
K.D. Adjata
Laboratoire de Virologie et de Biotechnologie Vegetales (LVBV), Ecole Superieure d`Agronomie, Universite de Lome, B.P. 1515, Lome, Togo
E. Muller
CIRAD-UMR BGPI (Biologie et Genetique des Interactions Plantes-Parasites) TA41/K, Campus International de Baillarguet, 34398 Montpellier Cedex 5, France
M. Aziadekey
Laboratoire de Virologie et de Biotechnologie Vegetales (LVBV), Ecole Superieure d`Agronomie, Universite de Lome, B.P. 1515, Lome, Togo
Y.M.D. Gumedzoe
Laboratoire de Virologie et de Biotechnologie Vegetales (LVBV), Ecole Superieure d`Agronomie, Universite de Lome, B.P. 1515, Lome, Togo
M. Peterschmitt
CIRAD-UMR BGPI (Biologie et Genetique des Interactions Plantes-Parasites) TA41/K, Campus International de Baillarguet, 34398 Montpellier Cedex 5, France
American Journal of Plant Physiology
Year: 2008 | Volume: 3 | Issue: 2 | Page No.: 73-80







ABSTRACT
Cassava is infected by numerous Begomoviruses in Africa and India that cause devastating losses to poor farmers. In order to identify viruses responsible for the disease and characterize them, surveys were conducted in all cassava production zones in Togo. The symptom severity of these viral diseases was recorded. Foliar samples from cassava and other infected plants were collected and analysed by PCR. The results obtained reveal that the severity of the symptoms varies from one locality to another, some more severely cassava infected plants than others. The percentage of the cassava plants presenting typical mosaic symptoms varies from 55 to 85% and the percentage of cassava plants severely infected (score 4 or 5) varies from 4.9 to 36%. Molecular analyses by PCR of the viral DNA extracted from the diseased samples using specific primers revealed for the first time that African cassava mosaic virus (ACMV) is not the only Begomovirus responsible of cassava mosaic disease in Togo, but two other Begomoviruses, East African cassava mosaic virus (EACMV) and Indian cassava mosaic virus (ICMV) are present in Togo. Their incidence rates are 73.59, 44.62 and 4.01%, respectively for ACMV, EACMV and ICMV. Mixed infections were also identified at the rates of 39.45% in the case of (ACMV + EACMV), 1.72% for ACMV + ICMV and 1.29% when the three viruses were combined (ACMV + EACMV + ICMV).
PDF Abstract XML References Citation
How to cite this article
K.D. Adjata, E. Muller, M. Aziadekey, Y.M.D. Gumedzoe and M. Peterschmitt, 2008. Incidence of Cassava Viral Diseases and First Identification of East African cassava mosaic virus and Indian cassava mosaic virus by PCR in Cassava (Manihot esculenta Crantz) Fields in Togo. American Journal of Plant Physiology, 3: 73-80.
DOI: 10.3923/ajpp.2008.73.80
URL: https://scialert.net/abstract/?doi=ajpp.2008.73.80
DOI: 10.3923/ajpp.2008.73.80
URL: https://scialert.net/abstract/?doi=ajpp.2008.73.80
INTRODUCTION
Cassava (Manihot esculenta Crantz) cultivated primarily for its tuberous roots, is the third source of carbohydrate in the world; its leaves are also consumed as vegetables in many region in Africa and constitute a source of proteins, vitamins and mineral salts (Dixon et al., 1992; Legg and Fauquet, 2004). This root plant which constitutes the main food crop in sub-Saharan Africa ensures the staple food of more than 200 million people through the tropics. Cassava plays thus an important role in food security. Unfortunately the production of cassava underwent considerable falls these last years and particularly in Togo. These falls are due to many biotic and abiotic factors. Among the biotic factors, several viral diseases have been reported (Atiri et al., 2004; Ndunguru et al., 2005). They include: Cassava Mosaic Disease (CMD) caused by African cassava mosaic Begomovirus; cassava brown streak disease caused by Cassava brown streak virus (CBSV) reported along the coast of Kenya (Bock, 1994); Tobacco leaf curl virus (TLCV); Cassava common mosaic virus (CCMV), (South America, Ivory coast); Cassava green mottle virus (CGMV), (Solomon islands); Cassava vein mosaic virus (CVMV), (Brazil); Cassava American latent virus (CALV), (Brazil and Guyana); Cassava ivorian bacilliform virus (CIBV), (Cote d`Ivoire); Cassava x virus (CsXV), (Columbia); Cassava c virus (an unnamed virus), (Cote d`Ivoire, Malawi and Cameroon) and Frogskin disease (FSD) for which the causal agent is unknown but a virus is suspected (Columbia). Other diseases are: Cassava antholysis (Brazil, the Caribbean, Central America) and Cassava witches` broom (Brazil and Mexico), diseases caused by phytoplasms. Among these virus diseases, CMD plays an important role and involves considerable losses which can reach 20 to 95%. Indeed, some cultivars selected for their resistance with respect to the ACMV proved very sensitive to the viral diseases once introduced in Togo. Recent studies proved that this viral disease is caused by three viruses (ACMV, EACMV and ICMV) belonging to the family of Geminiviridae and the genus of Begomovirus (Fauquet and Stanley, 2003). According to some studies, theses viruses occur in specific regions (Hong et al., 1993). For instance, EACMV is known to be present only in East Africa until Fondong et al. (1998) demonstrated recently its presence in some other part of Africa like West Africa. Up to now, ICMV is known to be present only in India. But these Begomoviruses were not intensely studied in Togo although preliminary results obtained at the University de Lomé/Togo, Laboratoire de Virologie et de Biotehnologie Végétales (LVBV) indicated the presence of ACMV and probably of EACMV in this country (Gumedzoe et al., 1999). The identification of Begomovirus other than the ACMV in Togo will give a new orientation to the research on cassava resistant or tolerant cultivars with respect to the population of Begomovirus in Togo. The prevalence of cassava viral diseases in Togo is very high; 100% according to certain authors (Dengel, 1980; Gumedzoe et al., 1999). According to Fauquet and Fargette (1990) CMD is present in all the areas where cassava is cultivated. The present study has the following objectives to evaluate the severity of the symptoms caused by cassava viral diseases in Togo and the incidence of the various components of Begomovirus which are ACMV, EACMV and ICMV in Togo using specific diagnostic tools.
MATERIALS AND METHODS
Field Survey and Sampling Procedure
During the period of July-September 2004 and 2005, fields were surveyed in the five economic regions of Togo (Central, Kara, Maritime, Plateaux and Savannas Regions). The prevalence of the viral diseases was evaluated in every region by calculating the number of fields in which at least one cassava plant presented symptoms of viral diseases divided by the total number of fields observed in this region. The severity of the symptoms was calculated for each region by using a 1 to 5 point scale of symptom expression (James, 1974). The prospected fields are, either planted with cassava only or associated with other crops (yam, leguminous plants, maize etc.) and are distant from each other of approximately 3 km in the same locality. In total, 183 fields were monitored. In each field, plants were observed along an imaginary broken line in the shape of Z. Foliar samples of cassava and wild plants presenting symptoms of mosaic, distortion and leaf curling, were collected. Six hundred and ninety seven foliar samples were collected during the field surveys. Samples were preserved with ice blocks and later on stored at -20°C in the laboratory until their use. The percentage of each of the three viruses tested was obtained by calculating the percentage of the number of samples where the particular virus was identified to the number of samples presenting symptoms analyzed.
Extraction of the Total DNA
The total viral DNA was extracted from the various foliar samples collected according to the method described by Dellaporta et al. (1983). Fifty milligram of cassava infected leaf tissue was ground in 500 μL of Dellaporta extraction buffer (100 mM Tris pH 8, 50 mM EDTA, 500 mM NaCl, 10 mM 2-mercaptoethanol) with pestle in 1.5 mL microtube and 33 μL of 20% (w/v) of SDS were added, the mix was vortexed and incubated at 65°C for 10 min. Then 160 μL of 5 M potassium acetate were added to the extract and vortexed and finally centrifuged 10 min at 14,000 g. The supernatant (580 μL) was transferred to new 1.5 mL microtube and 350 μL of ice cold isopropanol was added, vortexed and centrifuged for 10 min at 14,000 g and the supernatant was removed and discarded; the pellet was dried for 5 min in a Speed-Vac dryer and resuspended in 50 μL of RNase A (10 μg mg-1 in 10 mM Tris-HCl pH 8, 1 mM EDTA) and left at room temperature for 30 min. DNA was finally precipitated for 30 min at -80°C in 2.5 volumes of absolute ethanol in the presence of 0.3 M sodium acetate. The pellet was washed twice in 1 mL of ethanol 70%, dried for 5 min in a Speed-Vac dryer and resuspended in 500 μL distilled water.
Diagnosis by PCR
Identification of the begomovirus in the samples was done by PCR amplification using specific primers designed previously to detect ACMV, EACMV and ICMV (Fondong et al., 2000). These primers are described in Table 1. Primer pairs JSP001/JSP002 and JSP001/JSP003 were, respectively used to detect ACMV and EACMV and give fragments of 770 base pairs, primer pair JSP012/JSP013, specific of ICMV gives fragments of 702 base pairs.
The negative and positive controls (Table 2) were provided by Dr. Max Schoenfelder of DSMZ (Germany). One of the positive controls for ACMV was provided by Dr. Peterschmitt of CIRAD (France) and was isolated in Côte d`Ivoire.
The PCR reaction was performed in a volume of 25 μL containing 2 μL of extracted DNA, 1 unit of Taq Polymerase Extrapol 1 (Eurobio), 1.5 mM MgCl2, 100 μM of each dNTP and 0.8 μM of each primer in the reaction buffer provided by the manufacturer. The PCR conditions are as followed: a 94°C denaturation step of 2 min followed by 30 cycles of 30 sec at 94°C, 30 sec at annealing temperature (50°C for ACMV and EACMV and 52°C for ICMV) and 1 min at 72°C and then a final elongation step of 10 min at 72°C.
PCR Products Sequencing
To confirm the occurrence of EACMV and ICMV in Togo, PCR products of CP gene were sequenced commercially (Genome express, Meylan, France) and submitted to GenBank. The sequences obtained from both EACMV and ICMV isolates were compared to other Begomoviruses using Clustal algorithm and Darwin 4 software. Genbank accession numbers of EACMV sequences used for those comparisons were: AF259896 (from Ivory Coast/Cameroon), AJ717534 (from Kenya), AJ006461 (from Malawi), AY795985 (from Tanzania) and AF126804, AF423178 and AY562424 (from Uganda). Genbank accession number of ICMV sequence used for comparison was DQ780004.
| Table 1: | Specific primers used for the identification of ACMV, EACMV and ICMV |
 | |
| Source: Fondong et al. (2000) | |
| Table 2: | List of positive controls used |
 | |
RESULTS
Prevalence of Cassava Viral Diseases and Severity of the Symptoms in the Fields
The rate of viral diseases prevalence is 100% for the prospected regions. The percentages of the diseased cassava plants as well as the severity of the symptoms are variable according to the zones of production and arise as follows: 67.3% for the Central Region; 55.01% for the Maritime Region; 56% for the Plateaux Region; 78% for the Kara Region and 85% for the Savannas Region (Table 3).
The percentage of diseased plants with symptoms severity note of 4 to 5 are also variable as follows: 13.02% for the Central Region; 4.9% for the Maritime Region; 7.58% for the Plateaux Region; 35.14% for the Kara Region and 26.66% for the Savannas Region (Table 3).
The highness of the deviation at the level of the different rates could explain the variation in the expression of the disease and in symptom severity in the different zones of cassava production (Table 3).
Incidence Rate of the Three Begomoviruses Identified
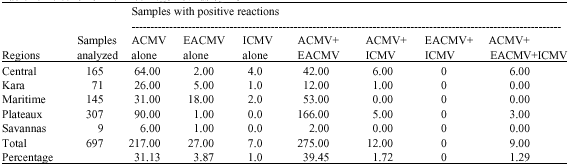
The results obtained by PCR with primers specific to ACMV, EACMV and ICMV, indicate that apart from the ACMV, two other viruses EACMV and ICMV infect cassava in Togo (Fig. 1-3). The percentage of plant infected with each virus species is: 73.60% for ACMV; 44.62% for EACMV and 4.01% for ICMV.
The three viruses which infect cassava in Togo are often in mixed infections with the following rates: 39.45% in the case of ACMV and EACMV, 1.72% in the case of ACMV and ICMV, 1.29% in the case of ACMV, EACMV and ICMV. Twenty two percent of the samples were infected by non identified viruses (in fact all the samples collected in this study present typical symptoms of virus) (Table 4).
| Table 3: | Percentages of diseased cassava plants and symptom severity rates |
 | |
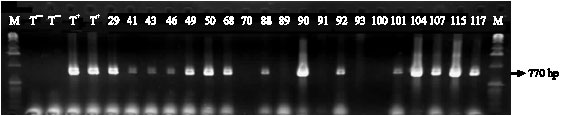 | |
| Fig. 1: | ACMV detection by PCR on DNA extracts obtained from diseased cassava plants from Togo. PCR performed using ACMV specific primers JSP001/JSP002 give the expected 770 base pairs fragment. Lane M, 1 kb DNA ladder (Roche); lanes T-, negative controls; lanes T+, ACMV positive controls (PV-0421); lanes T-e, water negative control; lanes 29, 41, 43, 46, 49, 50, 68, 70, 88, 89, 90, 91, 93, 100, 101, 104, 107, 115 and 117, diseased cassava plants form various zones of cassava production in Togo. 5 samples here are negative for the presence of ACMV |
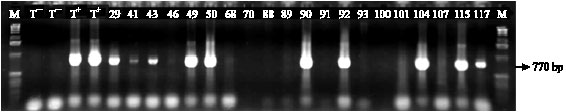 | |
| Fig. 2: | EACMV detection by PCR on DNA extracts obtained from diseased cassava plants from Togo. PCR performed using EACMV specific primers JSP001/JSP003 give the expected 770 base pairs fragment. Lane M, 1 kb DNA ladder (Roche); lanes T-, negative controls; lanes T+, EACMV positive controls (PV-0423); lanes 29, 41, 43, 46, 49, 50, 68, 70, 88, 89, 90, 91, 93, 100, 101, 104, 107, 115 and 117, cassava plants form various zones of cassava production in Togo. 10 samples here are negative for the presence of EACMV |
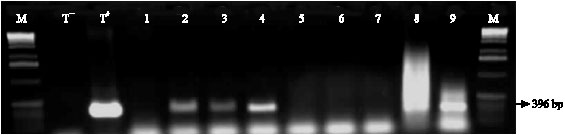 | |
| Fig. 3: | ICMV detection by PCR on DNA extracts obtained from diseased cassava plants from Togo. PCR performed using ICMV specific primers JSP012/JSP013 gives 396 base pairs fragment in this study. Lane M, 1 kb DNA ladder (Roche); lane T-, negative control; lane T+, ICMV positive control (PV-0424); lanes 1 to 9, plants from various zones of cassava production in Togo |
| Table 4: | Detection of the three Begomovirus by PCR |
 | |
Sequence Analysis and Comparison of EACMV and ICMV
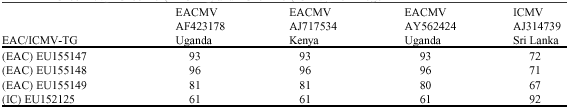
Three CP gene sequences of EACMV from were submitted to Genbank: EU155147, EU155148 and EU155149. One ICMV partial sequence (CP gene) was submitted to Genbank: EU152125. Sequence comparisons of EACMV and ICMV from Togo and EACMV and ICMV sequences from GenBank are presented in Table 5. Following Padidam et al. (1995), the criteria utilised to distinguish between different viruses were; 90-100% for isolates, 80-90% for strains and less than 80% for species demarcation. Clustal alignments revealed a percentage of nucleotide sequence identity from 93 to 96% between the Togolese EACMV sequences EU155147/EU155148 and sequences of EACMV isolates from Kenya and Uganda. Nucleotide sequence identity between EACMV sequence EU155149 from Togo and EACMV isolates from Ivory Coast/Cameroon, from Tanzania and from Malawi, is 93-96%. Nucleotide sequence identity between the Togolese ICMV sequence EU152125 and ICMV sequence DQ780004 from Sri Lanka is 92%.
| Table 5: | Nucleotide sequence identity of pairwise combinations of EACMV/ICMV from GenBank and EU155147, EU1551478, EU155149 (EACMV) and EU152125 (ICMV) from Togo |
 | |
DISCUSSION
The results from our field surveys reveal that these viral diseases are present in all the zones of cassava production in Togo. Percentages of cassava plants severely attacked (evaluation note from 4 to 5) and which could reflect the severity of the disease in the prospected fields vary from one locality to another with the rate of 4.5 to 36%. This high rate of symptom severity on the plants in the different regions can be explained by a possible co-infection by ACMV and EACMV or a recombination between the two viruses which unusually induces severe symptoms under field conditions as it was demonstrated by Fondong et al. (2000) and Pita et al. (2001) or by the susceptibility of some cultivars in the prospected fields.
The identification by PCR of the different Begomovirus with specific primers revealed for the first time that the ACMV is not the only Begomovirus responsible of cassava mosaic disease in Togo. Two other Begomovirus EACMV and ICMV are also identified in several zones of cassava production and are responsible of cassava mosaic. These three viruses were identified in the analysed samples with typical symptoms of virus, appear with the incidence rates of 73.60% (ACMV), 44.62% (EACMV) and 3.72% (ICMV). These results confirmed those obtained using the TAS-ELISA in the Plant Virology and of Biotechnology Laboratory (LVBV) of the University of Lome by Gumedzoe et al. (1999). It is important to notice that ACMV and EACMV were identified on the cultivated cassava varieties as well as on the wild cassava species and other wild plants. The results obtained also reveal that these three Begomovirus are often in mixed infections with the incidence rates showed in Table 4. The coexistence of these three Begomovirus which often infects cassava in mixed infections, can explain the severity of the symptoms of this viral diseases observed in the cassava production regions; though some cultivars like Fetonegbodji and Yovovi, which are very susceptible expressed the same symptom but were infected only by one of the viruses identified. These viruses can act in synergism as it was the case in Uganda (Pita et al., 2001) and these mixed infections could also provoke recombination within the populations of Begomovirus which infect cassava in this country and could explain the severity of the symptoms in the different regions. This recombinant strains between the EACMV and the ACMV which was detected in Uganda (Harrison et al., 1997; Pita et al., 2001) may also emerge in Togo.
In this study, 22% of the analysed samples did not react to the target Begomoviruses. In regard to symptoms on these samples which are virus-like symptoms it could be said that some other viruses not yet identified are infecting cassava in Togo. A further study on the other cassava viruses in Togo will enlighten this consideration.
These results make it possible to have a better understanding of the new development of cassava mosaic disease in Togo. Sequence analysis of sequences obtained from, EACMV and ICMV identified in Togo and those already identified in other countries in Africa revealed at least two groups of EACMV in Togo: one EACMV group containing one isolate from Kenya and three isolates from Uganda and one EACMV group including isolates from Ivory Coast/Cameroon, Malawi and Tanzania. It is possible that emergence of new Begomoviruses by recombination also occur in Togo as suggested by Padidam et al. (1999).
Though, only a partial sequence was obtained from the ICMV isolate identified in Togo, the nucleotide sequence identity between this sequence EU152125 and an ICMV sequence of GenBank from Sri Lanka is 92%. This result shows clearly that an isolate of ICMV exists in Togo. It is now important to continue this research because possible recombination could occur among these Begomoviruses identified in Togo.
ACKNOWLEDGMENTS
We are grateful to the ‘Agence Universitaire de la Francophonie (AUF)` and the ‘Service de Coopération et d`Action Culturelle SCAC` for sponsoring this project.
REFERENCES
- Al-Mallah, M.K., M.R. Davey and E.C. Cocking, 1990. Nodulation of oil seed rape (Brassica napus). Rhizobia. J. Exp. Bot., 41: 1567-1572.
CrossRefDirect Link - Amooaghaie, R., A. Mostajeran and G. Emtiazi, 2002. The effect of compatible and incompatible Azospirillum brasilense strains on proton efflux of intact wheat roots. Plant Soil, 243: 155-160.
Direct Link - Antoun, H., C.J. Beachamp, N. Goussard, R. Chabot and R. Lalande, 1998. Potential of Rhizobium and Bradyrhizobium species as plant growth promoting rhizobacteria on non-legumes: Effects on radishes (Rhaphanus sativus L.). Plant Soil, 204: 56-67.
Direct Link - Baldani, I.J. and L.V. Baldani, 2005. History on the biological nitrogen fixation research in graminaceous plants: Special emphasis on the Brazilian experience. Anais Da Acad. Brasiliera De Ciencias Sci., 77: 549-579.
PubMedDirect Link - Bashan, Y. and H. Levanony, 1985. An improved selection technique and medium for the isolation and enumeration of Azospirillum brasilense. Can. J. Microbiol., 31: 947-952.
CrossRefDirect Link - Bashan, Y. and H. Levanony, 1989. Effect of root environmental on proton efflux in wheat roots. Plant Soil, 119: 191-197.
Direct Link - Bashan, Y. and G. Holguin, 1997. Azospirillum-plant relationships: Environmental and physiological advances (1990-1996). Can. J. Microbiol., 43: 103-121.
Direct Link - Bashan, Y., G. Holguin and L.E. de-Bashan, 2004. Azospirillum-plant relationship: Physiological, molecular, agricultural and environmental advances (1997-2003). Can. J. Microbiol., 50: 521-577.
PubMedDirect Link - Bashan, Y. and L.E. de-Bashan, 2005. Plant Growth-Promoting Soil. In: Encyclopedia of Soils in the Environment, Hillel, D. (Ed.). Vol. 1, Elsevier, Oxford, UK., pp: 103-115.
Direct Link - Biro, B., K. Koves-Pechy, I. Voros, T. Takacs, P. Eggenberger and R.J. Strasser, 2000. Interrelations between Azospirillum and Rhizobium nitrogen-fixers and arbuscular mycorrhizal fungi in the rhizosphere of alfalfa in sterile, AMF-free or normal soil conditions. Applied Soil Ecol., 15: 159-168.
CrossRef - Christiansen-Weniger, C., 1997. Ammonium-excreting Azospirillum brasilense C3: gusA inhabiting induced tumors along stem and roots of rice. Soil Biol. Biochem., 29: 943-950.
CrossRefDirect Link - Chirstiansen-Weniger, C., 1998. Endophytic establishment of diazotrophic bacteria in auxin-induced tumors of cereal crops. Crit. Rev. Plant Sci., 17: 55-76.
Direct Link - Cocking, E.C., C.A. Batchelor, S.L. Kothari, S. Jain, G. Webster, J. Jones, J. Jotham and M.R. Davey, 1994. Cereal symbiotic nitrogen fixation-the interaction of Rhizobia with cereals for symbiotic nitrogen fixation. CIHEAM Options Mediterraneennes, 8: 67-72.
Direct Link - Cocking, E.C., P.J. Stone and M.R. Davey, 2005. Symbiosome-like intracellular colonization of cereals and other crop plants by nitrogen-fixing bacteria for reduced inputs of synthetic nitrogen fertilizers. Sci. China Ser. C, 48: 888-896.
Direct Link - Cocking, E.C., 2005. Intracellular colonization of cereals and other crop plants by nitrogen-fixing bacteria for reduced inputs of synthetic nitrogen fertilizers in vitro. Cell Dev. Biol. Plant, 41: 369-373.
CrossRefDirect Link - Dalla Santa, O.R., R.F. Hernandez, G.L.M. Alvarez, P. Ronzelli Junior and C.R. Soccol, 2004. Azospirillum sp. inoculation in wheat, barley and oats seeds greenhouse experiments. Brazil. Arch. Biol. Tech., 47: 843-850.
Direct Link - Denison, R.F. and E.T. Kiers, 2004. Why are most rhizobia beneficial to their plant hosts, rather than parasitic? Microbes Infect., 6: 1235-1239.
Direct Link - Dobbelaere, S., A. Croonenboghs, A. Thys, D. Ptacek and J. Vanderleyden et al., 2001. Responses of agronomically important crops to inoculation with Azospirillum. Aust. J. Plant Physiol., 28: 871-879.
CrossRefDirect Link - Elanchezhian, R. and J.D.S. Panwar, 1997. Effects of 2,4-D and Azospirillum brasilense on nitrogen fixation, photosynthesis and grain yield in wheat. J. Agron. Crop Sci., 178: 129-133.
CrossRefDirect Link - Er, F., M. Kilic, A.R. Brohi and Ogut, 2004. Nodulation and growth of bean plants inoculated with genetically modified Rhizobium etli strains and Azospirillum brasilense. Agrochimica, 48: 124-133.
Direct Link - Fages, J. and J.F. Arsac, 1991. Sunflower inoculation with Azospirillum and other plant growth promoting rhizobacteria. Plant Soil, 137: 87-90.
CrossRefDirect Link - Fahraeus, G., 1957. The infection of clover root hairs by nodule bacteria studied by a simple glass slide technique. Microbiology, 16: 374-381.
CrossRefPubMedDirect Link - Gough, C., J. Vasse, C. Galera, G. Webster, E. Cocking and J. Denarie, 1997. Interaction between bacterial diazotrophs and non-legume dicots: Arabidopsis thaliana as a model plant. Plant Soil, 194: 123-130.
CrossRefDirect Link - Hardy, R.W.F., R.D. Holsten, E.K. Jackson and R.C. Burns, 1968. The acetylene-ethylene assay for N2 fixation: Laboratory and field evaluation. Plant Physiol., 43: 1185-1207.
CrossRefPubMedDirect Link - Hardy, R.W.F., R.C. Burns and R.D. Holsten, 1973. Applications of the acetylene-ethylene assay for measurement of nitrogen fixation. Soil Biol. Biochem., 5: 47-81.
CrossRefDirect Link - Hirsch, A.M., M.R. Lum and D.J. Allan Downie, 2001. What makes the Rhizobia-legume symbiosis so special? Plant Physiol., 127: 1484-1492.
CrossRefPubMedDirect Link - Hoflich, G., W. Wiehe and G. Kuhn, 1994. Plant growth stimulation by inoculation with symbiotic and associative rhizosphere microorganisms. Experientia, 50: 897-905.
CrossRefDirect Link - Holguin, G. and Y. Bashan, 1996. Nitrogen-fixation by Azospirillum brasilense Cd is promoted when co-cultured with a mangrove rhizosphere bacterium (Staphylococcus sp.). Soil Biol. Biochem., 28: 1651-1660.
CrossRef - Humphry, R.D., M. Andrews, R.S. Santos, K.E. James, V.L. Vinogradova, L. Perin, M.V. Reis and P.S. Cummings, 2007. Phylogenetic assignment and mechanism of action of a crop growth promoting Rhizobium radiobacter strain used as a biofertiliser on graminaceous crops in Russia. Antonie van Leeuwenhoek, 91: 105-113.
Direct Link - Mateos, P.F., J.I. Jimenez-Zurdo, L. Chen, A.S. Squartini and S.K. Haack et al., 1992. Cell-associated pectinolytic and cellulolytic enzymes in Rhizobium leguminosarum biovar trifolii. Applied Environ. Microbiol., 58: 1816-1822.
PubMedDirect Link - Matiru, V.N. and F.D. Dakora, 2004. Potential use of rhizobial bacteria as promoters of plant growth for increased yield in landraces of African cereal crops. Afr. J. Biotechnol., 3: 1-7.
Direct Link - McInroy, J.A. and J.W. Kloepper, 1995. Survey of indigenous bacterial endophytes from cotton and sweet corn. Plant Soil, 173: 337-342.
CrossRefDirect Link - Molla, A.H., M.S. Shamsuddin, M. Morziah and A.B. Puteh, 2001. Potential for enhancement of root growth and noduation of soybean co-inoculated with Azospirillum and Bradyrhizobium in laboratory systems. Soil Biol. Biochem., 33: 457-463.
CrossRef - Mostajeran, A., R. Amooaghaie and G. Emtiazi, 2007. The participation of the cell wall hydrolytic enzymes in the initial colonization of Azospirillum brasilense on wheat roots. Plant Soil, 291: 239-248.
CrossRefDirect Link - Ogut, M., C. Akdag, O. Duzdemir and M.A. Sakin, 2005. Single and double inoculation with Azospirillum/Trichoderma: The effects on dry bean and wheat. Biol. Fert. Soils, 41: 262-272.
CrossRefDirect Link - Reinhold-Hurek, B., T. Hureck, M. Claeyssens and M. Van-Montasu, 1993. Cloning expression in Escherchia coli and characterization of cellulolytic enzyrnes of Azoarcus sp. J. Bacteriol., 175: 7056-7065.
PubMed - Rodelas, B., J. Gonzalez-Lopez, M.V. Martinez-Toledo, C. Pozo and V. Salmeron, 1999. Influence of Rhizobium/Azotobacter and Rhizobium/Azospirillum combined inoculation on mineral composition of faba bean (Vicia faba L.). Biol. Fert. Soils, 29: 165-169.
CrossRefDirect Link - Rueda-Puente, E.O., T. Castellanos, E. Troyo-Dieguez and J.L. Diaz-d-Leon-Alvarez, 2004. Effect of Klebsiella pneumoniae and Azospirillum halopraeferens on the growth and development of two Salicornia bigelovii genotypes. Aust. J. Exp. Agric., 44: 65-74.
Direct Link - Sadovnikova, Y.N., L.A. Bespalova and L.P. Antonyuk, 2003. Wheat germ agglutinin is a growth factor for bacterium Azospirillum brasilense. Doklady Biochem. Biophys., 389: 103-105.
CrossRefDirect Link - Saubidet, M.I., N. Fatta and A.J. Barneix, 2002. The effect of inoculation with Azospirillum brasilense on growth and nitrogen utilization by wheat plants. Plant Soil, 245: 215-222.
CrossRefDirect Link - Spencer, D., E.K. James, G.J. Ellis, J.E. Shaw and J.I. Sprent, 1994. Interaction between Rhizobia and potato tissues. J. Exp. Bot., 45: 1475-1482.
CrossRefDirect Link - Thuler, D.S., E.I.S. Floh, W. Handro and H.R. Barbosa, 2003. Plant growth regulators and amino acids released by Azospirillum sp. in chemically defined media. Lett. Applied Microbiol., 37: 174-178.
CrossRefPubMedDirect Link - Tripathi, A.K. and B.M. Mishra, 1996. Isolation and characterization of Azospirillum lipoferum locus that complements Rhizobium meliloti dctA and dctB mutations. Can. J. Microbiol., 42: 503-506.
PubMed - Tsavkelova, E.A., S.Y. Klimova, T.A. Cherdyntseva and A.I. Netrusov, 2006. Microbial producers of plant growth stimulators and their practical use: A review. Applied Biochem. Microbiol., 42: 117-126.
CrossRef - Van Rhijin, P. and J. Vanderleyden, 1995. The Rhizobium-plant symbiosis. Microbiol. Rev., 59: 124-142.
PubMedDirect Link - Webster, G., V. Jain, M.R. Davey, C. Gough, J. Vasse, J. Denarie and E.C. Cocking, 1998. The flavonoid naringenin stimulates the intercellular colonization of wheat roots by Azorhizobium caulinodans. Plant Cell Environ., 21: 373-383.
CrossRefDirect Link - Yanni, Y.G., R.Y. Rizk, V. Corich, A. Squartini and K. Ninke et al., 1997. Natural endophytic association between Rhizobium leguminosarum bv. trifolii and rice roots and assessment of its potential to promote rice growth. Plant Soil, 194: 99-114.
CrossRefDirect Link - Zaady, E., A. Perevolotsky and Y. Okon, 1993. Promotion of plant growth by inoculums with aggregated and single cell suspensions of Azospirillum brasilense Cd. Soil Biol. Biochem., 25: 819-823.
CrossRef - Zeman, A.M.M., Y.T. Tchan, C. Elmerich and I.R. Kennedy, 1992. Nitrogenase activity in wheat seedlings bearing para-nodules induced by 2,4-dichlorophenoxyacetic acid (2,4-D) and inoculated with Azospirillum. Res. Microbiol., 143: 847-855.
Direct Link - Zimmer, W. and H. Bothe, 1988. The phytohormonal interactions between Azospirillum and wheat. Plant Soil, 110: 239-247.
CrossRef