
Sriani Sujiprihati
Not Available
Ghizan Bin Saleh
Not Available
Eltahir Siddig Ali
Not Available
Asian Journal of Plant Sciences
Year: 2003 | Volume: 2 | Issue: 1 | Page No.: 51-57







ABSTRACT
Maize (Zea mays L.) single-cross hybrids derived from 12 x 12 half-diallel crosses among selected inbred lines were evaluated for performance of the hybirds and to determine phenotypic correlations among traits. Varied estimates of broad-sense heritability (h2B) between locations were found for all characters, indicating the presence of genotype x location interaction on the genetic performance of the hybrids. One hundred-grain weight gave the highest h2B estimates at both locations, Field 2 and Share farm (66.4 and 93.8%, respectively), as well as in the combined analysis (80.2%). Narrow-sense heritability (h2N) estimates obtained from the variance components method were generally in agreement with those from the parent-offspring regression method, although those obtained from the latter were slightly higher for almost all traits. The h2N estimates for grain yield recorded were 54.0 and 36.7%, at field 2 and Share farm, respectively, from variance components method, while 44.7 and 34.2%, respectively, from parent-offspring regression method. Grain yield of the hybrids was most highly correlated with grain weight and grain weight/ear. Grain yield correlations with grain weight gave r values of 0.87, 0.90 and 0.91, respectively at field 2, Share farm and the locations combined, while its correlation coefficients with grain weight/ear were 0.87, 0.81 and 0.90, respectively. Flowering and maturity characters were negatively correlated with most other characters, indicating that early flowering and maturing hybrids were more favourable for grain yield and the other characters measured. Among the yield components themselves, highest correlations were shown between ear weight and grain weight/ear, where r-values recorded were 0.99, 0.96 and 0.98, respectively at field 2, Share farm and the locations combined. Promising hybrids identified in the study could be further utilized in the hybrid maize development programme.
PDF Abstract XML References Citation
How to cite this article
Sriani Sujiprihati, Ghizan Bin Saleh and Eltahir Siddig Ali, 2003. Heritability, Performance and Correlation Studies on Single Cross Hybrids of Tropical Maize. Asian Journal of Plant Sciences, 2: 51-57.
DOI: 10.3923/ajps.2003.51.57
URL: https://scialert.net/abstract/?doi=ajps.2003.51.57
DOI: 10.3923/ajps.2003.51.57
URL: https://scialert.net/abstract/?doi=ajps.2003.51.57
INTRODUCTION
Phenotypic variance is the total variance among phenotypes when grown in a range of environments of interest to the plant breeder. This variance could be portioned into genetic and environmental components to enable the breeders to design effective selection programmes (Dudley and Moll, 1969). Total genetic variance is the portion of the phenotypic variance that can be attributed to genetic differences among phenotypes. The genotype x environment interaction variance is the portion of the phenotypic variance attributed to the failure of differences among genotypes to be the same in different environments. The total genetic variance can be further sub-divided into additive genetic variance, dominance genetic variance and epistatic genetic variance (Suzuki et al., 1981).
Heritability in the broad-sense (h2B), is the ratio of the total genetic variance to the phenotypic variance. Heritability in the narrow-sense (h2N), is the ratio of additive genetic variance to the phenotypic variance. The h2N is so important to plant breeders because the effectiveness of selection depends on the additive portion of the genetic variance in relation to the total variance (Falconer and Mackay, 1996). A high h2N estimate indicates that the character investigated is simply inherited, while a low estimate shows that the character is less heritable because high number of genes are involved in the control of the character (Suzuki et al., 1981).
Daniel and Batjay (1975) estimated h2B from variance components in a diallel cross involving 11 sweet corn inbred lines for ear and plant height and number of kernel rows/ear. The estimated values were high for all characters. In a recent study on tropical maize hybrids, moderate h2B estimate was recorded for grain yield, moderate to low estimates for plant and ear height, while variation in days to silking was found least heritable (Saleh et al., 2002). From a diallel analysis on 20 yellow maize varieties, Dhillon and Singh (1977) obtained low estimates of h2N for grain yield, but higher estimates for plant and ear heights, days to silking and number of kernel rows/ear. Investigating data on eight yield components for six maize inbred lines and their F1s. Mahmoud et al. (1990) found that h2B for all traits were higher than the h2N.
Information on correlations among characters is important to determine traits to be used as selection criteria for more effective selection program. Hallauer and Miranda (1988) indicated linkage as another important factor that causes correlation in populations under random mating. They added that, indirect selection would be effective if heritability estimates of the secondary characters were greater than that of the primary one. Tan and Yap (1973); Yap and Chiow (1974) and Sim (1987) reported positive correlations between grain yield and both ear and plant heights, while maturity characters were generally negatively correlated to grain yield (Daud, 1996). Saha and Mukherjee (1985) studied a set of crosses among six heterozygous genotypes of maize, found that grain yield was correlated with ovules/ear, ovules/row, grains/ear, grains/row and 100-grains weight, but ear length was not correlated with grains/ear and 100-grains weight.
Objectives of this study were to estimate broad-sense and narrow-sense heritabilities of characters from a population of hybrids derived from a diallel cross, to evaluate performance of the hybrids, and to determine phenotypic correlations among important traits.
MATERIALS AND METHODS
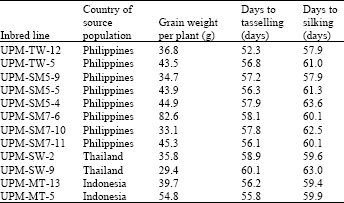
Plant materials used as parents for crosses in this study were 12 selected superior maize inbred lines (Table 1). These lines have been selected for their performance in previous evaluation trials (Sujiprihati, 1996). All possible crosses among these inbred lines were made in the diallel-crossing block. Sowing dates were adjusted to facilitate coincidence in flowering to ensure that reciprocal parental lines tasseled and silked at suitable time for crossing. In the crossing block, seeds were planted in two-row plots, 4.5 m in length and with the planting density of 1.00 X 0.30 m2. Ten crosses were made for each cross combination, to obtain ample amount of F1 hybrids seeds. The inbred lines were also concurrently selfed to obtain seeds for inbred maintenance purposes. The hand pollination procedure for the crosses and selfing followed that of Russell and Hallauer (1980). A total of 66 cross combinations were obtained through hand pollination of the inbred lines. In the evaluation trials, all hybrids were grown at each of the two locations, field 2 and Share farm, Universiti Putra Malaysia (UPM) in a randomized complete block design (RCBD) and planting density of 0.75 X 0.25 m2 under normal cultural practices (Sujiprihati, 1996). Equal amounts of seeds of a cross and its reciprocal were combined as one identity. These experiments were conducted in the period from January 1995 to December 1996.
Broad-sense heritability was estimated based on the variance components from the ANOVA.
| Table 1: | The 12 selected maize inbred lines used in diallel cross, their source populations and mean performances for grain weight per plant, days to tasselling and days to silking (after Sujiprihati, 1996) |
 | |
The formula used in the separate analysis was as suggested by Singh and Chaudhary (1977), while that used in the combined analysis was suggested by Becker (1984). Narrow-sense heritability for each location was estimated following the formula described by Rojas and Sprague (1952). Simple phenotypic correlations were computed using the formula proposed by Gomez and Gomez (1984).
RESULTS AND DISCUSSION
Broad-sense heritability estimates from field 2 data were high for 100-grains weight (66.4%), plant height (61.9%), days to tasselling (62.4%) and days to silking (60.7%). Moderate heritability estimates were shown by grain yield (31.6%), ear length (30.6%), ear diameter (33.0%), number of kernel rows/ear (48.2%), ear height (38.9%) and days to maturity (41.8%), while low estimates were shown by ear weight (19.1%), number of kernels/row (18.6%) and grain weight/ear (19.8%). For shelling percentage, the heritability estimate was negative and therefore, considered zero (Table 2).
At Share farm, however, estimates of broad-sense heritability (h2B) were high for grain yield (69.0%), ear weight (50.4%), ear length (61.7%), number of kernels/row (56.1%), 100-grains weight (93.8%), days to tasselling (81.5%), days to silking (77.8%) and days to maturity (62.6%).
| Table 2: | Broad-sense heritability estimates for 14 characters of maize hybrids. |
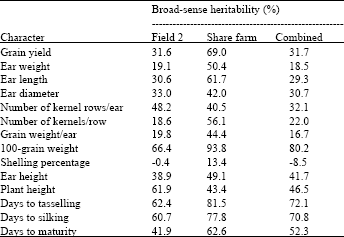 | |
Moderate h2B estimates were recorded for ear diameter, number of kernel rows, grain weight/ear, ear height and plant height, with values of 42.0, 40.5, 44.4, 49.1 and 43.4%, respectively (Table 2). Shelling percentage recorded the lowest h2B estimate (13.4%).
In the combined analysis, the highest h2B estimate was shown by 100-grains weight (80.2%), followed by days to tasselling (72.1%), days to silking (70.8%) and days to maturity (52.3%) (Table 2). Grain yield, ear length, ear diameter, number of kernel rows/ear, ear height and plant height gave moderate estimates of h2B (31.7, 29.3, 30.7, 32.1, 41.7 and 46.5%, respectively). Similar to that obtained from field 2, shelling percentage in the combined analysis also gave a negative h2B estimate and considered zero. The moderate h2B estimate for grain yield is in good agreement with that reported by Saleh et al. (2002), indicating a substantial amount of genetic variation in the control of grain yield among the hybrids evaluated. Moreover, similar high h2B estimates were also reported for plant height by Smith et al. (1998).
Results showing high estimates of broad-sense heritability for 100-grain weight, days to tasselling and days to silking at both locations indicate that, these characters were highly heritable. Selection for these characters would therefore be effective in populations generated from these hybrids. Mohd Rafii et al. (1994) and Fountain and Hallauer (1996) had reported that high heritability estimates for days to tasselling and days to silking. Moderate to low heritability estimates obtained for the other characters indicate that, if selection were to be based on them, a slower rate of progress would be expected, as compared to when characters with high heritabilities were used.
The h2N estimates for most characters revealed using the two methods, generally showed a similar trend and were in good agreement, although those from the parent-offspring regression method were generally higher (Table 3). The h2N estimates for grain yield obtained from the variance components in the combining ability analysis were low at both locations. Estimates obtained from the parent-offspring regression method were also low, although they were a little higher than those from the variance components method.
The h2N for grain yield obtained from the variance components in the combining ability analysis was estimated only at field 2 i.e., 0.3%, because negative variances were found at Share farm, making the estimation unrealistic. The estimates obtained from the parent-offspring regression method were 11.4 and 2.6%, at field 2 and Share farm, respectively.
For ear weight and grain weight/ear, heritabilities were estimated only by the parent-offspring regression method, due to the presence of negative variance components from the combining ability method. Estimates obtained were 20.9 and 21.3%, for the two characters, respectively, at field 2 and 19.4 and 23.9%, respectively, at Share farm. For ear length, h2N estimated using variance components in combining ability analysis was much lower at Share farm (0.3%), compared to that at field 2 (13.8%).
Narrow-sense heritability values for number of kernel rows/ear estimated from the variance components in the combining ability analysis (54.0% at field 2 and 36.7% at Share farm) were higher than those estimated by the parent-offspring regression (44.7% at field 2 and 34.2% at Share farm). In contrast, h2N values for ear and plant heights, estimated using the variance components in the combining ability analysis (15.1 and 89%, for the two characters, respectively at field 2, while 12.2 and 5.9%, for the same characters, respectively at Share farm) were lower than those estimated by parent-offspring regression (28.9 and 48.0%, respectively, at field 2, and 53.9 and 46.1%, respectively at Share farm). The h2N estimates for number of kernels/row and 100-grain weight were estimated only at field 2, due to negative variances at Share farm. For the same reason, h2N for shelling percentage at field 2 was only estimated by the regression method, with the value of 0.9%. At Share farm, h2N for shelling percentage obtained were higher (25.4%) when the variance components method was used, compared to that obtained by the regression method (5.5%).
For the maturity characters, h2N values were available only for days to maturity estimated by the variance components method at Share farm (6.7%), and by the regression method at both locations (31.8 and 14.3%, at field 2 and Share farm, respectively). Heritability estimates for days to tasselling and days to silking, at both locations, and days to maturity at field 2, were not obtained by the variance components method due to the negative variances.
Relatively low narrow-sense heritability estimates of the characters obtained by both methods showed that non-additive gene action was more important than the additive gene action in the genetic control of the characters investigated.
Similarly, when the broad-sense and narrow-sense heritability estimates were compared, the generally higher broad-sense heritability values showed for all characters studied indicate that non-additive gene action played a great role in the control of these characters. This result was in good agreement with that reported by Mahmoud et al. (1990). Low overall heritability estimates obtained also indicate that environmental factors had a pronounced effect relative to the genetic factors for most of the characters studied.
| Table 3: | Narrow-sense heritability estimates for 14 characters measured from variance components in the combining ability analysis, and from regression of progenies on maize inbred lines |
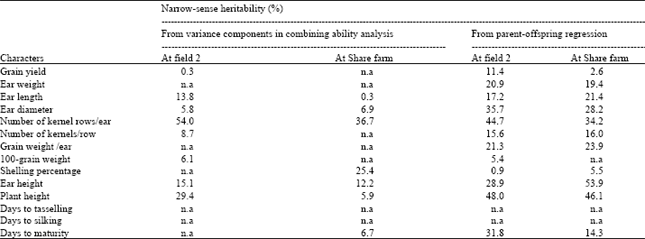 | |
| n.a = estimates not available because of negative values of the variance components | |
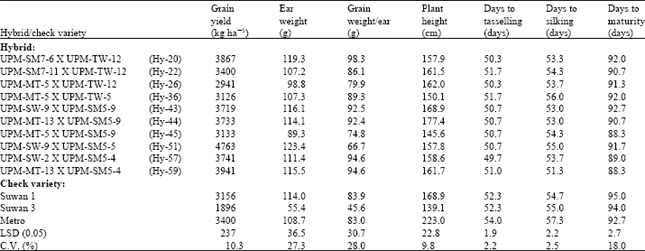
| Table 4: | Performance of the 10 top yielding maize hybrids from the diallel crosses evaluated at field 2 |
 | |
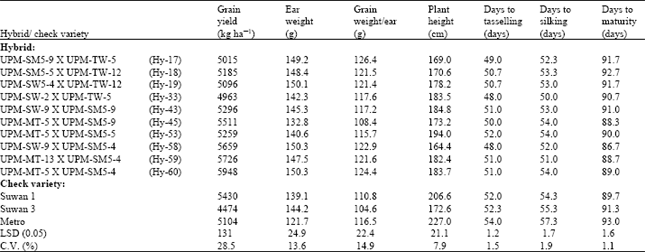
| Table 5: | Performance of the 10 top yielding maize hybrids from the diallel crosses evaluated at Share farm |
 | |
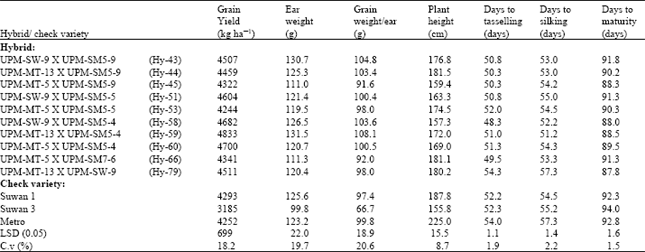
| Table 6: | Performance of the 10 top yielding maize hybrids from the diallel crosses evaluated at the two locations combined |
 | |
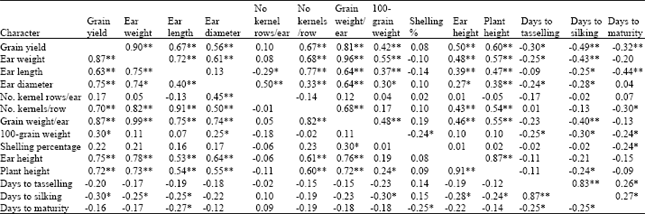
| Table 7: | Simple correlation coefficients among characters measured on maize hybrids derived from the diallel cross, at field 2 (below diagonal), and Share farm (above diagonal) |
 | |
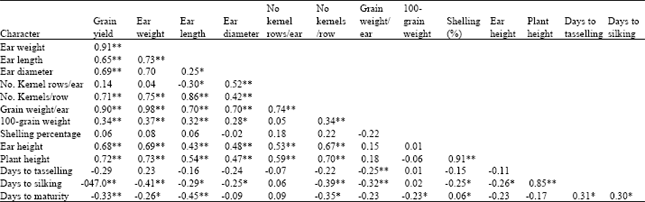
| Table 8: | Simple correlation coefficients among characters measured on maize hybrids evaluated at two locations combined |
 | |
| *Significant at P≤0.05, **Significant at P≤0.01 | |
At field 2 (Table 4), Hy-51, Hy-59 and Hy-20 gave the highest grain yields of 4763, 3941 and 3867 kg ha-1, respectively and were all higher than the three check varieties. However, Hy-60, Hy-59 and Hy-58 gave the highest grain yield at Share farm and in the combined analysis, giving 5948, 5726 and 5659 kg ha-1, respectively, at Share farm (Table 5) and 4700, 4833 and 4682 kg ha-1, respectively, in the combined analysis (Table 6). They all gave yields higher than the three check varieties. This indicates that the hybrids have high potential for utilization in the hybrid maize development programme. Similar results on grain yield potential of double cross hybrids were reported by Saleh et al. (2002), showing grain yields ranging from 3.7 to 5.7 t ha-1.
The correlation coefficients were generally consistent between the locations and locations combined. Significant positive correlations were shown between grain yield and the other characters, except number of kernel rows/ear and shelling percentage, at both locations and locations combined. The rvalues for correlations involving grain yield were 0.87, 0.63, 0.75, 0.70, 0.87 and 0.30, for ear weight, ear length, ear diameter, number of kernels/row, grain weight/ear and 100-grain weight, respectively at field 2, while 0.90, 0.67, 0.56, 0.67, 0.81 and 0.42, respectively, for the same characters, at Share farm (Table 7). In the combined analysis the same characters recorded rvalues of 0.91, 0.65, 0.69, 0.71, 0.90 and 0.34, respectively (Table 8). Correlations for grain yield were highest with ear weight (r = 0.87, 0.90 and 0.91, at field 2, Share farm and the combined analysis, respectively) and grain weight/ear (r = 0.87, 0.81 and 0.90, respectively, for the same). This study thus revealed that grain yield was primarily influenced by ear weight and grain weight/ear and secondarily by ear length, ear diameter and number of kernels/row, as direct contributing factors. Hallauer and Miranda (1988) suggested that selection may be exerted on yield components indirectly, but however, such selection would be effective if the character used possesses high heritability compared to the primary one. In addition, the correlation between them has to be substantial.
Ear and plant height were significantly correlated with grain yield, indicating that taller plants with high ear placement were better yielding compared to shorter plants with lower ear placement. This might be attributed to the high dry matter accumulation function carried out by the high number of leaves possessed in the case of tall plants.
With few exceptions, significant negative correlations were found between grain yield and the flowering and maturity characters. The correlation coefficients values were -0.20, -0.30 and -0.16 for days to tasselling, days to silking and days to maturity, respectively, at field 2, while -0.30, -0.49 and -0.32, respectively for the three characters, at Share farm. In the combined analysis, the correlation coefficients for the same characters were -0.29, -0.47 and -0.33, respectively. Correlations among the maturity characters were positive and significant and even consistent at both locations and locations combined.
Correlations between grain yield and yield components were highest between ear weight and grain weight per ear (r = 0.99, 0.96 and 0.98, respectively at field 2, Share farm and the combined analysis), but no inference could be made with regards to their use as selection criteria, due to their low heritabilities. The high correlations indicate that any increase in ear weight would simultaneously increase grain weight/ear and hence directly improve grain yield. Likewise, increase in ear length would be expected to increase number of kernels/row simultaneously. It is interesting to note that, there were positive correlations between ear length and ear diameter, at field 2 and the locations combined, although they were normally expected to be negatively correlated. Such positive relationship indicates that the favourable genes controlling these traits present in the population could be utilized for the improvement of the population sources in breeding programmes.
In conclusion grain yield was governed, to a great extent, by non-additive gene effects, and showed low heritability in the population of single cross hybrids evaluated. The strong correlations found between yield and yield components in the single cross hybrids suggested that, the yield improvement processes underwent by these high-performing hybrids in their previous breeding schemes must have been through direct or indirect improvements in the yield components of their parents.
ACKNOWLEDGMENTS
The authors gratefully acknowledge the Ministry of Science, Technology and Environment, Malaysia for the financial support on the research under the IRPA Grant No. 1-07-05-017.
REFERENCES
- Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons, New York, USA., ISBN-13: 9780471870920, Pages: 680.
Direct Link - Rafii, M.Y., G. Saleh and T.C. Yap, 1994. Response to simple and full-sib reciprocal recurrent selection in sweet corn varieties Bakti-1 and Manis Madu. J. Malaysian Applied Biol., 22: 173-180.
Direct Link - Saleh, G.B., D. Abdullah and A.R. Anuar, 2002. Performance, heterosis and heritability in selected tropical maize single, double and three-way cross hybrids. J. Agric. Sci., 138: 21-28.
CrossRefDirect Link - Singh, R.K. and B.D. Chaudhary, 1977. Biometrical Methods in Quantitative Genetic Analysis. Kalyani Publisher, New Delhi, India, ISBN: 8176633070, Pages: 318.
Direct Link - Smith, S.E., R.O. Kuehl, I.M. Ray, R. Hui and D. Soleri, 1998. Evaluation of simple methods for estimating broad-sense heritability in stands of randomly planted genotypes. Crop Sci., 38: 1125-1129.
CrossRefDirect Link