
Manal Hefny
Department of Agronomy, Faculty of Agriculture, Suez Canal University, Ismailia, 41522, Egypt
Asian Journal of Crop Science
Year: 2010 | Volume: 2 | Issue: 4 | Page No.: 236-249







ABSTRACT
The present research was designed to study the combining ability effects of 15 parents of maize (13 inbred lines and 2 testers) crossed in line x tester scheme and assessed for yield and its components. The genetic materials were evaluated using randomized complete block design with three replications in two sowing dates, June, 1 and July, 1 at the experimental farm of Suez Canal University, Ismailia, Egypt in 2009 cropping season. Parents and crosses showed significant differences for all measured traits at both and combined over dates. On the other hand, all measured traits were affected significantly by sowing date (except rows ear-1), parents x SD (except days to 50% silking) and crosses x SD (except ear diameter). Delaying in sowing caused reduction in means of all evaluated traits (except ear diameter and rows ear-1). Line x tester analyses confirmed the preponderance of δ2SCA in controlling the inheritance of the majority of traits relative to δ2GCA. Moreover, the contribution of lines was pronounced in the expression of all traits at both late and combined dates. L6, L7, L8, L10 and L12 were found to be good combiners where they recorded desirable GCA effects for yield plant-1, some of yield components and flowering traits in one or both sowing dates. L10 was distinguished as one which recorded desirable and favorable GCA for earliness, yield plant-1 and its components. In addition to their superiority as combiners, L8 and L12 recorded desirable SCA effects for earliness, ear characteristics and yield plant-1. The previous parents can be crossed with other promising lines to develop early maturing and high yielding hybrids/varieties for cultivating under late sowing.
PDF Abstract XML References Citation
How to cite this article
Manal Hefny, 2010. Genetic Control of Flowering Traits, Yield and its Components in Maize (Zea mays L.) at Different Sowing Dates. Asian Journal of Crop Science, 2: 236-249.
DOI: 10.3923/ajcs.2010.236.249
URL: https://scialert.net/abstract/?doi=ajcs.2010.236.249
DOI: 10.3923/ajcs.2010.236.249
URL: https://scialert.net/abstract/?doi=ajcs.2010.236.249
INTRODUCTION
Maize (Zea mays L.) is the third most important crop among cereal crops grown worldwide. It is cultivated and produced for human, animal and industrial purposes. Temperature, irrigation and soil fertility are three major limiting factors affecting maize growth and productivity. Experts agree that temperatures will increase and that there will be more year-to-year variability due to climate change (Maton et al., 2007), which depends largely on the geographical location (IPCC, 2001). Regarding rainfall, predictions tend to suggest drier summers and wetter springs and autumns. This variation in the rainfall regime will affect different technical operations, particularly irrigation and sowing date, as weather conditions for sowing date will be modified (Doll, 2002).
Sowing date is a particularly important technical operation as it determines the timing of the crop cycles, as well as the earliness of the crop, thus determines the length of the growing period (Girardin, 1999). Sowing dates depend not only on overall farmer’s objectives and constraints (machinery availability, area to be sown, work rate of the machinery, etc.) but also on the possibilities the farmer has to sow his crops, suitable soil conditions and the farmers time available for sowing.
Environmental changes associated with different sowing dates (sunshine and temperature) have a modifying effect on the growth and development of maize plants. Moreover, each maize hybrid has an optimum sowing date and the greater the deviation from this optimum (early or late sowing), the greater the yield loss. The optimal day temperature ranges from 25-32°C and night temperature ranges from 16.7 - 23.3°C for maize plant. At this temperature, photosynthetic rate is rapid than respiration which results in plant growth enhancement. Maize growth is affected adversely when temperature decreases to 5°C or increases beyond 32°C (Akbar et al., 2008). Raising temperature (+30°C) increases anthesis-silking interval and resulting in poor synchronization of flowering. Further increase in temperature reduces the pollen viability and silk receptivity resulting in poor seed set and yield reduction (Samuel et al., 1986).
Normally in Egypt, maize grows and yields at optimum date from the beginning of May to June, 1. Warmer temperature has generally a favorable effect on faster crop development. Campos et al. (2004) expected significant yield losses in maize caused by drought/heat stress due to changing global climate. The environmental and agronomic responses of maize hybrids determine their adaptability and influence improvements in maize production through agronomy and breeding. So, it is essential to have information on the nature of combining ability of parents, their behavior and performance in hybrid combinations Such knowledge is essential for selection of suitable parents for hybridization and identification of promising hybrids/varieties for a diverse agro-ecological zones (Bello and Olaoye, 2009). The line x tester analysis method is used to breed both self and cross-pollinated plants and to estimates favorable parents and crosses and their general and specific combining abilities (Kempthorne, 1957). Combining ability analysis is an important tool for the selection of desirable parents together with the information regarding nature and magnitude of gene effects controlling quantitative traits (Basbag et al., 2007).
Results on the inheritance of maize yield and agronomical traits are presented by many researchers. Grain plant-1 and 100-grain weight were found to be under non-additive gene control (Dubey et al., 2001; Sinsawat et al., 2004). Whereas, additive gene action was responsible for the genetic expression for days to 50% tesseling and rows ear-1.
The objective of the present study were to investigate combining ability for some agronomic traits in introduced and local maize inbred lines using line x tester analysis to identify parents with desirable GCA effects and cross combinations with desirable SCA effects and to study the nature of gene action involved in yield and related traits under two sowing dates.
MATERIALS AND METHODS
Genetic Materials and Experimental Design
Thirteen maize inbred lines and two white-grain single cross testers, SC.122 (T1) and Pioneer 30B51 (T2) were used for this study. The inbred lines including, four introduced inbreds with yellow endosperm from National Plant Germplasm System, USA. Whereas, the other eight inbreds were selected from either local populations and crossing between local and exotic populations on the basis of yielding ability. Self-pollination was practiced for eight generations from 1994 to 2001, at the Agricultural Research Farm of Suez Canal University in Ismailia.
| Table 1: | Pedigree and origin of the parental genotypes |
 | |
| Table 2: | Monthly rainfall (mm), average maximum (Tmax) and minimum (Tmin) temperatures (°C) at Ismailia, Egypt during the experiment's growth |
 | |
The pedigree and origin of the genetic materials are presented in Table 1.
In summer 2008 the inbred lines were topcrossed to both testers to produce 26 topcrosses. In summer 2009, all genetic materials (26 topcrosses, thirteen inbred lines and two testers) were evaluated in a field experiments at two sowing dates (SD), June, 1 and July 1. The monthly temperature ranges during the time of the experiment are given in Table 2.
The experiment was carried out at the experimental farm of Suez Canal University in a randomized complete block design with three replications. Each genotype (line, cross and tester) was planted in one row, 3 m length, 50 cm between rows and 20 cm between hills. Two weeks before planting, 35 m3 ha-1 of organic manure was added and ploughed. Plots received 75 kg P (15.5%) at seeding time and 333 kg N in the form of ammonium nitrate (33.5%) at three splits (two weeks after planting, at anthesis, during grain filling). One week after silking, 111 kg of K (46%) was applied.
Data Recording
Data were collected on a plot basis for days from sowing to 50% tasseling and 50% silking. Ten harvested ears from each entry and replicate were selected to record the following traits on yield and its components: ear length, diameter, number of rows ear-1, grain number row-1, ear weight plant-1, 100 grain weight and grain yield plant-1
Statistical Analysis
Analysis of variance was performed for the measured traits using CropStat software program 7.2 releases for each sowing date and combined over planting dates. Line x tester analysis was used to partition the significant differences among genotypes for each sowing date and for combined data following Kempthorne (1957). Estimates of general combining ability (GCA) for lines, testers and specific combining ability (SCA) for topcrosses were also estimated based on the same model.
RESULTS AND DISCUSSION
Mean Performance
Table 3 and 4 reveal that genotypes and their sub-sources of variation (parents, hybrids and parents vs hybrids) differed significantly for all characters at individual and across sowing dates. The variability among hybrids was less than that among parents for all traits except, ear weight plant-1 and yield plant-1 at optimum, late and combined over dates. The contrast of parents vs crosses was considerable and significant indicating the potential of heterotic effects among hybrids. Combined analysis showed that all investigated traits were affected significantly by changing in sowing date except row ear-1, parents x sowing date except days to 50% silking, genotypes and crosses x sowing date except ear diameter and rows ear-1.
| Table 3: | Line x tester analysis for flowering traits yield and its components of 26 maize topcrosses evaluated at two sowing dates |
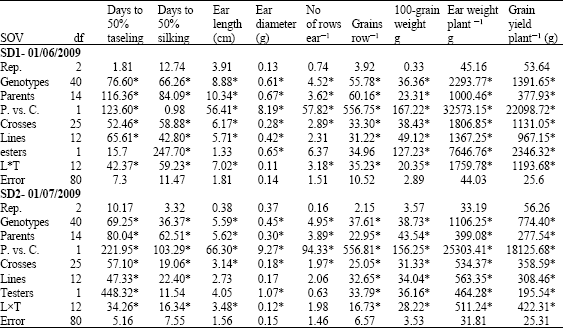 | |
| *Significant at p = 0.05 | |
| Table 4: | Mean squares from line x tester analysis based on data combined over 2 sowing dates for flowering traits, yield and its components of 26 maize topcrosses |
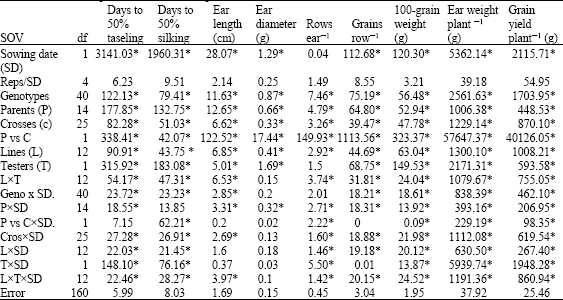 | |
| *Significant at p = 0.05 | |
The presence of parents and crosses x sowing date interactions for the studied traits indicated the different rankings and responses to two sowing dates. Kamara et al. (2009) stated that planting date and cultivars influenced grain yield significantly, in contrast non significant differences of cultivars and planting dates were recorded on days to silking. Similarly, Law-Ogbomo and Remison (2009) recorded a significant effect of sowing date on grain yield and days to 50% flowering. In contrast, Fabijanac et al. (2006) confirmed a non significant effect of sowing date on yield and its components in any of the tested hybrids, consequently tested maize hybrids may be successfully grown at delayed sowing dates due to their shorter than fully season maturity characteristics.
Lines showed significant differences for all studied traits except rows ear-1 at the optimum sowing date and rows ear-1, ear length and diameter at late planting. Significant differences were recorded between testers for days to silking, ear diameter, ear weight plant-1, 100-grain weight and yield plant-1 under optimum conditions. In contrast, at late planting, days to silking, ear length and rows ear-1 did not show significant differences (Table 3). Line x tester mean square revealed non-significant differences for ear diameter at optimum sowing and rows ear-1 at late sowing. The significant effect of line x tester is an indication to the predominance of dominance gene action in the controlling most of the measured traits at both planting dates and the weak effect of additive gene nature.
The view was different when data were combined over sowing dates. Table 4 demonstrate a significant effect of interactions of crosses, lines, testers and line x tester with sowing dates on the measured traits. However, ear diameter was not affected significantly by any interaction, except parents x SD. While, ear length showed non significant effect due to line and tester x SD, grains row-1 was not influenced by tester x SD. The significant effect with SD is an indication for different responses and ranking of lines and testers to sowing dates. The present result was supported by earlier view of Kang (1998) who mentioned a prominent role of environment in phenotypic expression of agronomic traits. Consequently, Bello and Olaoye (2009) suggested that variation in climate (rainfall, sunshine, relative humidity, etc.) could be an important factor in breeding for desirable characters including grain yield.
Means for each of the 26 topcrosses are given in Table 5-9. Most topcrosses recorded lower values for the measured traits at delayed planting.
Late sowing resulted in reducing days to 50% tasseling (9.36 d) and silking (6.87 d), lighter 100-grain weight (6.29%), reduction in grains row-1 (8.28%), ear weight plant-1 (14.90%) and yield plant-1 (12.35%) for the 26 topcrosses and two testers compared to sowing planting date. Similar finding have been observed by Law-Ogbomo and Remison (2009) who reported a highest grain yield from April 7 sown (3.81 t ha-1) compared with May 7 (3.68 t ha-1) and June 7 (3.54 t ha-1). Also, Kamara et al. (2009) recorded significant grain yield reduction by 19 and 28.5% as planting date was delayed to 21 and 28 July, respectively. In contrast, delaying sowing date showed no marked mean reductions on ear length and diameter and rows ear-1 for the same genotypes. Similarly, Rashid et al. (2004) recorded a reduction in plant height, No. of effective tillers plant-1, No. of grains ear-1 and 1000 grain weight in December, 30 seeding compared to December, 15 seeding of wheat. They attributed the reduction in 1000-grain weight to the shorter grain filling period. Regarding flowering traits, the majority of crosses were earlier in days to tasseling and silking than both testers at each sowing date and combined over dates. The most earliest-anthesis topcrosses were L5 and L6 x SC.122, L11 x Pioneer 30B51 at optimum, late and combined plantings. The topcrosses L8 and L10 x SC 122 recorded the lowest days to 50% silking at first sowing and across dates.
| Table 5: | Means for days to 50% tasseling and silking of 26 top crosses and 2 testers of maize evaluated at two sowing dates and combined over dates |
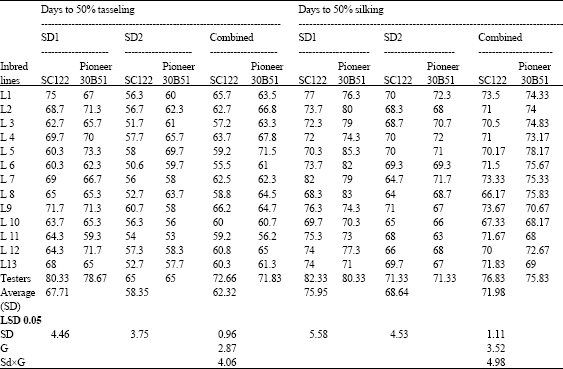 | |
| Table 6: | Means for ear length (cm) and ear diameter (cm) of 26 topcrosses and 2 testers of maize evaluated at two sowing dates and combined over dates |
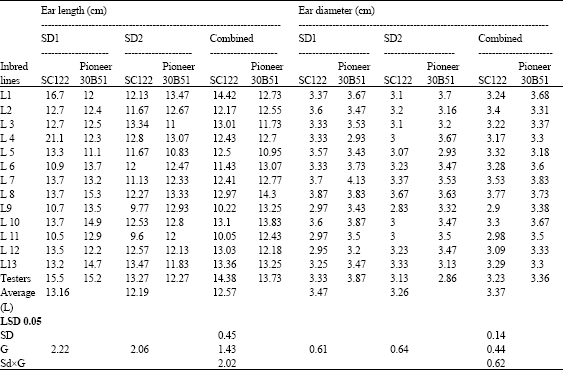 | |
Whereas, the crosses L11 and L 10 x Pioneer 30B51 were the earliest in 50% silking appearance at late and over plantings compared to both testers. In contrast, the topcrosses L5 x Pioneer 30B51 at first sowing was the latest in days to 50% teseling.
| Table 7: | Means for grains row-1 and rows ear-1 of 26 top crosses and testers of maize evaluated at two sowing dates and combined over dates |
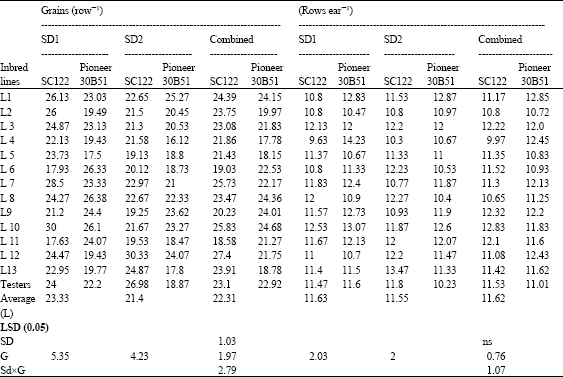 | |
| Table 8: | Means for ear weight plant-1 (g) and 100-grain weight (g) of 26 top crosses and testers of maize evaluated at two sowing dates and combined over dates |
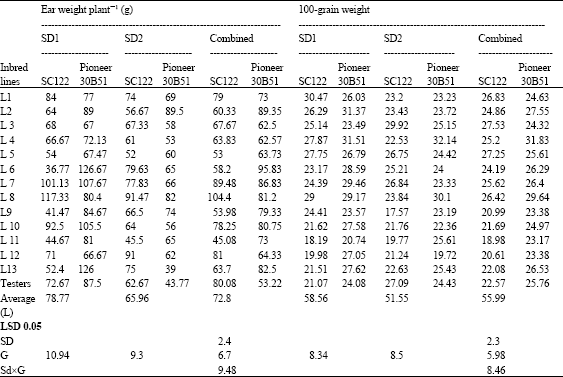 | |
The entries L7 x SC.122 and L5, L6 and L8 x Pioneer 30B51 at June planting, L5 and L7 x Pioneer 30B51 and L5 x Pioneer 30B51, L8 x Pioneer 30B51 at combined sowing reached days to 50% silking after or the same time as the checks.
| Table 9: | Means for grain yield plant-1 (g) of 26 top crosses and testers of maize evaluated at two sowing dates and combined over dates |
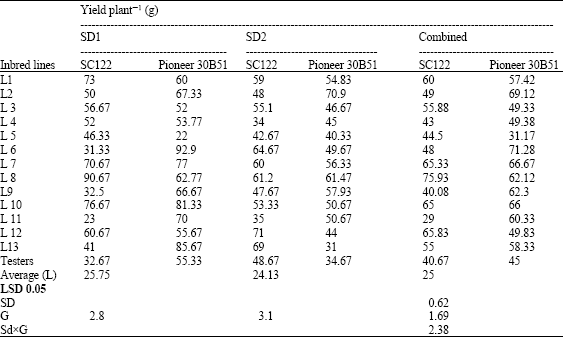 | |
The crosses L1 x SC.122 and L8 x T2 gave the longest ears at optimum planting date, at late sowing the crosses L13 and L3 x SC.122, L1, L8 and L4 x Pioneer 30B51 surpassed the testers. However, the L8 x Pioneer 30B51 cross recorded the highest lengths at optimum, late and over dates. Regarding ear diameter, the crosses involving both lines 7 and 8 with both testers gave the maximum ears at both sowing dates and combined over dates.
The crosses L1 x SC.122 and L8 x Pioneer 30B51 gave the longest ears at optimum planting date, at late sowing the crosses L13 and L3 x SC.122, L1, L8 and L4 x T2 surpassed the testers. However, L8 x Pioneer 30B51 cross recorded the highest lengths at optimum, late and over dates. Regarding ear diameter, the crosses involving both lines 7 and 8 with both testers gave the maximum ear diameters at both sowing dates and combined over dates.
The highest grains row-1 was observed for the entries, L10 x SC.122, L7 X SC.122, L8, L10 and L6 x Pioneer 30B51. At late sowing, the crosses L12 x SC.122, L1 and L12 x Pioneer 30B51 possessed the highest number over both testers. When data were combined over planting dates, the topcrosses L12, L10 and L7 x SC.122 produced the highest grains row-1.
Although, there was no significant effect of sowing date on number of rows ear-1, some crosses recorded slight increase compared to testers (L3 x SC.122, L10 x SC.122, L4 x Pioneer 30B51, L7 x Pioneer 30B51 and L10 x Pioneer 30B51) at optimum date. When sowing was delayed to July 1, the crosses L13 x SC.122, L10 x Pioneer 30B51 and L1 x Pioneer 30B51 showed the highest rows number over testers.
The heaviest 100-grains weight were obtained by the crosses L1 and L8 x SC.122; L4, L7 and L8 X Pioneer 30B51, at optimum planting. At delayed sowing; only L3 x SC.122, L4 and L8 x Pioneer 30B51 were the maximum 100-grain weight. Combined data ranked L3, L5 and L8 x SC.122, L4 and L8 x Pioneer 30B51 as the highest crosses in grain weight.
The highest ear weight plant-1 were significantly recorded for the crosses L8, L7 and L10 x SC.122, L6, L13, L7 and L10 x Pioneer 30B51 over the testers for June planting. Whereas July planting resulted in lowering ear weight for both testers and topcrosses, the entries L8, L12, L6 and L1 x SC.122 and only L8 and L2 x Pioneer 30B51 possessed the highest weights over the both tester.
The topcrosses L7, L8, L10 and L1 x SC122; L6, L13, L10 and L7 x Pioneer 30B51 surpassed both testers in yield plant-1 when planted at optimum date and combined over dates. At late planting, the crosses L12, L13 and L6 x SC.122; L2, L8 and L9 x Pioneer 30B51 produced the highest yield compared to both testers.
Line X Tester Analysis
Combining Ability Effects
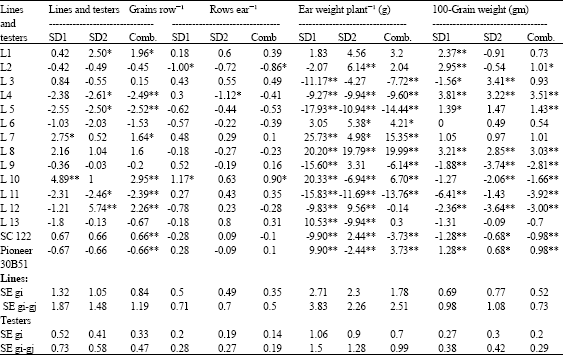
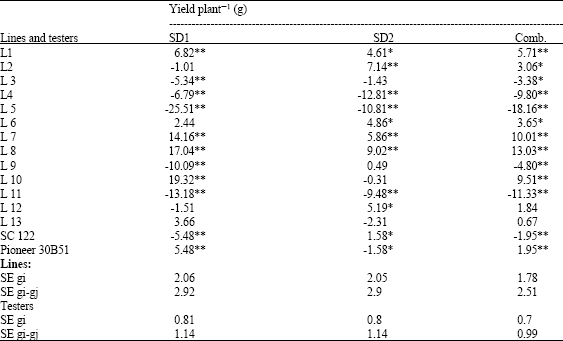
General combining ability effects for all parents are presented in Table 10-12. There is no specific line recorded a desirable GCA effects for all traits at individual and across sowing dates. The inbred lines L7 and L8 showed a significant and high GCA effects for yield plant-1, ear weight plant-1 and ear diameter at both sowing dates and across SD's. On the other hand, L8 recorded positive and significant values for 100-grain weight when corn planted at late and optimal sown. The inbred line 10 demonstrated favorable and significant GCA effects for a group of traits relate with earliness and yield and its components (days to tasseling and silking, yield plant-1, ear weight plant-1, rows ear-1, grains row-1) at optimum and combined over planting dates. The superiority of L10 encourage the use of this line in further breeding programs for hybrid production and synthetic variety characterized by high yield, earliness and yield components. The present result is confirmed by that obtained by Bello and Olaoye (2009) who recorded some populations with significant and desirable GCA effects for yield and flowering traits. Therefore they suggested the possibility of introgression these populations into other promising lines for early flowering and high yielding ability.
Regarding testers, SC 122 presented a significant and/or non significant favorable GCA at late planting for the earliness (days to 50% tasseling and silking), yield and its components (yield plant-1, ear weight plant-1, grains row-1 and rows ear-1). In contrast, Pioneer 30B51 showed desirable and significant GCA values for 100-grain weight, ear length and diameter (positive and non significant effects) at late sown.
| Table 10: | General combining ability effects for days to 50% taseling, 50% silking, ear length and ear diameter for 15 parents evaluated at 2 sowing dates and combined over dates |
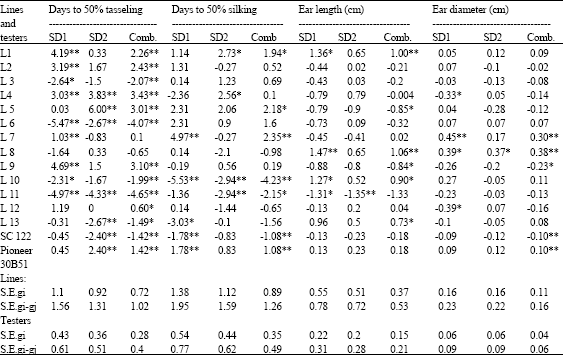 | |
| *Significant at p = 0.05; **Significant at p = 0.01 | |
| Table 11: | General combining ability effects for grains row-1, rows ear-1, ear weight plant-1 and 100-grain weight for 15 parents evaluated at two sowing dates and combined over sowing dates |
 | |
| *Significant at p = 0.05; **Significant at p = 0.01 | |
| Table 12: | General combining ability effects for yield plant-1 for 15 parents evaluated at two sowing dates and combined over sowing dates |
 | |
| *Significant at p = 0.05; **Significant at p = 0.01 | |
Whereas it was not a good combiner for yield plant-1, ear weight plant-1 and flowering traits since it recorded non-desirable effects for these traits.
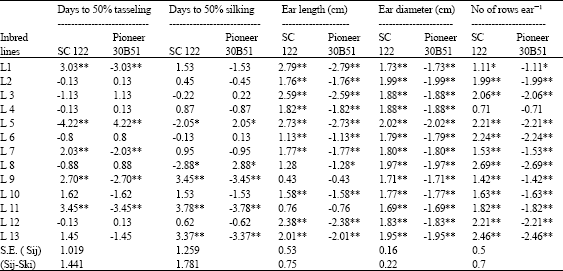
Specific combining ability effects for the 26 topcrosses combined at two sowing dates are illustrated in Table 13 and 14. The topcross L5 X SC.122 showed significant and favorable SCA effects for all measured traits.
| Table 13: | Estimates of SCA effects for flowering traits, yield and yield components for 26 top crosses combined over two sowing dates |
 | |
| *Significant at p = 0.05; **Significant at p= 0.01 | |
| Table 14: | Estimates of SCA effects for grains row-1, 100-grain weight (gm), ear weight plant-1 (g) and yield plant-1 (g) for 26 top crosses combined over two sowing dates |
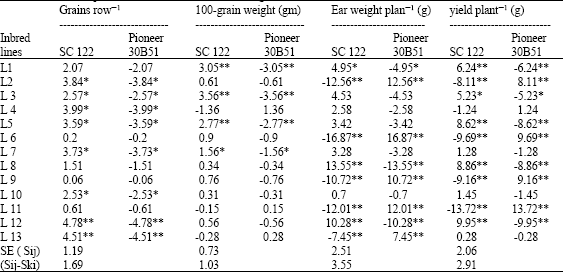 | |
L12 x SC.122 recorded desirable and significant SCA values for ear length and diameter, rows ear-1, grains row-1, ear weight plant-1 and yield plant-1. L8 x SC.122 gave best results for earliness (silking date), ear diameter, rows ear-1, ear weight plant-1 and yield plant-1. L11 and L9 x Pioneer 30B51 showed desirable effects towards earliness, high ear weight plant-1 and yield plant-1. None of entries recorded desirable SCA effects with Pioneer 30B51 for ear length, ear diameter, rows ear-1 and grains row-1. It is interesting that the L11, L9, L5 and L8 are of prime importance for building new maize hybrid combining early flowering and yield characteristics when combined at both optimal and late sowing dates.
Genetic Components
Estimates of genetic components, GCA (for inbred lines and testers) and SCA (for topcrosses) at each sowing date and combined over planting dates for all measured traits are illustrated in Table 15. The results showed that, the magnitude of δ2SCA variance was higher than δ2GCA for all studied traits, except ear diameter at optimum, late and combined over dates.
| Table 15: | Estimates of genetic variance components for flowering traits, yield and its components of line x tester model evaluated at two sowing dates and combined over dates |
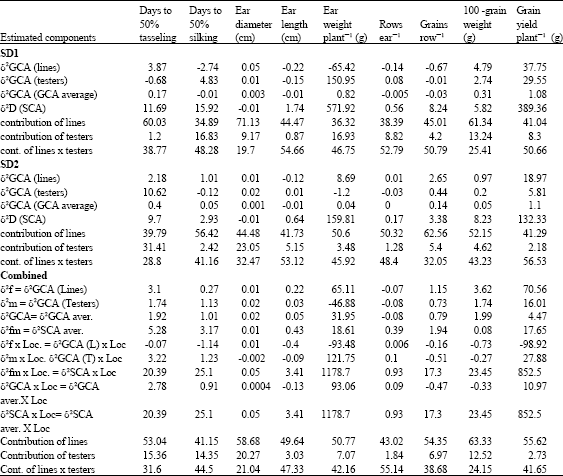 | |
| T: Denote testers, L: Inbred lines and SD: Sowing dates. Any negative value of variances is considered to be equal zero (Robinson et al., 1955) | |
In a study to characterize the genetic architecture of local and CIMMYT corn inbred lines, Sofi and Rather (2006) recorded a significant effect due to progenies, lines, testers, crosses and parents vs crosses for all traits except kernels rows ear-1 where mean square due to lines was not significant. This finding means a substantial variability in parental lines for these traits.
There was a great role of variance due to GCA of lines in controlling days to 50% tesseling, ear diameter, 100-grain weight and yield plant-1 under optimum sowing. At late sowing, GCA variance due to lines was predominant for days to 50% silking, yield plant-1 and its components. The previous results are in partial agreement with that obtained by Iqbal et al. (2007) who found a great importance of dominance gene action in controlling kernels row-1, kernels rows ear-1 and 1000-grain weight. In contrast yield plot-1, ear length and diameter were under the additive gene effect.
The proportional contribution of lines was the highest (tasseling date, ear diameter, 100-grain weight and yield plant-1), followed by line x tester, whereas, the testers' contribution was the least at optimum planting. With delayed sowing, only ear length and yield plant-1 were under the contribution of line x tester, where other traits were controlled by lines. The previous finding is confirmed by the highest GCA values for lines compared to testers and line x tester.
Bello and Olaoye (2009) recorded a high gca/sca ratio and affirmed the preponderance of additive type of gene action in determining grain yield and other agronomic traits. There is a partial agreement with the present findings, which may be due to different breeding materials or environmental variation.
The great contribution of lines in the total variation is an indication for the maternal influence of the inbred lines on the mentioned traits. When data were combined, the same trend was obtained, since inbred lines ranked the highest in their contribution for all traits except, ear diameter, days to 50% silking and rows ear-1. The present results were confirmed with those obtained by Tassawar et al. (2007), who found a greater contribution of inbred lines for ears plant-1, anthesis-silking interval, maturity and seed vigor. While seed setting, plant growth rate and yield plant-1 were more contributed by line x tester, whereas tester contribution constituted the least parental influence. The same authors found a predominant role of non-additive gene action for all measured traits on line x tester crosses and evaluated under moderate temperature stress conditions.
The combined analyses showed also a greater values for δ2SCA X SD than δ2GCA x SD that were either negative or small for all measured traits. This result confirms the influence of environmental conditions (high temperature, short growing season) on non-additive gene effect than additive one. There was great influence of sowing date on δ2GCAT for flowering characters, ear weight plant-1, rows ear-1 and yield plant-1, which confirming the changing of additive gene action for testers with different planting dates.
CONCLUSION
Changes in sowing date had a significant effect on maize growth and yielding ability. Late sowing resulted in means reduction of most traits. All measured traits were affected significantly by parents and crosses x SD interaction, meaning that their order differ with SD change. The magnitude of δ2SCA variance was higher than δ2GCA for the most studied traits, at optimum, late and combined over dates. The proportional contribution of lines was the highest followed by line x tester, whereas, the testers' contribution was the least both sowing dates and combined over sowings. Some parents (L6, L7, L8 and L10) were identified for their desirable GCA effects for grain yield plant-1 and yield components, such lines are promising for incorporation in hybrid formation to improver the aforementioned traits.
REFERENCES
- Akbar, M., M Saleem, F. Muhammad, M.K. Ashraf and R.A. Ahmed, 2008. Combining ability analysis in maize under normal and high temperature conditions. J. Agric. Res., 64: 27-38.
Direct Link - Basbag, S., R. Ekinci and O. Gencer, 2007. Combining ability and heterosis for earliness characters in line x tester population of Gossypium hirsutum L. Hereditas, 144: 185-190.
CrossRefDirect Link - Bello, O.B. and G. Olaoye, 2009. Combining ability for maize grain yield and other agronomic characters in a typical southern guinea savanna ecology of Nigeria. Afr. J. Biotechnol., 8: 2518-2522.
Direct Link - Campos, H., M. Cooper, J.E. Habben, G.O. Edmeades and J.R. Schussler, 2004. Improving drought tolerance in maize: A view from industry. Field Crops Res., 90: 19-34.
Direct Link - Doll, P., 2002. Impact of climate change and variability on irrigation requirements: A global perspective. Clim. Change, 54: 269-293.
Direct Link - Dubey, R.B., V.N. Joshi and N.K. Pandiya, 2001. Heterosis and combining ability for quality, yield and maturity traits in conventional and non-conventional hybrids of maize (Zea mays L.). Ind. J. Genet. Plant Breed., 61: 353-355.
Direct Link - Fabijanac, D., B. Varga, Z. Svecniak and D. Grbesa, 2006. Grain yield and quality of semiflint maize hybrids at two sowing dates. Agric. Cons. Sci., 71: 45-50.
Direct Link - Iqbal, A.M., F.A. Nehvi, S.A. Wani, R. Qadir and Z.A. Dar, 2007. Combining ability analysis for yield and yield related traits in Maize (Zea mays L.). Int. J. Plant Breed. Genet., 1: 101-105.
CrossRefDirect Link - Kamara, A.Y., F. Ekeleme, D. Chikoye and L.O. Omoigui, 2009. Planting date and cultivar effects on grain yield in dryland corn production. Agron. J., 101: 91-98.
CrossRefDirect Link - Law-Ogbomo, K.E. and S.U. Remison, 2009. Growth and yield of maize as influenced by sowing date and poultry manure application. Not. Bot. Hort. Agrobot. Cluj, 37: 199-203.
Direct Link - Maton, L., J.E. Bergez and D. Leenhardt, 2007. Modelling the days which are agronomically suitable for sowing maize. Eur. J. Agron., 27: 123-129.
CrossRef - Sofi, P. and A.G. Rather, 2006. Genetic analysis of yield traits in local and CIMMYT inbred line crosses using line x tester analysis in maize (Zea mays L.). Asian J. Plant Sci., 5: 1039-1042.
CrossRefDirect Link - Rashid, M.H., S.C. Samanta, P. Biswas, M.A. Mannan and A.K.M.M. Zaman, 2004. Performance of thirty wheat genotypes under late sown conditions in Southern region of Bangladesh. Asian J. Plant Sci., 3: 286-289.
CrossRefDirect Link - Robinson, H.F., R.E. Comstock and P.H. Harvey, 1955. Genetic variance in open pollinated varieties of corn. Genetics, 40: 45-60.
Direct Link - Sinsawat, V., J. Pandy, P. Leipner, P. Stamp and Y. Fracheboud, 2004. Effect of heat stress on the photosynthetic apparatus in maize (Zea maize L.) grown at control or high temperature. Environ. Exp. Bot., 52: 123-129.
CrossRef - Tassawar, H., A.K. Iftikhar and A. Zulfiqar, 2007. Study on gene action and combining abilities for thermotolerant abilities of corn (Zea mays L.). Int. J. Plant. Prod., 1: 1-12.
Direct Link