
M.S. El-Hersh
Agricultural Research Center, Department of Microbiology, Soils, Water and Environment Research Institute, Giza, Egypt
K.M. Abd El-Hai
Agricultural Research Center, Department of Leguminous and Forage Crop Diseases, Plant Pathology Research Institute, Giza, Egypt
K.M. Ghanem
Department of Environmental and Bio-Agriculture, Faculty of Agriculture, AL-Azher University, Cairo, Egypt
Asian Journal of Plant Pathology
Year: 2011 | Volume: 5 | Issue: 3 | Page No.: 102-114







ABSTRACT
Lentil is one of the most important legume crops in the world, especially in developing countries. In Egypt, root rot and wilt are the most important fungal diseases. Greenhouse and field experiments were conducted to study the effect of Molybdenum (Mo) or Cobalt (Co) on lentil pathogens (Rhizoctonia solani and Fusarium oxysporum) and nitrogen fixation compared with fungicide Rizolex T50. Lentil seeds (cvs. Giza 4 and Giza 9) were soaked 8 h in Mo or Co at concentration of 2 and 5 ppm before seeding while Rizolex T50 was used as seed-coating at 3 g kg-1 seeds. A split plot design with three replicates was used under greenhouse conditions. Results showed that damping off significantly increased in Giza 9 compared to Giza 4. While both Mo and Co decreased it in both cultivars; low level (2 ppm) was more effective. In the field, Mo and Co treatments showed significant decrease in pre and post-emergence damping off as well as dead plants (resulted from root rot and wilt). Additionally, plant height of lentil varieties showed significant increase by Mo (2 ppm) and Co (2 ppm) treatments. Different concentrations of Mo and Co were found to induce high root nodules, nitrogenase activity and nitrogen fixing bacteria. Seed yield increased significantly by Mo and Co treatments except Mo (5 ppm) on Giza 4 variety in second season. Soaking lentil seeds in Mo or Co at 2 ppm is recommended to be incorporated into the production program of lentil to decrease root rot and wilt diseases as well as improve growth and productivity.
PDF Abstract XML References Citation
Received: June 16, 2011;
Accepted: August 10, 2011;
Published: September 26, 2011
How to cite this article
M.S. El-Hersh, K.M. Abd El-Hai and K.M. Ghanem, 2011. Efficiency of Molybdenum and Cobalt Elements on the Lentil Pathogens and Nitrogen Fixation. Asian Journal of Plant Pathology, 5: 102-114.
URL: https://scialert.net/abstract/?doi=ajppaj.2011.102.114
URL: https://scialert.net/abstract/?doi=ajppaj.2011.102.114
INTRODUCTION
Lentil (Lens esculenta) is a major grain and widely distributed legume crop grown under a broad range of climates in many developing countries (Turk et al., 2004; Abd-Allah and Hashem, 2006). This crop has been grown mainly as an inexpensive source of high quality protein in human diets (Salehpour et al., 2009; Rahman et al., 2010). Lentil plants are affected by a wide range of pathogens. Fungal diseases led to decrease in productivity through infection and damage to leaves, stems, roots and pods as well as reduced marketability by discoloring seeds (Taylor et al., 2007). Fungal diseases of lentil, i.e., root rot and wilt that associated with Rhizoctonia solani and Fusarium oxysporum infections, are the most important diseases in Egypt (Hamdi et al., 2002; Morsy, 2005). These pathogens are difficult to control because of their persistency in the soil and wide host range. Thus, control of seed borne fungi and their transmission is currently limited to use of prophylactic fungicide treatments (Thomas and Sweetingham, 2003). Because of the adverse environmental effects of fungicide applications, there is a need to explore the potential of alternative strategies including the use of bioactive agent, for disease control (Jacobsen and Backman, 1993). Another strategy of control depends upon enzymatic and non-enzymatic antioxidant defense system. It allows the scavenging of reactive oxygen species and provides protection to plants from oxidative damage that associated with fungal infections (Gratao et al., 2005; Wani et al., 2008). The mechanisms that protect plant include local protection and systemic protection which could be achieved either biotically or abiotically (Goodman et al., 1986; Galal et al., 1994; Galal and Abdou, 1996). Mo is an essential trace element for most organisms as it occurs in more than sixty enzymes catalyzing diverse oxidation reduction reactions (Datta et al., 2011). Some authors studied the effects of Mo on plant health, especially in legumes. Abd El-Hai (1995) stated that Mo increases photosynthetic pigments, phosphorus and potassium contents and ultimately, support plant health. Increase in photosynthetic pigments leads to increase in carbohydrate content. Carbohydrates are the main repository of photosynthetic energy and comprise structurally polysaccharides of plant cell walls and principally cellulose, hemicelluloses and pectin that consider a barrier against plant pathogens (Hahlbrock and Scheel, 1989). They added that phenolic compounds are associated with the structural carbohydrates which play an important role in plant defense. On the other hand, Cobalt (Co) is known to promote many processes of plant growth including leaf expansion, stem and root elongation (Yu and Yang, 1979; Atta-Aly et al., 1991). Moreover, many workers explained the role of cobalt in enhancing the resistance in plants. In this respect, Zaky et al. (2002), Mazen (2004), Morsy (2005) and Mahmoud et al. (2009) stated that, cobalt can activate new proteins, chitinase and other pathogenesis-related proteins and activate many other plant defense enzymes.
The present study was designed to study the effect of Mo and Co on lentil pathogens (Rhizoctonia solani and Fusarium oxysporum) compared with the seed-coating fungicide Rizolex T50 under greenhouse and field conditions. Effects of Mo and Co on nitrogen fixation was also investigated.
MATERIALS AND METHODS
Source of lentil seeds and tested chemicals: Two lentil seed cultivars (Giza 4 and Giza 9) were obtained from Legume Crop Research Department, Field Crop Research Institute, Agriculture Research Center, Giza, Egypt. Molybdenum (Mo) in the form of molybdic acid (H2Mo4.H2O) and Cobalt (Co) in the form of cobalt sulphate (CoSO4.5H2O) were obtained from Al-Gomhoria Company, Egypt.
Isolation, purification and identification of pathogenic fungi: The pathogenic fungi used in this study were isolated from lentil plants showing symptoms of root rot and wilt diseases in 2007/2008 winter season. The infected plant parts were cut into small pieces and washed with sterilized water, then placed on PDA medium in petri-dishes supplemented with streptomycin sulfate (at the rate of 100 μg mL-1). The growing fungi were purified and identified by spore morphology and pigmentation according to Booth (1977) and Sneh et al. (1992) for Fusarium oxysporum and Rhizoctonia solani, respectively.
Preparation of fungal inocula: Inocula of the most aggressive isolates of F. oxysporum and R. solani were prepared using sorghum-course sand-water (2:1:2 v/v) medium. The ingredients were mixed, bottled and autoclaved for 30 min at 121°C and 1.5 air pressure. The sterilized medium was inoculated using agar discs, obtained from the periphery of 7 days old colony of each isolated fungi. The inoculated media were incubated at 25±1°C for 15 days, then used for soil infestation in greenhouse experiment for studying the pathogenecity test.
Greenhouse experiment: Pot experiment was carried out for testing the response of two lentil cultivars to tested chemical under soil infested with causal pathogens. Disinfected soil was sterilized by 5% formalin and thoroughly mixed separately with the previously prepared fungal inocula at the rate of 3% of soil weight (w/w) placed in plastic pots (30 cm diameter), irrigated twice at 3 day intervals before sowing to enhance fungal growth. Lentil seeds were soaked per 8 hours in Mo or Co at concentration of 2 and 5 ppm and sown in pots at the rate of 20 seeds/pot. Untreated seeds were sown as check treatment and also seeds coating with Rizolex-T50 (3 g kg-1 seeds) were sown as a fungicide treatment. Three pots were used as replicates for each particular treatment. Percentage of pre-and post-emergence damping-off were recorded at 15 and 30 days after sowing.
Field experiment: Two field experiments were carried out during 2008/2009 and 2009/2010 seasons in naturally infested soil of Tag EL-Ezz Agriculture Research Station Farm, Dakahlia Governorate, Egypt. Lentil seeds (Giza 4 and Giza 9) either treated with the same previous treatments in the greenhouse experiment or untreated ones were used. Seeds were sown on November, 25th in the first season and 15th in the second one. Two seeds/hill were sown with 5 cm apart between hills. A split plot design with three replicates was used. The main plots were varieties of lentil while sub-plots were located to treatments. The area of each sub-plot was 10.5 m2 (3.5x3 m) consisting 5 rows. Pre- and post-emergence damping-off as well as dead plants (resulted from root rot and/or wilt) were recorded at 15, 30 and 60 days from sowing, respectively.
Nodulation status, nitrogenase activity and microbial counts: Samples of lentil roots were collected after 70 days from sowing in the second season only for the determination of number and dry weight (at 70°C until constant weight) of nodules. Nitrogenase activity was determined according to Lethbridage et al. (1982). The counting of total bacteria, nitrogen fixers and total fungi in the rhizospheric soil of lentil plants was performed using the media of Bridson (1978), Watanabe and Barraquio (1979) and Martin (1950), respectively.
Growth, yield and seed quality: At harvest stage (150 days from sowing), plant height (cm), number of branches, number of pods plant-1, weight of 1000 seeds (g) and seed yield (kg fed-1) were recorded. Lentil seeds were dried at 70°C for 48 h., grounded and analyzed for phosphorus content (Chapman and Pratt, 1965) and total nitrogen by semi-micro-Kjeldahl (Pregl, 1945). The protein percentage was calculated by multiplying N% by 6.25.
Statistical analysis: All data were subjected to the proper statistical analysis of variance, mean values of treatments were differentiated using Least Significant Difference (LSD). Simple linear correlation coefficient (r) was used to examine the relationships between individual properties. The statistical analysis software; CoStat v6.4 was used.
RESULTS
Greenhouse studies: Pre and post-emergence damping off in lentil varieties, i.e., Giza 4 and Giza 9 caused by F. oxysporum and R. solani were studied in pot experiments. Table 1 indicated that pre and post emergence damping off ratios were significantly increased in Giza 9 compared to Giza 4. Damping off either in pre or post-emergence showed to be significantly decreased in Rizolex compared to Mo (2 and 5 ppm) and Co (5 ppm) treatments. However, no significant differences in pre-emergence damping off under soil infested by F. oxysporum obtained by Co at 2 ppm (7.3%) in Giza 4 compared to Rizolex (6%). Interestingly, damping off in varieties of lentil either in low concentration (2 ppm) of Mo or Co significantly decreased compared to their higher concentrations (5 ppm).
| Table 1: | Pre- and post-emergence damping off caused by Fusarium oxysporum and Rhizoctonia solani under greenhouse conditions |
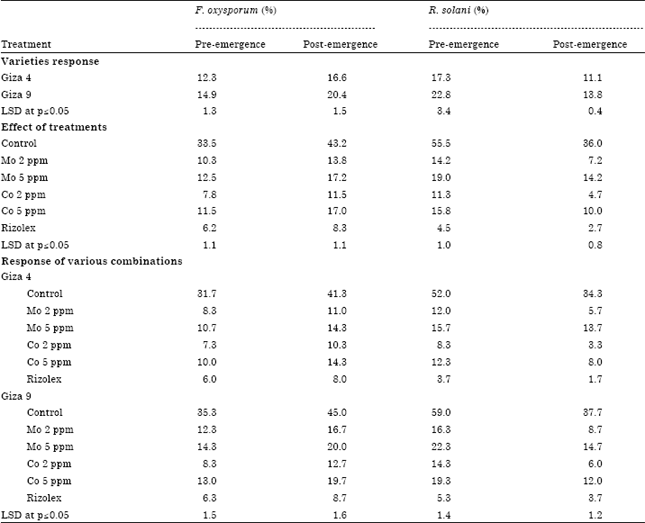 | |
| Table 2: | Pre and post-emergence damping off as well as dead plants caused during two successive growing seasons under field conditions |
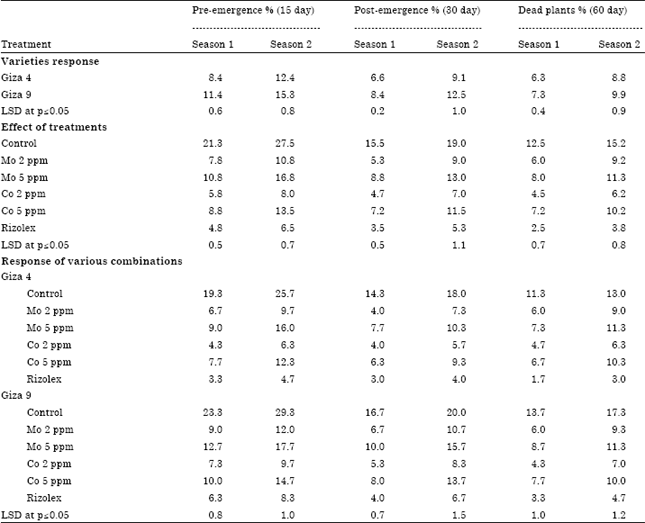 | |
Field studies: Table 2 presents the percentage of pre and post-emergence damping off and dead plants that occurred during two successive growing seasons under field conditions. Damping off percentage significantly increased in Giza 9 compared to Giza 4 in both seasons. Significant differences (p≤0.05) also have been obtained among the treatments. Pre and post-emergence damping off showed to be significantly decreased in Mo and Co treatments compared to control. Moreover, Mo (2 ppm) and Co (2 ppm) showed significant increase in plant height of lentil varieties compared to control. Giza 9 showed significant increase in plant height (48.9 and 53.6 cms) compared to Giza 4 (41.8 and 45.7 cms) during first and second season, respectively (Table 3). However, no significant difference (p≤0.05) in branches number has been obtained.
Data presented in Fig. 1 show number and dry weight of nodules and nitrogenase activity as response to Mo and Co treatments. Generally, Mo and Co at their concentrations (2 and 5 ppm) increased the aforementioned parameters. However, low concentration (2 ppm) of Mo or Co showed to be great inducers for tested parameters compared to the high concentrations (5 ppm). The highest values of nodules number (16 nodule) in Giza 4 and (15 nodule) in Giza 9 occurred under the application of Co (2 ppm).
| Table 3: | Growth parameters of 70 days old lentil plants as affected by different combinations of varieties and tested elements |
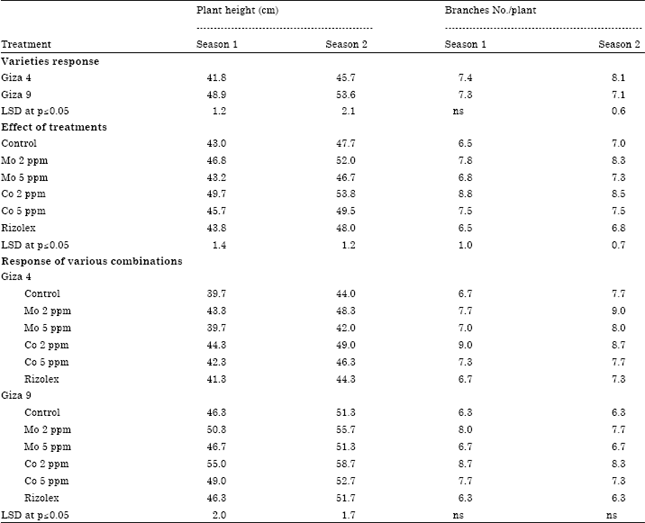 | |
Moreover, nitrogenase activity was more than 120 MnC2H4/g dry nodules/h in Giza 4 by low treatment of Mo and Co. Furthermore, data obtained (Fig. 2) indicated that the prevalence of Log bacterial count during Mo and Co treatments, as well as, the nitrogen fixing bacteria showed increasing trends as compared to control. Interestingly, low concentrations of Mo (2 ppm) and Co (2 ppm) were stimulating for the numbers of total bacterial count (from 7:7.5 Log. count/g soil) and nitrogen fixing bacteria (5:5.8 Log. Count/g soil).
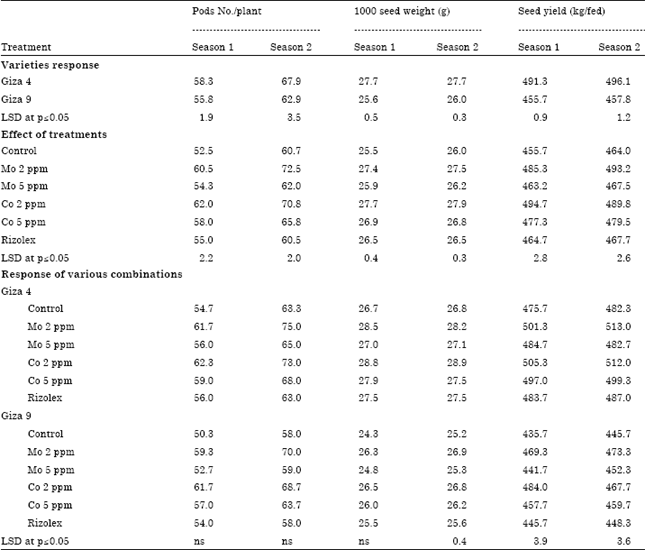
Table 4 indicates that there is a significant difference (p≤0.05) in varieties response to different treatments. Giza 4 yielded the highest number of pods per plant (58.3 and 67.9) in the first and second season, respectively. Also, it gives the highest 1000 seed weight (27.7 g) in both seasons and seed yield/ fed (491.3 and 496.1 kg fed-1). However, no significant difference (p≤0.05) in pod numbers/plant among the treatments. Likewise, the 1000 seed weight (g) of lentil showed no significant difference in season one. As well as, no significant difference has been determined in seed weights between Mo (5 ppm) or Co (5 ppm) and Rizolex. Interestingly, Mo and Co at concentration of 2 ppm showed to be more stimulating to seed weight and seed yield compared to their higher concentration (5 ppm). On the other hand, the quality of lentil seeds was evaluated according to their content of nitrogen, protein and phosphorus.
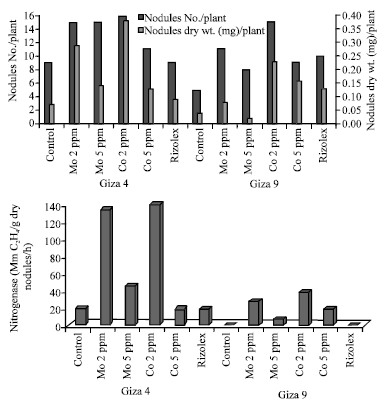 | |
| Fig. 1: | Nodulation status and nitrogenase activity of lentil nodules as affected by different treatments after 70 days of growth |
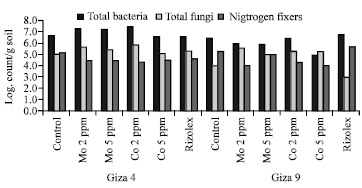 | |
| Fig. 2: | The population of some microbial groups in the rhizosphere of lentil plants as affected by different treatments after 70 days of growth |
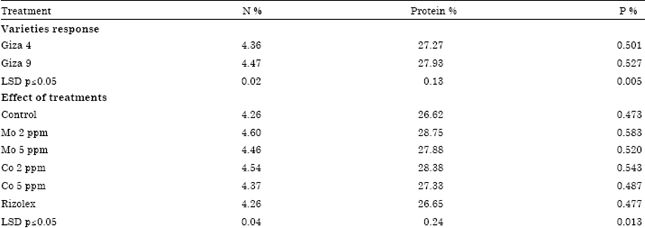
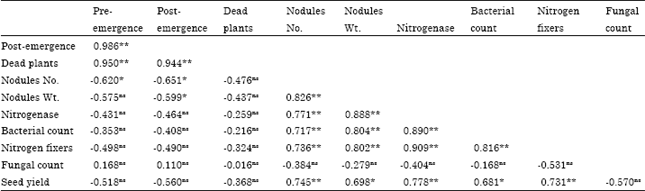
Table 5 revealed that Giza 9 was the best in seed quality (27.93% protein and 0.527% P). Moreover, any concentration of elements used increased these parameters significantly. Mo at 2 ppm followed by Co at 2 ppm was more effective. While the interaction between variety and treatments showed no significant effect. The correlation between parameters was evaluated (Table 6) and the summarized data showed that there is a significant correlation (p≤0.05) between pre and post-emergence (0.986) as well as pre, post-emergence and dead plants. Also there is a significant correlation between nodules number and nodules weight (0.826), nodules weight and nitrogenase activity (0.888) and nitrogenase activity with total count (0.890) and nitrogen fixing bacteria (0.909). Moreover, there is a significant correlation between seed yield with nodules number (0.745), nitrogenase activity (0.778), total bacterial count (0.681) and nitrogen fixing bacteria (0.731).
| Table 4: | Yield of lentil as affected by different combinations of varieties and tested elements |
 | |
| Table 5: | Quality of lentil seeds recovered from the second season as affected by different combinations of varieties and the tested elements |
 | |
 | |
| Table 6: | Correlation between every two pairs of main parameters |
 | |
| nsNot significant, *Significant at p≤0.05, **Significant at p≤0.01 | |
DISCUSSION
Results indicated that Mo and Co at 2 ppm concentrations showed significant decrease in pre and post-emergence damping off both in in vitro and in vivo conditions compared to control. The role of Mo in decreasing damping off may be due to increasing the level of cytokinin, consequently enhancing the total phenols, calcium content and activity of catechol oxidase which protect plants against pathogens infection (Wahdan, 1991; Chowdhury, 2003). Besides, Co concentrations played an important role in reduction of damping off and wilt diseases. Our results agree with Abd El-Kareem et al. (1993), Zaky et al. (2002) and Mahmoud et al. (2009). They explained the role of cobalt in inducing the resistance in plants by activating new proteins and several enzymes that may have a role in disease resistance. In addition, the difference in damping off ratios as well as growth and yield between the two varieties may be due to the genetic structure that plays a role in these parameters (Khan et al., 2001; Akem et al., 2006; Panahyan-e-Kivi et al., 2009). Moreover, Mo and Co may have inducer effect on the antioxidant enzymes such as glutathione reductase that produced under metal stress (Cardoso et al., 2005) and reduced metal toxicity as well as protect plants from pathogens. Results obtained also showed significant increase in plant height by Mo and Co at concentration of 2 ppm compared with control. Mo increased plant height, photosynthetic pigments, mineral content and number of pods and ultimately, support plant health (Abd El-Hai, 1995; Datta et al., 2011). Because increasing of photosynthetic pigments led to increases carbohydrate content. Carbohydrates are the main repository of photosynthetic energy, they comprise structural polysaccharides of plant cell walls and principally cellulose, hemicelluloses and pectin that consider the barrier against plant pathogen. Also phenolic compounds are associated with the structural carbohydrates which play an important role in plant defense (Hahlbrock and Scheel, 1989). Likewise, cobalt is known to promote many processes of plant growth including leaf expansion, stem and root elongation (Lau and Yang, 1976; Yu and Yang, 1979; Atta-Aly et al., 1991). Moreover, Cobalt can be activated new proteins, as chitinase and other pathogenesis-related proteins and activate many of enzymes which have a role in disease resistance (Zaky et al., 2002; Mazen, 2004; Morsy, 2005; Mahmoud et al., 2009).
On the other hand, the stimulatory effects of Mo and Co in nodules number, nodules dry weight and nitrogenase activity were described by Hashimato and Yamasaki (1976) and Sutcliffe and Baker (1981). They stated that Mo is required for organisms utilizing atmospheric nitrogen, also, Mo is believed to serve as an activator of enzymes involved in nitrogen fixation process. Regard to Co treatments, data showed that Co increased number and dry weight of nodules and nitrogenase activity. This is supported by Mathur et al. (2006). Although, low concentration of Mo and Co was found to be more inducer for nitrogenase activity, these results may be due to the effect of higher concentration of Mo or Co on rhizobacteria. Molybdenum is an essential constituent of two important enzymes involved in the assimilation of nitrogen, namely nitrate reductase and nitrogenase. Nitrate reductase is a molybdoflavo-protein and is present in leaves and roots, probably in the cytoplasm. Nitrogenase, the enzyme responsible for the reduction of dinitrogen, contains of two iron-sulphur proteins, the larger one of which contains molybdenum and is an iron-molybdenum protein and the smaller one is an iron protein (Lauchli et al., 1983; Siddiky et al., 2007). While, cobalt is as the metal component of coenzyme B12 which required for leghaemoglobin formation. In addition to other important reactions in nitrogen fixation dependent on either vitamin B12 or the coenzyme form (Marschner, 1986). Moreover, rhizobacteria was found to reduce the toxicity of heavy metals and produce growth promoting substances (Gupta et al., 2004; Wani et al., 2008). The results also, indicated that the genetic structure may play a vital role in the significant differences of varieties response to the treatments. Obtained results also indicated that, Mo and Co at low concentration (2 ppm) showed a great inducing effect on seed weight and seed yield compared to the higher concentrations. Xia and Xiong (1991) pointed out that Mo stimulated N uptake and content, as well as, Mo increased cytokinin which induced lateral bud development (Tucker and Mansfield, 1973) with subsequent growth of lateral shoots. On the other hand, lentil seeds were chemically evaluated for their content of nitrogen, protein and phosphorus. Results indicated that no significant difference has been obtained.
The positive correlation among the tested parameters may explain the obvious increment of lentil yield and its growth, in which, the increase in total bacterial count and nitrogen fixing bacteria led to increase in nodules number. In turn, increase in nitrogenase activity consequently induce nitrogen fixation. Nitrogen is usually the fourth most abundant element in plants, following carbon, oxygen and hydrogen. It has an essential role as a constituent of proteins, nucleic acid, chlorophyll and growth hormones (Engelhard, 1993). The importance of Mo and Co may be due to its effects as co-factor for some enzymes that play a vital role in protection of plants against pathogens. As well as its activators for metallo-enzymes that participate in the plant metabolism and nitrogen fixation process (Ahmad et al., 2002).
CONCLUSION
Finally, it was found that Mo and Co at low level (2 ppm) decreased damping off and wilt diseases and enhanced growth and yield of lentil. So, it could be recommended to use Mo and Co at 2 ppm for seed soaking of lentil to overcome the injurious effects of pathogens. In addition to disease suppression, Mo and Co have the potential to enhance growth, nitrogenase activity and to maximize the yield and seed quality.
REFERENCES
- Ahmad, M., M. Hussain and M. Shafique, 2002. Important macro and microelements in chickpea and lentil. Nucleus, 39: 101-105.
Direct Link - Akem, C., M. Bellar and B. Bayaa, 2006. Comparative growth and pathogenicity of geographical isolates of Sclerotinia sclerotiorum on lentil genotypes. Plant Pathol. J., 5: 67-71.
CrossRefDirect Link - Cardoso, P.F., P.L. Gratao, R.A. Gomes-Junior, L.O. Medici and R.A. Azevedo, 2005. Response of Crotalaria juncea to nickel exposure. Braz. J. Plant Physiol., 17: 267-272.
CrossRefDirect Link - Chowdhury, A.K., 2003. Control of Sclerotium blight of groundnut by growth substances. Crop Res., 25: 355-359.
Direct Link - Gratao, P.L., A. Polle, P.J. Lea and R.A. Azevedo, 2005. Making the life of heavy metal-stressed plants a little easier. Funct. Plant Biol., 32: 481-494.
CrossRefDirect Link - Gupta, D.K., U.N. Rai, S. Sinha, R.D. Tripathi, B.D. Nautiyal, P. Rai and M. Inouhe, 2004. Role of Rhizobium (CA-1) inoculation in increasing growth and metal accumulation in Cicer arietinum L. growing under fly-ash stress condition. Bull. Environ. Contam. Toxicol., 73: 424-431.
PubMed - Hahlbrock, K. and D. Scheel, 1989. Physiology and molecular biology of phenylpropanoid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol., 40: 347-369.
CrossRefDirect Link - Hashimato, K. and S. Yamasaki, 1976. Effects of molybdenum application on the yield, nitrogen nutrition and nodule development of soybeans. Soil Sci. Plant Nutr., 22: 435-443.
Direct Link - Jacobsen B.J. and P.A. Backman, 1993. Biological and cultural plant disease controls: Alternatives and supplements to chemicals in IPM systems. Plant Dis., 77: 311-315.
Direct Link - Lau, O.L. and S.F. Yang, 1976. Inhibition of ethylene production by cobaltous ion. Plant Physiol., 58: 114-117.
CrossRefDirect Link - Khan, A., M. Rahim and A. Khan, 2001. Performance of exotic lentil varieties under rainfed conditions in Mingora (NWFP) Pakistan. J. Biological Sci., 1: 343-344.
CrossRefDirect Link - Martin, J.P., 1950. Use of acid, rose bengal and streptomycin in the plate method for estimating soil fungi. Soil Sci., 69: 215-232.
Direct Link - Mathur, N., J. Singh, S. Bohra, A. Bohra and A. Vyas, 2006. Effect of soil compaction, potassium and cobalt on growth and yield of moth bean. Int. J. Soil Sci., 1: 269-271.
CrossRefDirect Link - Morsy, K.M.M., 2005. Induced resistance against damping-off, root rot and wilt diseases of lentil. Egypt. J. Phytopathol., 33: 53-63.
Direct Link - Panahyan-e-Kivi, M., A. Ebadi, A. Tobeh and Sh. Jamaati-e-Somarin, 2009. Evaluation of yield and yield components of lentil genotypes under drought stress. Res. J. Environ. Sci., 3: 456-460.
CrossRefDirect Link - Rahman, T., A.U. Ahmed, M.R. Islam and M.I. Hosen, 2010. Physiological study and both in vitro and in vivo antifungal activities against Stemphylium botryosum causing Stemphylium blight disease in lentil (Lens culinaris). Plant Pathol. J., 9: 179-187.
CrossRefDirect Link - Salehpour, M., A. Ebadi, M. Izadi and Sh. Jamaati-e-Somarin, 2009. Evaluation of water stress and nitrogen fertilizer effects on relative water content, membrane stability index, chlorophyll and some other traits of lentils (Lens culinaris L.) under hydroponics conditions. Res. J. Environ. Sci., 3: 103-109.
CrossRefDirect Link - Siddiky, M.A., N.K. Halder, Z. Islam, R.A. Begam and M.M. Masud, 2007. Performance of brinjal as influenced by boron and molybdenum. Asian J. Plant Sci., 6: 389-393.
CrossRefDirect Link - Thomas, G.J. and M.W. Sweetingham, 2003. Fungicide seed treatments reduce seed transmission and severity of lupin anthracnose caused by Colletotrichum gloeosporioides. Aust. Plant Pathol., 32: 39-46.
CrossRefDirect Link - Tucker, D.J. and T.A. Mansfield, 1973. Apical dominanace in Xanthium strumarium. J. Exp. Bot., 24: 731-740.
CrossRefDirect Link - Turk, M.A., A.R.M. Tawaha and K.D. Lee, 2004. Seed germination and seedling growth of three lentil cultivars under moisture stress. Asian J. Plant Sci., 3: 394-397.
CrossRefDirect Link - Wani, P.A., M.S. Khan and A. Zaidi, 2008. Effect of metal-tolerant plant growth-promoting Rhizobium on the performance of pea grown in metal-amended soil. Arch. Environ. Contam. Toxicol., 55: 33-42.
CrossRef - Watanabe, I. and W.L. Barraquio, 1979. Low levels of fixed nitrogen required for isolation of free-living N2-fixing organisms from rice roots. Nature, 277: 565-566.
CrossRefDirect Link - Xia, M.Z. and F.Q. Xiong, 1991. Interaction of molybdenum, phosphorus and potassium on yield in Vicia faba. J. Agric. Sci. Cambridge, 117: 85-89.
CrossRef - Yu, Y.B. and S.F. Yang, 1979. Auxin-induced ethylene production and its inhibition by aminoethoxyvinyiglycine and cobalt ion. Plant Physiol., 64: 1074-1077.
CrossRefDirect Link