferred to the offsprings. Narrow sense heritability was also high and significant suggesting the predominant role of additive genetic variance for the inheritance of stomata size and indicating the effectiveness of selection for the trait during earlier generations. These findings are similar to those of Shahid (1993) who also observed high heritability for stomata size.
Epidermal cell size: It is evident from Table 4 hat epidermal cell size was also highly heritable trait among the 6 wheat genotypes studied. Although genotypic variance was smaller than phenotypic variance but almost equal percentage of genotypic and phenotypic coefficients of variability depicted that all of the variation in the epidermal cell size occurred due to genotypic differences. A significantly higher estimate of broad sense heritability (Table 4) indicated the transmission of almost all of the genetic variance to the next generation from the parental genotypes. A fairly high narrow sense heritability estimate also suggested the usefulness of additive genetic variance which was transmitted in a high quantity. Thus, the character can be easily fixed, through selection in the early generations. These results are in accordance with the findings of Abid (1987), Shahid (1987), Ahmad (1990) and Shahid (1993) who also reported high heritability for epidermal cell size in wheat.
Grains per spike: Genotypic variance (2.67) was smaller than the phenotypic variance (2.81) for grains per spike with low and similar coefficients of genotypic and phenotypic variation (2.67 and 2.74%,respectively This fact indicated that mostly genetic factors were responsible for differences in grains per spike among genotypes while environmental influences were small. Similarly, a very high and significant estimate of broad sense heritability (Table 4) displayed that this trait was highly heritable among the genotypes and more than 94% of the genetic variability for the grains per spike was transmitted to the offsprings. Similarly, narrow sense heritability estimates also highlighted that more than 97% of the genetic variation was of additive nature which suggest the usefulness of selection for grains per plant during early generations. High estimates of heritability for grains per spike have also been reported by Riaz (1990), Lu et al. (1991) and Mahmood and Chowdhry (2000).
1000-kernel weight: Very small differences in genotypic and phenotypic variances (2.49 and 2.63, respectively) and genotypic and phenotypic coefficients of variability (3.73 and 3.83%, respectively) were observed for 1000-grain weight. Which indicated a pure genetic nature of the trait. Broad sense heritability estimate was fairly high and significant displaying that almost whole of the genetic variability for the trait was transferred to the offsprings. Similarly, significantly high narrow sense heritability estimate indicated the additive nature of that genetic variance which was transmitted from parents to offsprings. This suggested that 1000-kernel weight can easily be fixed in the genotypes by selection in early generations. These findings are in line with the results of Ahmad (1990), Riaz (1990), Lu et al. (1991), Shahid (1993) and Subhani (1997) who also observed high heritability estimates for 1000-kemel weight.
Grain yield per plant: Although grain yield per plant is a highly variable character and easily influenced by the environmental factors but this character also showed very small differences in genotypic and phenotypic variances in the genotypes studied (Table 4). Phenotypoic coefficient of variability (6.25%) was slightly higher than the genotypic coefficient of variability (5.92%) displaying a small influence of other than genetic factors on the grain yield per plant. Broad sense heritability for grain yield per plant was high and significant displaying the inheritance of most of the genetic variation for the trait to the offsprings. Similar, high narrow sense heritability indicated the additive nature of most of the genetic variation transmitted from the parents to the progeny. Selection during early generations, for grain yield per plant, may thus, be practiced on single plant basis with high efficiency. High heritability for grain yield per plant has also been reported by Ahmad (1990), Lu et al. (1991), Shahid (1993) and Mahmood and Chowdhry (1999).
High estimates of both broad and narrow sense heritability estimates indicated that traits like leaf venation, stomata size, epidermal cell size, grains per spike 1000-grain weight and grain yield per plant were highly heritable in the breeding material used in this study. A fairly high amount of genetic variation for these traits was transmitted to the offsprings and almost whole of that variation was additive in nature. Thus, single plant selection for these traits can be practiced during early generations of offsprings with high efficiency.
REFERENCES
- Alexander, W.L., E.L. Smith and C. Dhanasobhari, 1984. A comparison of yield and yield components selection in Nvinterwheat. Euphytica, 33: 953-961.
CrossRefDirect Link - Mahmood, N. and M.A. Chowdhry, 1999. Inheritance of some growth parameters in bread wheat. Pak. J. Biol. Sci., 2: 781-790.
CrossRefDirect Link - Mahmood, N. and M.A. Chowdhry, 2000. Genetic performance of bread wheat genotypes for spike parameters under normal and late planting. Pak. J. Biol. Sci., 3: 440-447.
CrossRefDirect Link