
P. Kashiani
Department of Crop Science, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
G. Saleh
Department of Crop Science, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
N.A.P. Abdullah
Department of Crop Science, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
S.N. Abdullah
Department of Crop Science, Faculty of Agriculture, Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
Asian Journal of Crop Science
Year: 2010 | Volume: 2 | Issue: 2 | Page No.: 78-84







ABSTRACT
Nine advanced tropical sweet corn (Zea mays L. saccharata) inbred lines were evaluated to determine their performance, to estimate broad-sense heritability of the traits measured and to determine phenotypic correlations among the traits. The inbred lines showed a wide range of performance for husked fresh ear yield and its components. Based on performance and earliness, inbred lines Bakti-1-S7, TSS Melaka-S5, Thailand-S6, MM x Indonesia-S4 and Manis Madu-S7 were found to be superior and could be short-listed for further testing for general and specific combining ability analysis towards development of hybrid varieties. Broad-sense heritability (h2B) estimates obtained from the variance components method were found to be high for number of days to silking (80.5%), plant height (79.9%), number of days to tasseling (66.9%) and ear height (63.7%), moderate for husked ear yield (56.7%), total soluble solid concentration (54.2%), number of kernel rows per ear (53.9%), ear diameter (46.7%), dehusked ear yield (43.1%), number of kernels per row (42.6%) and number of ear per hectare (34.5%), while lowest estimates for dehusked ear length (12.9%) and husked ear length (0.3%). Traits found to be highly correlated with husked ear yield were dehusked ear yield (0.97), number of kernel rows per ear (0.71), plant height (0.69), husked ear length (0.67), ear height (0.66), dehusked ear length (0.63), ear diameter (0.55), number of kernels per row (0.50) and number of ears per hectare (0.49). For selection purposes, it is therefore suggested that emphasis should be given on traits like number of kernel rows per ear, plant and ear height and ear length for yield improvement among the inbred lines.
PDF Abstract XML References Citation
How to cite this article
P. Kashiani, G. Saleh, N.A.P. Abdullah and S.N. Abdullah, 2010. Variation and Genetic Studies on Selected Sweet Corn Inbred Lines. Asian Journal of Crop Science, 2: 78-84.
DOI: 10.3923/ajcs.2010.78.84
URL: https://scialert.net/abstract/?doi=ajcs.2010.78.84
DOI: 10.3923/ajcs.2010.78.84
URL: https://scialert.net/abstract/?doi=ajcs.2010.78.84
INTRODUCTION
In Malaysia there is an increasing trend in sweet corn production mainly due to its high economic return (Saleh et al., 2001). However, the local varieties, which are currently planted throughout the country, are late in maturity, tall and less sweet. One of the most important approaches to improve the performance of the populations is by development of inbred lines which can produce high-yielding hybrid varieties. Inbred line development is the main prerequisite for production of hybrid varieties. This process is achieved through successive generations of inbreeding followed by repeated testing and selection. Inbreeding is the process of mating between genetically related individuals. The fundamental genetic change that inbreeding produces a loss of heterozygosity, was well understood early in the development of the field of genetics (Hill et al., 2006; Marquez-Sanchez, 1998; Wright, 1921).
Selfing, the most extreme form of inbreeding, reduces heterozygosity by 50% each generation. The decrease in heterozygosity typically results in phenotypic changes that we call inbreeding depression, the genetic basis of which has been debated for almost a century. As a consequence of selfing, recessive genes, which were earlier masked in the heterozygous forms, become homozygous. These genes, if conferring to undesirable phenotypes, will result in the deterioration of the succeeding generations. In cross-pollinated crops which do not have self-incompatibility problems, like sweet corn, inbred lines for hybrid varieties are developed through selfing. Extensive studies on inbreeding depression in sweet corn (Zea mays L. saccharata) have indicated that selfing is important in inbred line development because it leads to rapid gene homozygosity, whereby desirable dominant genes can be accumulated while the undesirable ones are eliminated (Gallais, 2009; Saleh et al., 1993). Selected plants are usually self-pollinated for several generations until homozygosity is reached. Hallauer and Miranda (1988) reported that many undesirable recessive genes are eliminated from families as a result of the inbreeding process, then selection is applied within and between lines for the best individual plants (Acquaah, 2007). Hallauer (1990) added that corn inbred lines developed from improved source populations would have greater vigour and grain yield as compared to those developed from unimproved sources. Stoskopf et al. (1993) also cited that inbred lines are developed after five to seven generations of selfing, during which selection for characters of interest is also conducted. Heritability is a measure of the degree to which the variance in distribution of a phenotype is due to genetic causes (Ullrich, 2007; Sleper and Poehlman, 2006). Ali et al. (2003) found that broad-sense heritability estimated from their sweet corn population were moderate to high for the traits measured. The highest estimate was for ear height (99.8%), while the lowest was revealed by ear diameter (61.9%). Saleh et al. (2002a) reported that broad-sense heritability estimates for grain yield in maize were generally moderate. Correlation is the co-relationship between two variables and a correlation coefficient is the measure of the degree of association between two variables (Gepts, 2002; Wallace et al., 1993; Mayo, 1987). Correlations may occur in positive or negative values from zero to ±1.0. The closer the coefficient is to either -1 or 1, the stronger is the correlation between variables (Miles and Shevlin, 2001). Hence, correlation is an important parameter to be estimated as it helps to determine relationship among traits before selection is conducted (Kashiani et al., 2008). According to Saleh et al. (2002b), number of ears per hectare, plant height, ear weight, ear length and number of kernel rows per ear were found to be most correlated with ear yield. However, days to tasselling, days to silking, ear diameter and total soluble solids content were not correlated with ear yield. The objectives of this study were to determine the performance of nine inbred lines as potential hybrid parents, to estimate broad-sense heritability of the traits measured on the nine inbred lines and to determine phenotypic correlations among those traits.
MATERIALS AND METHODS
Location of Study
The study was conducted from March to May 2009, at filed 10, Faculty of Agriculture, Universiti Putra Malaysia, Serdang, Selangor, located at 3° 40' North; 101° 42' East and 31 m above sea level.
Plant Materials
This study was an advanced stage of a long-term tropical sweet corn inbred line development programme conducted at Universiti Putra Malaysia where a series of near-homozygous inbred lines were formed from various tropical origin source populations.
The inbred lines were namely Bakti-1-S7, Manis Madu-S7, TSS Tin-S7, Mas Madu-S6, Thailand-S6, Indonesia-S6, TSS Melaka-S5, Manis Madu x Indonesia-S4 and SBY-S2.
Experimental Design and Plot Arrangement
The evaluation was carried out in a Randomized Complete Block Design (RCBD) with three replications. Each replicate consisted of nine plots, each represented by an inbred line. The planting density used was 0.75x0.25 m. Each plot consisted of seven 12 m long plant rows, where only the three middle rows measuring 4 m in length were used as the harvest area. All experimental plots were subjected to uniform agronomic practices.
Cultivation Practices
Before planting, the soil was ploughed twice to a depth of 15 to 30 cm, followed by rotorvation. The soil type in the evaluation plot was sandy loam (56% sand, 23% silt and 21% clay). The soil pH was raised up to 6.5 by applying ground magnesium limestone one month before planting. Planting was done manually, at the rate of two seeds per point and later thinned to just one plant per point at 10 days after planting. Fertilizer at the rate of 160 kg ha-1 N, 100 kg ha-1 P2O5 and 100 kg ha-1 K2O was given in split applications, where the balanced compound fertilizer (15:15:15) was used at seven days after planting, followed by Urea (46% N) at 20 and 35 days after planting. Weeds were controlled manually using a pre-emergence herbicide, Lasso (2-chloro-2'-6'-diethyl-N-methoxymethyl) and a post-emergence contact herbicide, Gramoxone (1,1, dimethyl-dimethyl-4, 4'-bipyridinium). A sprinkler irrigation system was used to supply water to the plants when necessary.
Data Collection
Data were collected from both pre- and post-harvest characters including days to tasselling, days to silking, plant height (cm), ear height (cm), number of ears per hectare, fresh ear yield (kg ha-1), ear length (cm), ear diameter (mm), number of kernels per row, number of kernel rows per ear and Total Soluble Solids (TSS) concentration (%). Fresh ears were harvested from each plot separately, at 21 days after plant tasselling. Three middle rows of each plot measuring four meters long, giving an area of 9 m2, were used as the harvest area for yield and number of ears per hectare estimations. Among the plants in the harvest area, 12 plants were used as samples for the measurements of individual plant and ear data.
Data Analysis
Data collected were analyzed using SAS computer package. The Analysis of Variance (ANOVA) was used to determine the significance of variation among inbred lines. Subsequently, the Duncan New Multiple Range Test (DNMRT) was applied for comparison of mean performance of the inbred lines.
Broad-sense heritability (h2B) for the traits was estimated using the variance components method suggested by Becker (1984), as follows:
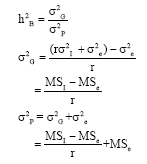 |
Where:
| h2B | : | Broad-sense heritability |
| σ2G | : | Genotypic variance |
| σ2P | : | Phenotypic variance |
| σ2I | : | Variance for inbred lines |
| σ2e | : | Environmental variance |
| MSI | : | Mean squares due to inbred line |
| MSe | : | Error mean squares |
| r | : | No. of replications |
Simple correlations among the characters measured were also analyzed using the formula based on Gomez and Gomez (1984), as follows:
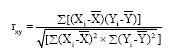 |
Where:
| rxy | : | Correlation between the traits X and Y |
| Xi | : | X value |
| Yi | : | Y value |
| : | Mean value of character X | |
| : | Mean value of character Y |
RESULTS AND DISCUSSION
Results of the analysis of variance showed significant effects among the inbred lines evaluated for all characters studied except husked and dehusked ear lengths. Mean values for the performance of the inbred lines are shown in Table 1. This indicates that the inbred lines evaluated varied substantially in many aspects and these differences could be exploited for specific purpose in breeding programmes. From separation of mean values indicated by DNMRT (Table 1), highest fresh husked ear yields were revealed by inbred lines Bakti-1-S7, TSS Melaka-S5, Thailand-S6 and Manis Madu-S7, with yields of 8043, 7532, 6053 and 5298 kg ha-1, respectively. The superiority of inbred lines Bakti-1-S7, TSS Melaka-S5, Thailand-S6 and Manis Madu-S7 was also revealed by the high number of productive ears per hectare they produced, amounting to 35556, 41481, 44444 and 35556, respectively. There is therefore, a strong indication of the potential of these inbred lines for their utilisation towards production of new hybrid varieties. Bakti-1-S7 was found to be significantly taller than other inbred lines with (175.1 cm) and also tasseled and silked significantly sooner than the others. Indonesia-S6 was the only inbred line which tasseled about the same time as Bakti-1-S7. The lowest Total Soluble Solids (TSS) concentration was obtained from MM x Ind-S4, while no significant difference among other inbred lines were observed. Kashiani et al. (2008) reported that the inbred lines extracted from the source populations including Bakti-1, TSS Melaka, Thailand and Manis Madu performed better than the rest in term of fresh ear yield, number of productive ears per hectare and days to silking and tasseling.
Genotypic and phenotypic variances and broad-sense heritability (h2B) estimates for yield and yield components measured among the nine inbred lines evaluated are shown in Table 2. Days to silking was found to be the most heritable trait in the inbred lines, with heritability of 80.5%, followed by plant height (79.9%), number of days to tasselling (66.9%) and ear height (63.7%).
| Table 1: | Mean values for traits measured on nine sweet corn inbred lines evaluated |
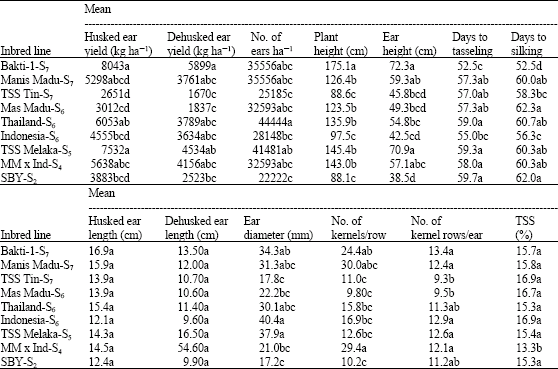 | |
| Means followed by the same letter(s) in the same column are not significantly different at p≤0.05 based on DNMRT | |
| Table 2: | Genotypic variances (σ2G), phenotypic variances (σ2P) and broad-sense heritability estimates (h2B) for traits measured on nine sweet corn inbred lines |
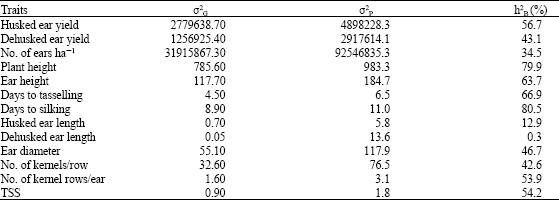 | |
This indicates that selection for these traits in these inbred lines would be most effective for the expression of these traits in the succeeding generations. Similar results on broad-sense heritability were reported by Kashiani et al. (2008) and Saleh (2003) showing high estimates for days to silking, plant height and ear height. Saleh et al. (2002c) also reported high broad-sense heritability estimates for plant height and number of days to tasselling on selected sweet corn synthetic populations. Moderate broad-sense heritability estimates were obtained from husked ear yield (56.7%), total soluble solids concentration (54.2%), number of kernel rows per ear (53.9%), ear diameter (46.7%), dehusked ear yield (43.1%), number of kernels per row (42.6%) and number of ear per hectare (34.5%), while lowest estimates were revealed by husked ear length (12.9%) and dehusked ear length (0.3%). Fresh ear yield, ear diameter and number of kernels per row were reported to possess moderate heritability estimates (54.7, 37.0 and 36.0%, respectively) in a study on sweet corn breeding population conducted by Nigussie and Saleh (2007).
| Table 3: | Simple correlation coefficients among traits measured on nine sweet corn inbred lines |
 | |
| **, *Significant at p = 0.01, p = 0.05, respectively, ns: Non-significant, TSS: Total soluble solids 1: Husked ear yield, 2: Dehusked ear yield, 3: No. of ears ha-1, 4: Plant height, 5: Ear height, 6: Days to tasselling, 7: Days to silking, 8: Husked ear length, 9: Dehusked ear length, 10: Ear diameter, 11: No. of kernels/row, 12: No. of kernel rows/ear | |
Results on simple correlation among characters are presented in Table 3. Husked ear yield was found to be highly positively correlated (p≤0.01) with dehusked ear yield, plant height, ear height, husked ear length, dehusked ear length, ear diameter, number of kernels per row and number of kernel rows per ear (with correlation coefficients of 0.96, 0.69, 0.66, 0.67, 0.63, 0.55, 0.50 and 0.71, respectively), while positively correlated (p≤0.05) with number of ears per hectare (r = 0.48). This indicates that high measurements of these traits had direct positive contribution to husked ear yield. Therefore, for selection purposes to improve the inbred lines, it is suggested that emphasis should be given on these traits. Plant height was highly significantly correlated with husked and dehusked ear yields, indicating that taller plants were better yielding compared to the shorter ones. This might be attributed to the high dry matter accumulation function carried out by the high number of leaves possessed by tall plants. A similar finding has been reported by Kashiani et al. (2008), Sujiprihati et al. (2003) and Saleh et al. (2002b).
In conclusion, significant differences in performance indicated that lines varied among the inbred lines for the measured traits substantially in many aspects and these differences could be exploited for specific purposes in breeding programmes. This also gave an indication of positional of the inbred lines to be used as promising parents for potential hybrids in Malaysia. Genotyping of inbred lines can be performed as an extra tool to select superior inbred lines for further diallel analysis to reveal general and specific combing abilities.
ACKNOWLEDGMENT
The authors gratefully acknowledge the Ministry of Science, Technology and Innovation (MOSTI) for the financial support on the research under e-Science Fund (Project No. 05-01-04-SF0700).
REFERENCES
- Ali, E.S., G.B. Saleh, Z.B. Wahab and A.A. Rahim, 2003. Performance, heritability and correlation studies on varieties and population cross of sweet corn. Asian J. Plant Sci., 2: 756-760.
CrossRefDirect Link - Gallais, A., 2009. Full-Sib reciprocal recurrent selection with the use of doubled haploids. Crop Sci., 49: 150-152.
Direct Link - Gepts, P., 2002. A comparison between crop domestication, classical plant breeding and genetic engineering. Crop Sci., 42: 1780-1790.
Direct Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wiley and Sons, New York, USA., ISBN-13: 9780471870920, Pages: 680.
Direct Link - Hallauer, A.R. and J.B. Miranda, 1988. Quantitative Genetics in Maize Breeding. 2nd Edn., Iowa State University Press, Ames, Iowa, ISBN: 9780813815220, Pages: 468.
Direct Link - Hill, W.G., N.H. Barton and M. Turelli, 2006. Prediction of effects of genetic drift on variance components under a general model of Epistasis. Theoretical Populat. Biol., 70: 56-62.
Direct Link - Marquez-Sanchez, F., 1998. Expected inbreeding with recurrent selection in maize: I. Mass selection and modified ear-to-row selection. Crop Sci., 38: 1432-1436.
Direct Link - Mayo, O., 1987. The Theory of Plant Breeding. 2nd Edn., Oxford University Press, New York, USA., ISBN: 0198541716,.
Direct Link - Saleh, G., D. Abdullah and A.R. Anuar, 2002. Effects of location on performance of selected tropical maize hybrids development in Malaysia. Pertanika J. Trop. Agric. Sci., 25: 75-86.
Direct Link - Saleh, G.B., D. Abdullah and A.R. Anuar, 2002. Performance, heterosis and heritability in selected tropical maize single, double and three-way cross hybrids. J. Agric. Sci., 138: 21-28.
CrossRefDirect Link - Saleh, G.B., S.A.S. Alawi and K. Panjaitan, 2002. Performance, correlation and heritability studies on selected sweet corn synthetic populations. Pak. J. Biol. Sci., 5: 251-254.
CrossRefDirect Link - Saleh, G., M.R. Yusop and T.C. Yap, 1993. Inbreeding depression and heterosis in sweet corn varieties Manis Madu and Bakti-1. Pertanika J. Trop. Agric. Sci., 16: 209-214.
Direct Link - Sujiprihati, S., G.B. Saleh and E.S. Ali, 2003. Heritability, performance and correlation studies on single cross hybrid of tropical maize. Asian J. Plant Sci., 2: 51-57.
CrossRefDirect Link - Wallace, D.H., J.P. Baudoin, J. Beaver, D.P. Coyne and D.E. Halseth et al., 1993. Improving efficiency of breeding for higher crop yield. Theor. Applied Genet., 86: 27-40.
Direct Link - Wright, S., 1921. Systems of mating. II. The effects of inbreeding on the genetic composition of a population. Genetics, 6: 124-143.
Direct Link