
Go-Eun Hong
Department of Food Science and Biotechnology of Animal Resources, Konkuk University, Seoul, 143-701, Korea
Prabhat Kumar Mandal
Rajiv Gandhi College of Veterinary and Animal Sciences, Puducherry, 605 009, India
Ki-Won Lim
Department of Physical Education, Konkuk University, Seoul, 143-701, Korea
Chi-Ho Lee
Department of Food Science and Biotechnology of Animal Resources, Konkuk University, Seoul, 143-701, Korea
Asian Journal of Animal and Veterinary Advances
Year: 2012 | Volume: 7 | Issue: 6 | Page No.: 502-511







ABSTRACT
Isoflavone aglycone content increases after fermentation in soy pulp which is known for prevention of postmenopausal disease. The purpose of this study was to establish optimum fermentation conditions and suitable culture for mass production of aglycone using black soybean pulp, a by-product of soybean processing. Black soybean pulp was fermented up to 48 h using Lactobacillus acidophilus (LA) and Bacillus subtilis (BS) culture. Samples were collected every 12 h to measure isoflavones (daidzin and genistin and daidzein and genistein) using High-performance Liquid Chromatography (HPLC) and β-glucosidase activity using spectrophotometry. Isoflavone aglycone content was verified by Liquid Chromatography-tandem Mass Spectrometry (LC-MS/MS) analysis. Genistein concentrations in black soybean pulp after 12 and 24 h of fermentation with L. acidophilus were 6.8 and 7.2 fold, respectively, compared to controls. Genistein concentrations in black soybean pulp after 12 and 24 h of fermentation with B. subtilis were 6.8 and 6.9 fold greater than controls. β-glucosidase activity was highest in the black soybean pulp fermented for 12 h with L. acidophilus. Black soybean pulp fermented for 12 h with L. acidophilus was most efficient to produce isoflavone aglycone.
PDF Abstract XML References Citation
Received: October 19, 2011;
Accepted: December 20, 2011;
Published: February 17, 2012
How to cite this article
Go-Eun Hong, Prabhat Kumar Mandal, Ki-Won Lim and Chi-Ho Lee, 2012. Fermentation Increases Isoflavone Aglycone Contents in Black Soybean Pulp. Asian Journal of Animal and Veterinary Advances, 7: 502-511.
DOI: 10.3923/ajava.2012.502.511
URL: https://scialert.net/abstract/?doi=ajava.2012.502.511
DOI: 10.3923/ajava.2012.502.511
URL: https://scialert.net/abstract/?doi=ajava.2012.502.511
INTRODUCTION
Phenolic compounds from soybeans known as isoflavones exist in glycoside or aglycone (without the glycoside part) forms (Ogbuewu et al., 2010). Daidzin and genistin are the two major isoflavones found in non-fermented soybean products and daidzein and genistein are two major soybean isoflavones aglycone produced by bacterial fermentation (Chukeatirote et al., 2010). During fermentation microbial enzymes break the carbohydrate bond and increases isoflavone aglycone content (Wang and Murphy, 1996; Choi and Sohn, 1998; Hong et al., 2009). While all isoflavones are absorbed into the mucosa of the small intestine, the aglycone form is absorbed at a greater rate and also has higher antioxidant activity than the glycoside form (Izumi et al., 2000; Setchell et al., 2001; Chen et al., 2002; Amadou et al., 2009). Lipolytic effects of genistein and daidzein in adipocytes derived from normal diet-fed rats and high fat diet-fed rats has been reported (Pulbutr and Rattanakiat, 2010). Therefore, it is more effective to partake food containing Isoflavone in the aglycone form than the glycoside form for disease prevention (Izumi et al., 2000; Setchell et al., 2001). The isoflavone phytoestrogen has a structure and biological actions similar to that of the female hormone 17 β-estradiol (Ogbuewu et al., 2010).
Soflavones are known for having anticancer activity and an effect on cell cycle and growth control (Katadare et al., 2002; Kim et al., 2002; Gourineni et al., 2010). They have also been associated with the prevention or treatment of hypertension and cardiovascular diseases and hormone associated disorders, in particular postmenopausal syndrome, osteoporosis, breast cancer and colorectal cancer (Moon et al., 1996; Choi and Sohn, 1997, 1998; Cassidy, 1996; Kim et al., 2000; Lee et al., 2002; Choi et al., 2006). Among the soybean types, black soybeans (Glycine max L. Merr.) are known for their high isoflavone concentration and physiologic activity (Kim et al., 1992; Shon et al., 2000; Ryu and Moon, 2003). Black soybeans have long been highly regarded as folk remedies for their pharmacological effects. In addition, bean pods are known to have a great deal of isoflavone or anthocyanin (Adlercreutz et al., 1994). Along with increasing life expectancy, people who use these remedies are found to be very healthy.
Soy pulp is generated as a by-product during the production of tofu or soymilk. Approximately 1.1 kg of fresh soybean pulp is produced from every kg of soybeans processed for soymilk (Khare et al., 1995). Soy pulp is sometimes used as animal food but mostly it is burnt as waste (Ohno et al., 1993; Kwon et al., 1998; Otoole, 1999). The anti osteoporosis effect of isoflavone aglycone of fermented soy pulp has been reported (Hong et al., 2009). Effect of temperature and processing methods on biochemical changes induced by Bacillus subtilis during fermentation of soybean into condiment (Soy-daddawa) has been reported (Omafuvbe et al., 2007; Omafuvbe, 2008). The demand for functional health foods is increasing very fast both in the internal and external market. The importance of isoflavones in particular has increased because of its preventive effect on the diseases associated with hormone deficiency in postmenopausal women (Hanachi and Golkho, 2008; Radhakrishnan et al., 2009). The low yield of the isoflavone compared to the price has caused distribution difficulties (Kim et al., 2008). To address this issue, we designed this study to find the optimum condition for fermentation of black soybean pulp using Lactobacillus acidophilus or Bacillus subtilis with the aim of producing isoflavone aglycone in more quantity.
MATERIALS AND METHODS
This work was done during September, 2009 to February, 2011.
Materials and reagents: In this study, black soybean (Korean cultivar; Seoritae) was purchased at a local market in Hwayang-dong, Seoul, Korea. Daidzin, genistin, daidzein and genistein were purchased from Sigma-Aldrich (Korea) and HPLC grade ethanol, methanol and acetonitrile were purchased from J.T. Baker Co. (Phillipsburg, NJ, USA).
Black soybean pulp production: Black soybeans were soaked in water in 3:1 ratio over 8 h. Black soybean pulp was obtained after grinding and removing the milk with muslin cloth and freeze-dried.
Fermentation of soy pulp: Pure cultures of Lactobacillus acidophilus (ATCC4356) and Bacillus subtilis (KCCM12511) were obtained from Korean Collection for Type Culture (Daejeon, Korea). L. acidophilus (LA) was activated in de Mann Rogosa Sharpe (MRS) broth (Oxoid LTD., Basingstoke, Hampshire, England) at 37°C aerobically over 24 h (De Man et al., 1960), then activated cultures were inoculated into MRS broth at 37°C for 48 h. B. subtilis (BS) was activated in Nutrient Broth (DifcoTM, USA) at 30°C aerobically over 24 h. For fermentation, pre-activated culture was inoculated into 1000 mL flasks with black soybean pulp (5%, v/v) and incubated at 37°C in case of L. acidophilus and at 30°C in case of B. subtilis for 48 h. Samples were collected every 12 h and stored at -80°C after freeze-drying.
Proximate analysis: Moisture, crude protein, crude fat, ash and crude fiber were estimated in the fermented and non-fermented black soybean pulp. Carbohydrate content was calculated by deducting the sum total of moisture, crude protein, crude fat, ash and crude fiber content from 100.
Sample preparation for analysis of isoflavone: The extraction of isoflavone including daidzin, genistin, daidzein and genistein from fermented and non-fermented black soybean pulp for HPLC analysis was performed using a modified method described by Chung et al. (2008) and Lee et al. (2008). A 0.5 g freeze-dried sample in 50 mL of 80% ethanol was extracted using a Branson 3210 Sonicator (Branson Ultrasonics Co., CT, USA) for 60 min. Supernatant was collected after centrifugation (3000xg) for 20 min and repeat filtered through Whatman filter paper No. 1 into a 50 mL tube. The filtrate was concentrated using rotary vacuum evaporator (EYELA Co., USA) and resuspended in 5 mL of 80% methanol. Sample liquids were filtered with syringe filters (0.2 μm, Whatman Co., England) for HPLC analysis.
HPLC analysis of isoflavone: HPLC analysis was done in an Agilent 1100 Series HPLC system equipped with an autosampler, diode array and multiple wavelength detectors with ChemStation software. HPLC separation was performed on a BondapakTMC18 column (300x3.9 mm). Experimental conditions for HPLC were followed as per the modified method of Chung et al. (2008) and GolKhoo et al. (2008). Mobile phases were 0.1% acetic acid in water (A) and 0.1% acetic acid in acetonitrile (B) in gradient. Linear gradient profile was from 15 to 35% B in 50 min. The flow rate was 1.0 mL min-1. The wavelength of UV detection was 254 nm. The four isoflavone standard solution of daidzin, genistin, daidzein and genistein were prepared in 80% methanol at 0.1 mg mL-1 concentration for peak identification. All isoflavone concentrations were calculated back to dry weight basis (mg isoflavone 100 g-1 black soybean pulp).
Determination of β-glucosidase activity: β- glucosidase activity was determined according to the method described by Matsuura et al. (1995) using ρ-nitropheno-β-D-glucopyranoside (ρ-NPG) as the synthetic substrate. A volume of 1.5 mL of 0.05 M citrate buffer (pH 4.5) containing 0.1 M NaCl was added to 100 mg samples and allowed to settle for 1 h at room temperature to extract β-glucosidase. Samples were then centrifuged (8000xg) for 30 min and supernatant collected to measure enzyme activity. A 2 mL volume of 0.1 M phosphate-citrate buffer (pH 5.0) containing 1 mM ρ-NPG was placed in a test tube in a water bath at 30°C for 10 min. A total of 0.5 mL of supernatant was added to the test tube and left to settle for 30 min at 30°C, after which 2.5 mL of 0.5 M sodium carbonate was added to stop the reaction. The amount of ρ-nitrophenol released during the reaction was determined with a spectrophotometer (Optizen 2120UV, Mecasys Co. Ltd., Korea) at 420 nm. A ρ-nitrophenol calibration curve was previously prepared to calculate the enzyme activity. A unit of activity was defined as the amount of β-glucosidase in 1 μM of ρ-nitrophenol per minute under the current experimental conditions. The results are described as UA g-1 of whole black soybean pulp in dry weight basis.
According to the HPLC analysis of isoflavone and β-glucosidase activity, the sample fermented for 12 h using L. acidophilus had the highest yield efficiency (Fig. 1, 2). Therefore, subsequent experiments were performed using black soybean pulp fermented with L. acidophilus for 12 h after freeze-drying.
LC/MS-MS analysis: LC/MS-MS analysis was performed with an Agilent 1200 Series G1367D (Agilent Co., USA), autosampler combined with an Agilent 6410 Triple-Quadrupole using a modified method described by Holder et al. (1999). The LC column was a Hypersil BDS-C18 (4.0x100 mm, 3 μm, Agilent Co.), flow rate was 0.5 mL min-1 and injection volume was 10 μL. The LC mobile phase solvent A was 0.1% formic acid and solvent B was acetonitrile set as an isocratic elution (65% formic acid (0.1%)/35% acetonitrile). Ionization was performed in the positive mode of Electro Spray Ionization (ESI) and ion detection was in MRM (multiple reaction monitoring)mode. Nebulizing gas (N2) temperature was 320°C, gas flow was 12 mL min-1 and capillary voltage was 4000 V. Each isoflavone ion’s optimizing condition determined using the Agilent Co., optimizer program is shown in Table 1.
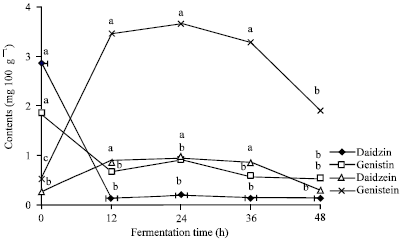 | |
| Fig. 1: | Isoflavone contents in black soybean pulp fermented with L. acidophilus as estimated by HPLC |
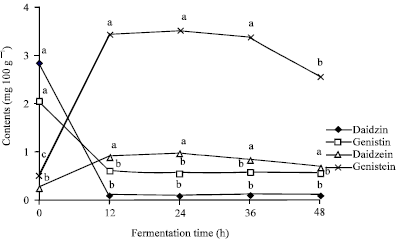 | |
| Fig. 2: | Isoflavone contents in black soybean pulp fermented with B. subtilis as estimated by HPLC |
| Table 1: | Conditions for isoflavone analysis by LC-MS/MS in MRM mode |
 | |
| Table 2: | Proximate composition of fermented and non-fermented black soybean pulp |
 | |
| 1Black soybean pulp fermented with L. acidophilus, 2Black soybean pulp fermented with B. subtilis | |
Statistical analysis: Each experiment was repeated three times. All data collected were subjected to analysis of variance (ANOVA) after estimation of means and Standard Error of the Mean (SEM) with the Statistical Analysis System (SAS, Version 9.1 TS Level 1M3). A Turkey test was used to determine which means were significant with a minimal 5% probability difference (p<0.05).
RESULTS AND DISCUSSION
Results of proximate analysis: The proximate analysis of fermented and non-fermented black soybean pulp is shown in Table 2. Moisture content was 0% in both fermented and non-fermented black soy pulp as freeze-drying was done to the maximum intensity. Crude fat content was 28.46% in non-fermented black soybean pulp and 31.90 and 35.01% in LA and BS fermented black soybean pulp, respectively. The increase in crude fat content might be due to bacterial fermentation. Crude protein content was 12.94% in non-fermented black soybean pulp and 16.25 and 15.43% in LA and BS fermented black soybean pulp, respectively. Ash content without fermentation was 5.5, 5.2 in LA and 5.0% in BS. Crude fiber also decreased with fermentation from 14.33 to 11.21 and 11.29%. The contents of crude fat and crude protein were increased after fermentation. Choi et al. (2010) obtained similar results with rice bran fermented by B. subtilis, Lactobacillus casei and Saccharomyces cerevisiae. They reported increased protein content and decreased carbohydrate content after fermentation. Ye and Bae (2010) also reported similar results of decreased carbohydrate content and increased protein and fat contents during fermentation of mulberry leaves.
HPLC analysis of isoflavone
Black soybean pulp fermented with L. acidophilus: The isoflavone content of black soybean pulp fermented with L. acidophilus is presented in Fig. 1. The isoflavone (daidzin, daidzein, genistin and genistein) concentrations in non-fermented black soybean pulp were 2.85, 0.27, 1.85 and 0.51 mg 100 g-1, respectively. Daidzin concentration significantly (p<0.05) decreased during fermentation to 0.13 mg 100 g-1 at 48 h. There were no significant difference in daidzin contents between 12, 24, 36 and 48 h. Daidzein concentration significantly (p<0.05) increased after 12 h and there were no significant difference between 12, 24 and 36 h of fermentation from 0.89 to 0.96 mg 100 g-1 and decreased after 36 and 48 h from 0.86 to 0.32 mg 100 g-1. There were no significant difference in daidzein contents between 12, 24 and 36 h. Genistin concentration was 0.67 mg 100 g-1 in the 12 h fermented sample and slightly increased in the 24 h fermented sample to 0.91 mg 100 g-1 but decreased after 36 and 48 h of fermentation to 0.59 and 0.56 mg 100 g-1, respectively. Genistein concentration increased after 12 and 24 h of fermentation from 3.45 to 3.66 mg 100 g-1 but decreased after 36 and 48 h to 3.28 and 1.91 mg 100 g-1, respectively. Total isoflavone and aglycone contents were highest in 24 hr fermented sample but there were no significant (p<0.05) differences between 12 h (Total-5.1 mg 100 g-1; aglycone-4.3 mg 100 g-1) and 24 h (Total, 5.7 mg 100 g-1, aglycone, 4.6 mg 100 g-1) fermented samples.
Black soybean pulp fermented with B. subtilis: The isoflavone content of black soybean pulp fermented with B. subtilis is presented in Fig. 2. The isoflavone (diadzin, daidzein, genistin and genistein) concentrations in non-fermented b lack soybean pulp were 2.84, 0.26, 2.05 and 0.50 mg 100 g-1, respectively. Daidzin concentration gradually decreased to 0.13 mg 100 g-1 during fermentation. Daidzein concentration increased from 12 to 24 h of fermentation from 0.90 to 0.95 mg 100 g-1 and decreased after 36 and 48 h from 0.86 to 0.69 mg 100 g-1, respectively. Genistin concentration consistently decreased during fermentation from 0.60 to 0.57 mg 100 g-1. Genistein concentration increased from 12 to 24 h of fermentation from 3.41 to 3.50 mg 100 g-1 and decreased after 36 and 48 h from 4.91 to 3.94 mg 100 g-1, respectively. Total isoflavone content is highest in 0 hr fermented sample (Total 5.7 mg 100 g-1, aglycone 0.8 mg 100 g-1). Aglycone content is the highest in 24 h fermented sample but there were no significant (p<0.05) differences between 12h (Total 5.0 mg 100 g-1, aglycone 4.3 mg 100 g-1) and 24 h (Total 5.1 mg 100 g-1, aglycone 4.4 mg 100 g-1) fermented sample.
β-glucosidase activity: β-glucosidase activity results are shown in Fig. 3. β-glucosidase activity in black soybean pulp fermented by L. acidophilus significantly (p<0.05) increased from 1070 UA g-1 powder before fermentation to 2140 UA g-1 powder after 12 h of fementation. In the case of black soybean pulp fermented by B. subtilis, β-glucosidase activity significantly (p<0.05) decreased at 12 h and the highest level (2030 UA g-1 powder) was at 24 h and then significantly (p<0.05) decreased after 24 h. According to these results, 12 h fermentation by L. acidophilus and 24 h fermentation by B. subtilis was found to have maximum enzyme activity.
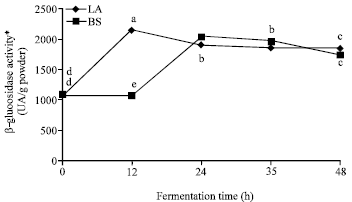 | |
| Fig. 3: | β-glucosidase activity in fermented black soybean pulp, LA : Black soybean pulp fermented with L. acidophilus, BS : Black soybean pulp fermented with B. subtilis, *Enzyme activity was defined as the amount of β-glucosidase that released 1 μ moL of p-nitrophenol from the substrate p-nitrophenyl-D-glucopyranoside per minute (n = 5) |
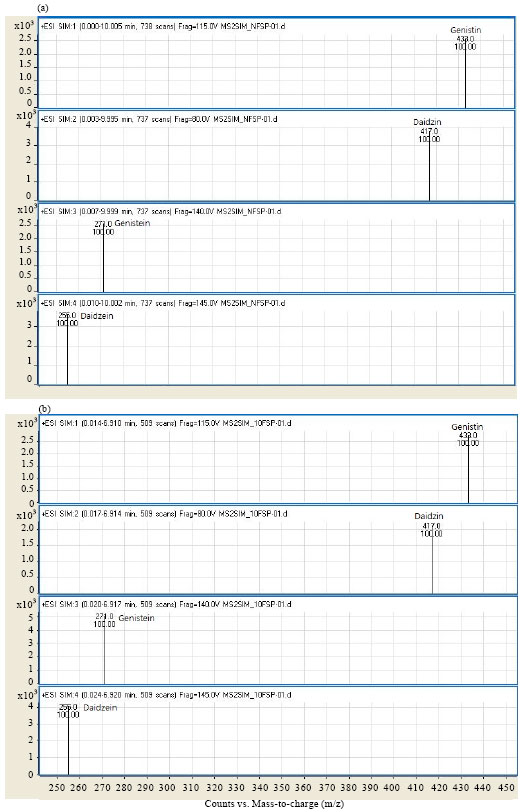 | |
| Fig. 4: | The change of isoflavone concentration spectrum in MRM mode for (a)non-fermented black soybean pulp and (b) black soybean pulp fermented with L. acidophilus for12 h |
LC/MS-MS analysis: Each of the four isoflavone spectra was acquired for quantitative analysis. Each precursor ion at daidzin (m/z 417), daidzein (m/z 255), genistin (m/z 433) and genistein (m/z 271) was selected for protonated molecules, [M+H]+ in positive mode. To find characteristic fragment ions for individual peaks by product ion scan of daidzin (m/z 255), daidzein (m/z 91), genistin (m/z 271) and genistein (m/z 153) were chosen. The standard curve of LC-MS/MS from the peak area shows reliable linearity with a high coefficient of correlation (r2>99%) daidzin-0.998, daidzein-0.999, genistin-0.995 and genistein-0.996. Throughout the LC-MS/MS analysis spectrum in MRM mode, the protonated molecule of genistin (433) and daidzin (417) were decreased and daidzein (255) and genistein (271) were increased compared to pre-fermentation (Fig. 4).
Isoflavone has a variety of types of glycosides including malonyl and acetyl forms. Using LC-MS/MS, more accurate quantitative analysis is attainable when protonated molecular weight is measured as well as the characteristics of product ions. On the basis of this LC-MS/MS quantitative data analysis, the isoflavone aglycone concentration of black soybean pulp fermented by L. acidophilus for 12 h increased 2-fold for genistein and 1.2-fold for daidzein compared to non-fermented samples. These results are similar to our previous report (Hong et al., 2009) in which the isoflavone aglycone concentration of fermented soybean pulp was found to increase during fermentation and the optimum time was 12 h. β-glucosidase activity also was found to be optimum after 12 h of fermentation in our previous report (Hong et al., 2009).
CONCLUSION
The results suggest that the optimum condition to increase the isoflavone content in black soybean pulp is 12 h fermentation with L. acidophilus. This study indicated the possibility of mass production of isoflavone aglycone by fermentation of soybean pulp, a byproduct of soybean processing. Therefore, fermented black soybean pulp could be used as a functional food especially for postmenopausal women to prevent osteoporosis. Further study of mass production of isoflavone aglycone from soybean and its efficacy on prevention of diseases such as osteoporosis caused by estrogen deficiency in human subject is necessary.
ACKNOWLEDGMENT
This research was supported by iPET, Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries, Republic of Korea.
REFERENCES
- Adlercreutz, H., T. Fotis, S. Watanabe, J, Lampe, K. Wahala, T. Makela and T. Hase, 1994. Determination of lignans and isoflavonoids in plasma by isotope dilution gas chromatography-mass spectrometry. Cancer Detect. Prev., 18: 259-271.
PubMedDirect Link - Amadou, I., S. Yong-Hui, J. Sun and L. Guo-Wei, 2009. Fermented soybean products: Some methods, antioxidants compound extraction and their scavenging activity. Asian J. Biochem., 4: 68-76.
CrossRefDirect Link - Chen, C.Y., G.I. Holtzman and R.M. Bakhit, 2002. High-genistin isoflavone supplementation modulated erythrocyte antioxidant enzymes and increased running endurance in rats undergoing one session of exhausting exercise-A pilot study. Pak. J. Nutr., 1: 1-7.
CrossRefDirect Link - Choi, Y.B. and H.S. Sohn, 1998. Isoflavone content in Korean fermented and unfermented soybean foods. Korean J. Food Sci. Technol., 30: 745-750.
Direct Link - Choi, Y.B. and H.S. Sohn, 1997. Recovery of isoflavones from soybean cooking water produced during soymilk manufacturing process. Korean J. Food Sci. Technol., 29: 522-526.
Direct Link - Chukeatirote, E., K. Dajanta and A. Apichartsrangkoon, 2010. Thua nao, Indigenous Thai fermented soybean: A review. J. Biol. Sci., 10: 581-583.
CrossRefDirect Link - Chung, W.Y., S.K. Kim and J.Y. Son, 2008. Isoflavone contents and physiological activities of soybeans fermented with Aspergillus oryzae or Basillus natto. J. Korean Soc. Food Sci. Nutr., 37: 141-147.
Direct Link - GolKhoo, S., A.R. Ahmadi, P. Hanachi, F. Barantalab and M. Vaziri, 2008. Determination of daidzein and genistein in soy milk in Iran by using HPLC analysis method. Pak. J. Biol. Sci., 11: 2254-2258.
CrossRefPubMedDirect Link - Gourineni, V.P., M. Verghese and J. Boateng, 2010. Anticancer effects of prebiotics synergy1® and soybean extracts: Possible synergistic mechanisms in caco-2 cells. Int. J. Cancer Res., 6: 220-233.
CrossRefDirect Link - Hanachi, P. and S. Golkho, 2008. Assessment of soy phytoestrogens and exercise on lipid profiles and menopause symptoms in menopausal women. J. Boil. Sci., 8: 789-793.
CrossRefDirect Link - Holder, C.L., M.I. Churchwell and D.R. Doerge, 1999. Quantification of soy isoflavones, genistein and daidzein and conjugates in rat blood using LC/ES-MS. J. Agric. Food Chem., 47: 3764-3770.
CrossRefDirect Link - Hong, G.E., P.K. Mandal, C.W. Pyun, K. Choi and S.K. Kim et al., 2009. Isoflavone aglycone from fermented soy pulp prevents osteoporosis in ovariectomized rats. Asian J. Anim. Vet. Adv., 4: 288-296.
CrossRefDirect Link - Izumi, T., M.K. Piskula, S. Osawa, A. Obata and K. Tobe et al., 2000. Soy isoflavone aglycones are absorbed faster and in higher amounts than their glucosides in humans. J. Nutr., 130: 1695-1699.
PubMedDirect Link - Katadare, M., M. Osborne and N.T. Telang, 2002. Soy isoflavone genistein modulates cell cycle progression and induces apoptosis in HER-2/neu oncogene expressing human breast epithelial cells. Int. J. Oncol., 21: 809-815.
PubMedDirect Link - Khare, S.K., J. Krishna and A.P. Gandhi, 1995. Citric acid production from okara (soy-residue) by solid-state fermentation. Bioresour. Technol., 54: 323-325.
Direct Link - Kim, S.D., Y.H. Kim, S.H. Lee and E.H. Hong, 1992. Characteristics of black soybean for cooking with rice in Korea. Korea Soybean Digest, 9: 1-13.
Direct Link - Kim, M.H., A.M. Gutierrez and R.H. Goldfarb, 2002. Different mechanisms of soy isoflavones in cell cycle regulation and inhibition of invasion. Anticancer Res., 22: 3811-3817.
PubMedDirect Link - Lee, K.J., K.H. Row and I.C. Jun, 2008. Preparative separation of isoflavones from Korean soybean by HPLC. Asian J. Scientific Res., 1: 288-292.
CrossRefDirect Link - De Man, J.C., M. Rogosa and M.E. Sharpe, 1960. A medium for the cultivation of Lactobacilli. J. Applied Bacteriol., 23: 130-135.
CrossRefDirect Link - Ogbuewu, I.P., A.A. Omede, O.K. Chukwuka, O.O.M. Iheshiulor and M.C. Uchegbu et al., 2010. The overview of the chemistry, health benefits and the potential threats associated with prolonged exposure to dietary soy isoflavones. Int. J. Agric. Res., 5: 1084-1099.
CrossRefDirect Link - Ohno, A., T. Ano and M. Shoda, 1993. Production of the antifungal peptide antibiotic, iturin by Bacillus subtilis NB22 in solidstate fermentation. J. Ferment. Bioeng., 75: 23-27.
CrossRefDirect Link - Omafuvbe, B.O., 2008. Effect of temperature on biochemical changes induced by Bacillus subtilis (SDA3) during starter culture fermentation of soybean into condiment (soy-daddawa). Am. J. Food Technol., 3: 33-41.
CrossRefDirect Link - Omafuvbe, B.O., E.O. Esosuakpo, T.S. Oladejo and A.A. Toye, 2007. Effect of soaking and roasting dehulling methods of soybean on Bacillus fermentation of soy-daddawa. Am. J. Food Technol., 2: 257-264.
CrossRefDirect Link - OToole, D.K., 1999. Characteristics and use of okara, the soybean residue from soy milk production a review. J. Agric. Food Chem., 47: 363-371.
CrossRefDirect Link - Pulbutr, P. and S. Rattanakiat, 2010. Lipolytic effects of genistein and daidzein in adipocytes derived from normal diet-fed rats and high fat diet-fed rats. J. Biol. Sci., 10: 754-760.
CrossRef - Radhakrishnan, G., N.A. Rashmi and N.B. Vaid, 2009. Evaluation of isoflavone rich soy protein supplementation for ostmenopausal therapy. Pak. J. Nutr., 8: 1009-1017.
CrossRefDirect Link - Ryu, S.H. and G.S. Moon, 2003. Antioxidative and antiaging effects of dietary yellow and black soybean in rats. J. Korean Soc. Food Sci. Nutr., 32: 591-597.
Direct Link - Setchell, K.D., N.M. Brown, P. Desai, L. Zimmer-Nechemias and B.E. Wolfe et al., 2001. Bioavailability of pure isoflavones in healthy humans and analysis of commercial soy isoflavone supplements. J. Nutr., 131: 1362S-1375S.
PubMed - Wang, H.J. and P.A. Murphy, 1996. Mass balance study of isoflavones during soybean processing. J. Agric. Food Chem., 44: 2377-2383.
CrossRefDirect Link - Ye, E.J. and M.J. Bae, 2010. Comparison of components between mulberry leaf tea and fermented mulberry leaf tea. J. Korean Soc. Food Sci. Nutr., 39: 421-427.
Direct Link