
Sabri Yurtseven
Department of Animal Sciences, Faculty of Agriculture, Harran University, 63200 Sanliurfa, Turkey
Irfan Ozturk
Department of Animal Sciences, Faculty of Agriculture, Harran University, 63200 Sanliurfa, Turkey
Asian Journal of Animal and Veterinary Advances
Year: 2009 | Volume: 4 | Issue: 2 | Page No.: 76-85







ABSTRACT
This study was performed to evaluate the effect of different cereal source in choice feeding systems on performance and on emission of carbon dioxide (CO2) and enteric methane (CH4) in dairy Awassi ewes. Total 16 dairy ewes were divided into two groups: the corn based free choice (CFC) group received feed ingredients separately (corn, wheat bran, soybean meal (SBM), cottonseed meal (CSM) and alfalfa hay) and the barley based free choice group (BFC) group received barley instead of corn as carbon hydrate source. The results showed no significant differences (p > 0.05) between treatments in live weight, live weight gain, milk yield, milk composition and CO2 production. However, the results of CH4 measurement indicated significant differences between groups in the amounts of CH4 produced. The ewes in the CFC group produced less CH4 than the ewes that received the BFC system (CFC: 21.82; BFC: 38.34 g/day/sheep). The results indicate that the CFC system modified ruminal fermentation and affected the Volatile Fatty Acid (VFA) components and levels in ruminal fluid. In ewes on the CFC system, the level of propionate was greatly increased (CFC: 19.77 vs. BFC: 14.53%) and the level of acetate decreased (CFC: 68.34 vs. BFC: 75.58%). Butyrate level was not changed relative to the total VFA components. There were no significant differences in ruminal pH level between treatments. The results indicate that the CFC system has a potential mitigating effect on enteric emission of CH4 but not CO2.
PDF Abstract XML References Citation
How to cite this article
Sabri Yurtseven and Irfan Ozturk, 2009. Influence of Two Sources of Cereals (Corn or Barley), in Free Choice Feeding on Diet Selection, Milk Production Indices and Gaseous Products (CH4 and CO2) in Lactating Sheep. Asian Journal of Animal and Veterinary Advances, 4: 76-85.
DOI: 10.3923/ajava.2009.76.85
URL: https://scialert.net/abstract/?doi=ajava.2009.76.85
DOI: 10.3923/ajava.2009.76.85
URL: https://scialert.net/abstract/?doi=ajava.2009.76.85
INTRODUCTION
Domestic livestock produce 80-115 million tons of methane per year (IPCC, 2001). Animal nutritionists have focused attention on methods to mitigate methane emission because of its energy production loss as gaseous products in ruminant and the role of methane in global warming. Ruminants typically lose 2-12% of their gross energy intake as emissions of methane (Johnson et al., 1995). In the ruminant industry, decreased production of CH4 can represent an improvement in feed efficiency. Carbon dioxide and methane is the main greenhouse gases related to animal nutrition and methane has greater global warming potential than carbon dioxide (IPCC, 1996). The main source of CH4 is hydrogen, which is produced from fermentative reactions in ruminal fluid. Excess hydrogen must be utilized in order to decrease the production of methane. Since methane production is negatively correlated with total VFA and the proportion of propionate (Wang et al., 2009), improvements in ruminal fermentation that favor propionic acid may also allow a decrease in methane production, because propionic acid contains more hydrogen than other Volatile Fatty Acids (VFA). For this reason, the pattern of ruminal fermentation may be changed by the use of different cereal sources that encourage effective utilization of the other feed ingredients. Feeding systems that allow choice may provide one mechanism by which ruminants can select optimum feed ingredients especially cereal sources to balance ruminal fermentation in favor of the propionic acid. It is well known that the degradation rate of cereal starch differs based on the source (corn starch less degradable than barley) which in turn may modify ruminal fermentation pattern (Schimidely et al., 1999). When the ruminant animal consumes high amount of concentrate, low ruminal pH may reduce fiber digestion (Santini et al., 1992). In addition, a higher content of fibrous carbohydrates (NDF) in diets of low feeding value has been shown by Moe and Tyrell (1979) to increase the methane emitted per unit feed digested. An increase in the concentrate ratio may also improve the nutritional status of sheep by standardizing consumption of energy, protein and mineral and will also give an opportunity to increase the ratio of propionic to acetic acid. Supplementation of diets with concentrate diminishes enteric methane production from cows (Hindrichsen et al., 2005). Ruminants can learn the physiological consequences of ingestion of particular foods and can recognize the feed ingredients offered as free choice according to their post-ingestive effects. Thus, ruminants can consume high concentrate feed without suffering from metabolic disorders (Fedele et al., 2002; Yurtseven and Gorgulu, 2004, 2007). The chemical variation of cereal is the first theme of methane emission differences among rations, which may affect digestibility and consequently feed intake. Theoretically easily fermentable carbohydrates have been suggested to have a significant reducing effect compared to slow fermentable carbohydrates. A higher rate or ruminal starch digestion for barley than for corn has resulted in greater efficiency of microbial synthesis for dairy ruminants (McCarty et al., 1989) but detailed knowledge about their effects on methane production is yet to be ascertained. Since obviously the diet composition prepared from different cereal sources (corn, barley) may have influences on enteric methane emission it is necessary that different cereal are included in diets for determining total CO2 and CH4 emissions.
The aim of the present study was to evaluate the effect of different cereal sources in choice feeding systems as a tool to mitigate CO2 and enteric emissions of CH4 in Awassi dairy ewes.
MATERIALS AND METHODS
Animal Management and Experimental Diets
The study was carried out at the Sanliurfa Harran University in the Southeast Anatolian Region of Turkey, which lies on longitude 38° 46’E and latitude 37° 08’N. The mean temperature and relative humidity for the experimental period (from the end of May through June, 2008) were 32.12 °C and 25.3%. The experiment was conducted with a total of 16 Awassi dairy ewes in their second and third lactation, each with a single lamb. The ewes were allocated after weaning to two experimental groups, with eight replicates comprising one ewe in a 2.25 m2 (1.5x1.5 m) pen. This gave, two treatments as corn based (CFC) and barley based (BFC) free choice groups. The groups received corn and barley besides wheat bran, cotton seed meal (CSM) and soybean meal (SBM) and alfalfa hay. The ewes were assigned to each treatment according to their live weight and milk yield at the beginning of the experiment. Each pen contained a 15 L bucket for fresh water, which was available during the entire experimental period which included acclimatization/adjustment and feeding trial duration. The feed fed by free choice was divided into six equal parts so that the feed ingredients were presented separately, but ad libitum. The experiment, including a two-week preliminary period, lasted for a total of four weeks. During the second week of the preliminary period, the ewes of the free choice feeding groups were given the opportunity to feed on the ingredients that were subsequently given as choices. During this adaptation period, alfalfa hay was available at all times and the other ingredients were offered alternately every six days for a period of 12 days. Each feed ingredient, except alfalfa hay, was mixed with 1.33% limestone, 1% salt and 0.11% vitamin-mineral mixture in order to ensure adequate intake of micronutrients and also to prevent any possible effect of micronutrients on selection of feed ingredients.
Methane and Carbondioxide Measurement
Respiration Chamber
During the second 14 days of the data collection period, the ewes were taken into one chamber one by one. In each experiment, all of the animals in each treatment group were used and CH4 measurements were made using a ventilated respiration chamber. The respiration chamber formed a closed system in which the fresh intake airflow, based on a continuous system, was controlled. The calculation of the volume of methane emissions was based on the concentration measured in the airflows into and out of the chamber (McGinn et al., 2004). The chamber was 2.5 m wide x 3.9 m tall x 3.7 m deep. The walls and windows of the chamber were insulated to prevent risk of leakage. During the experiment, no significant difference was seen between the outside and inside barometric pressure, which would have encouraged leakage of gas (941-952 mb). Ventilation of the chamber was provided by means of individual fresh air intake and chamber exhaust ducts. The ducts were 10 cm diameter and 30 cm long. Fresh intake and exhaust air (90 m3 h-1) was fed directly into the duct, which contained a pair of fans of the same turning velocity (31.8 m sec-1). The chamber was equipped with a dedicated infrared gas analyzer to detect CH4 and CO2. The ewes were left with their feed trough in the chamber for 23 h, from early morning (08:30) until the following day (07:30) in order to measure differences between the emissions during day and night. The concentration (ppm, on a volume basis) of CH4 in the intake and exhaust ducts of chamber was measured by pumping a sample of the air stream in each duct through the infrared gas analyzer. The level of CH4 and CO2 emission generated from the chamber was calculated via data loggers for each 3 min period from the concentrations in the fresh air intake and the chamber exhaust (Ci and Ce, respectively; ppm).
where, FCO2, FCH4, (g sec-1) are emissions for each chamber, Ve and Vi are the mean air velocity (m sec-1) in the fresh air and the chamber exhaust, MW is the molecular weight of CH4 (16 g mol-1) or CO2 (14 g mol-1), P is barometric pressure (Pa), R is the universal gas constant (8.31 J mol-1 deg K-1), T is the temperature of the air stream (°K) and A is the cross-sectional area of the duct (0.0250 m2).
On the following day the ewes were removed from the chamber and the air inside the chamber was ventilated for 1 h. Manure and urine were removed and the chamber floor was prepared for new gas measurements the following day. During the experiment, the temperature and barometric pressure inside and outside the chamber were recorded for each 3 min period by data loggers. The concentrations of CH4 and CO2 were calculated after adjustment for the temperature, humidity and barometric pressure. The values shown also include emissions of gas from the manure.
Ruminal Fermentation Measurement
Ruminal pH was measured once for two animals per group 30 days after the experiment, in order to prevent changes in their feeding behavior and diet selection caused by stress. A rubber tube was inserted into the rumen via the esophagus and a sample of the ruminal contents (250 mL) were removed using a manual pump. The pH was measured immediately using a pH meter. The ruminal contents were filtered through four layers of cheesecloth. After filtration, the filtrate was centrifuged at 3000 x g for 15 min according to the method of Erwin et al. (1961). Five milliliters of the filtrate was combined with 1 mL of 25% (wt/vol) metaphosphoric acid and stored frozen (-25 °C) until VFA analysis.
Chemical and Statistical Analysis
The compositions of the feed ingredients were determined by the method of AOAC (1998). The neutral detergent fiber (NDF) and acid detergent fiber (ADF) were analyzed based on the method of Van Soest et al. (1991) using an ANKOM fiber analyzer. The metabolizable energy content of the diets was calculated based on the tabulated values published by NRC (1985). Live weight change, milk yield and feed intake were determined every two weeks. The animals were milked by hand during the morning and milk samples were analyzed for milk fat, dry matter, ash, milk protein, non protein nitrogen (NPN) and casein nitrogen using a FOSS MilcoScan FT120 (Denmark). The rest of the nitrogen fractions were determined by calculation. Data were analyzed using a t-test for independent samples in the Compare Means procedure of SPSS version 15.0 (SPSS, 2006). An independent sample t-test was also carried out to determine the significance of the effect of the treatments on the cumulative fluxes of CO2 and CH4.
RESULTS
The components of the ration selected by the ewes are shown in Table 1 for the total experimental period because there were no significant differences between the first and the second half of the experiment.
The results showed that diet selection, with respect to consumption of feed ingredients and nutrient intake was different for the CFC and the BFC rations. The ewes receiving CFC feed ingredients selected a ration that contained similar protein source and more wheat bran (CFC: 15.77; BFC: 1.79%) compared BFC group (p < 0.05). The differences for CSM and SBM were not statistically significant (p > 0.05). The different composition of the ration in the two groups didn’t affect the nutrient composition of the diet. The CFC group selected a ration with a higher content of ether extract (4.26 vs. 3.06). However, there were no differences between the two groups in the energy, protein, ADF, NDF and crude fiber content of the diet.
| Table 1: | Compositions of the diets selected by ewes offered different grain, protein sources (DM basis) |
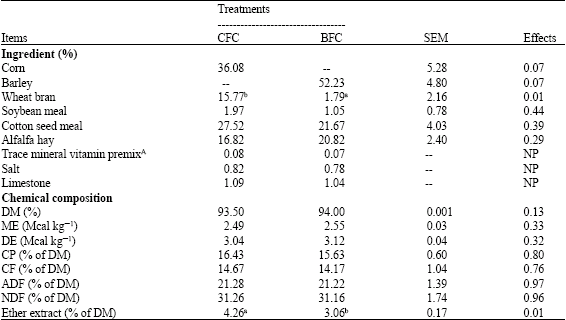 | |
| A: Each kg vitamin-mineral premix provides vit. A 8.000.000 IU; vit.D3 1.000.000 IU; vit E 30.000 mg; Mn 50.000 mg; Fe 50.000 mg; Cu 10.000 mg; Co 150 mg; I 800 mg and Se 150 mg; a-b: Denotes the difference among diets selected by in different groups in the same period (p < 0.05), *: There is a significant difference between two groups (p < 0.05), NS: Not statistical significant (p > 0.05), NP: Not performed statistical analysis | |
| Table 2: | Dry matter and nutrient intakes, milk yield and composition of Awassi ewes fed by different feeding methods |
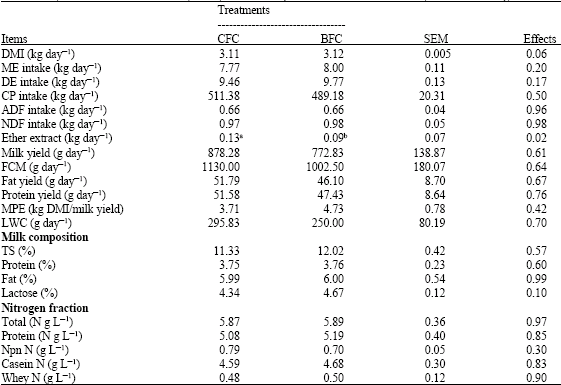 | |
| DMI: Dry matter intake NS: p > 0.05, SEM: Standard error mean, a,b: Means having the different superscript in the same row are significantly different (p < 0.05). ME: Metabolizable energy, DE: Digestible energy, CP: Crude protein, FCM: 4% fat corrected milk, MPE: Milk production efficiency, LWC: Live weight change, TS: Total solid, NPN: Nonprotein nitrogen | |
| Table 3: | Dry matter intake, CO2, CH4 emissions and ruminal fermentation variables for Awassi ewes fed by different feeding methods |
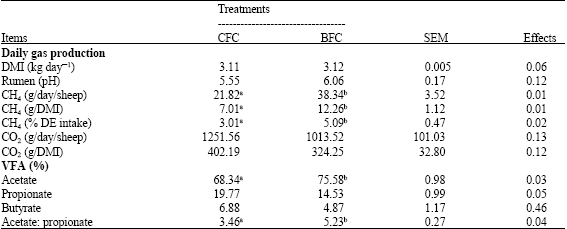 | |
| DMI: Dry matter intake, NS: p > 0.05, VFA: Volatile fatty acids, SEM: Standart error mean, a,b: Means having the different superscript in the same row are significantly different (p < 0.05) | |
The ewes in the CFC group consumed similar dry matter, metabolizable energy, digestible energy, crude protein, ADF and NDF (p > 0.05) but tended to consume more crude fat than the ewes in the BFC group (Table 2, p < 0.05).
The feeding method had no significant effects on milk yield, 4% fat corrected milk yield or milk components (p > 0.05). Ruminal pH values were not affected by the treatments (Table 3). Although there were significant differences in the proportion of acetate, propionate and acetate: propionate ratio between the treatments, the proportion of butyrate was not changed (p > 0.05). Consequently, the acetate:propionate ratio averaged 3.46 for the CFC treatment compared with 5.23 for the BFC treatment (p > 0.05). The CO2 emissions per animal were not significantly different, but the emission of CH4 values per animal, per kg dry matter and per digestible energy intake decreased significantly in the CFC group.
DISCUSSION
The ewes in the BFC group selected a diet containing lower wheat bran but tended to select a diet that contained a higher cereal source (barley) ration than that received corn as cereal source by the CFC group. However there were no significant differences between groups in the energy, ADF and NDF of the diets except for ether extract. The selection of high wheat bran in CFC group could be a reflection of the regulation constant NDF ratio of diet with respect to alternative NDF source such as wheat bran preferences. The results also revealed that the ewes having free access different feed ingredients in two groups made the diets containing similar ratio, energy, protein, NDF and ADF content. In the present study as the ewes received different grain (corn, barley) with alternative protein source (CSM, SBM), they could have a chance to create better nutrient synchronization to rumen microorganism and the host animal. It is well known that ruminants having free access to multiple choices could make their diets to meet nutrient requirements (Fedele et al., 2002; Gorgulu et al., 2003; Provenza, 1995; Yurtseven and Görgülü, 2004, 2007). Although SBM and CSM have similar contents of rumen undegradable protein, when SBM was presented as a choice with CSM, the ewes consumed almost 6.6 times more CSM than SBM. This is probably a reflection of the high protein content of SBM (51.94% CP) and the ewes could meet their protein requirements by consuming less SBM than CSM (21.4% CP). The high protein content of SBM may have also induced negative post-ingestive feedback based on high protein (Provenza, 1995) and thus it was consumed less by ewes in two groups. The less preference for SBM with respect to CSM in both groups could also be attributed to its dusty texture and high protein content supplying more amino acids (especially lysine) to small intestine with less protein with similar ruminal degradability to CSM (NRC, 2001). The diets consumed by the ewes in both groups contained less alfalfa hay than optimal concentrate:forage ratio (60:40) of dairy ewes, thus their diet had lower bulk and higher dry matter content than the classical dairy diet. The changes in diet preferences in groups according to the present study could be also a reflection of the changes in the environmental conditions and the feeding behavior of the sheep in time. The ambient temperature and relative humidity were higher (34.4 vs 21.7 °C and 29.1 vs 23.4%) in the second half of the experiment. It is well documented that ruminants increase their intake of concentrate and reduce their intake of roughage to minimize the heat increment in their bodies under high environmental temperature, when concentrate and roughage are available as free choices (Fedele et al., 2002; Gorgulu et al., 2008; Yurtseven and Gorgulu, 2004, 2007). This could be explained by the changes in feeding behavior (the order of consumption of feed ingredients, the interval between meals, the meal size and meal length, etc.) of the ewes that had free access to feed ingredients (Abijaoude et al., 2000; Gorgulu et al., 2003). In this experiment no metabolic disorder derived from feeding of a high concentrate ration was seen in both groups during the experiment. Small ruminants have a lower capacity to digest forage than larger animals. Their gastrointestinal size and capacity may also limit the intake and utilization of diet with higher forage content.
Depending on the cereal source and carbohydrate source fed, these data indicated no difference in milk yield or milk composition (Table 2). This agrees with some researches (Hadjipanayiotou, 2004) but not with others who observed an increase (Casper and Schingoethe, 1989) or decrease (Poore et al., 1993) in milk yield of dairy cows. There was no difference in milk N fraction especially NPN content of milk between the CFC and BFC groups. It was revealed that the ewes in both groups selected a synchronized diet from among the different cereal and nitrogen sources. Gorgulu et al. (2008) reported that does given free access only to SBM with barley produced higher milk NPN, although all does had similar protein intake. This probably resulted from inefficient use of ammonia nitrogen released from the highly degradable soybean protein used in their experiment. However, in the present study, the sheep used alternative protein sources such as soybean meal and cotton seed meal to provide a synchronized diet (Table 2).
The reductions in CH4 were high on the basis of an individual animal in CFC and were statistically significant. The decrease in energy lost as CH4 was also significant and was the result of selected cereal source in the CFC group compared with the BFC group. Cereal selection that leads to substantial reductions in CH4 emissions may maintain an optimal rumen ecosystem. In present study, the corn as selected cereal source in CFC system had a significant effect on the ruminal content of VFA and clearly decreased the acetate:propionate ratio. The reduction in acetate concentrations as a result of substituting corn for barley is in agreement with the work of McCarty et al. (1989). The effect of the increasing proportion of concentrates in diet on ruminal VFA concentrations is well documented, with an increase in the proportion of propionate. This would be expected to impact on methane production despite the ewes in the CFC group consumed similar amounts of cereal source. Although barley starch may be more rapidly fermented in the rumen than is corn starch, the latter contains greater concentrations of nonstructural carbohydrates (Casper et al., 1999). Casper et al. (1990) also observed an increase of 9.4% in propionate concentration when barley replaced corn in diet. For this reason, the differences between the two groups are also attributed to the differences in starch of two cereals due to more starch content in CFC compared with BFC diet. Moe and Tyrell (1979) found fermentation of soluble carbohydrate to be less methanogenic than that of cell wall carbohydrates. Although ruminal fiber digestion was not measured directly in this study, other variables (for example the acetate:propionate ratio) indicated a considerable reduction in ruminal fiber digestion. In this study, the fat content of the CFC diet was increased by utilization of corn (Table 1). Several studies have reported that the effects of dietary fat content on decreasing of total gas emission are demonstrated (Jenkins et al., 1989; Toral et al., 2009; Wettstein et al., 2000).
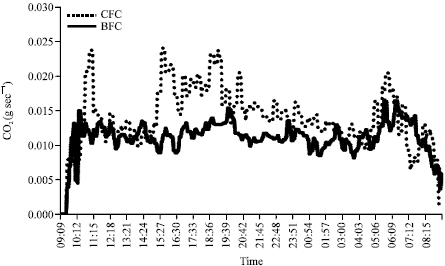
These data suggest that the CFC feeding system had a potential mitigating effect on enteric methane emissions in dairy ewes. However the values of daily methane emission showed a similar trend in both groups with time and followed the feeding and digestive processes. For example, after each meal the emissions of CO2 and CH4 increased sharply and then slowly decreased overnight to the lowest values at 08:30 h, just before the morning feed (Fig. 1).
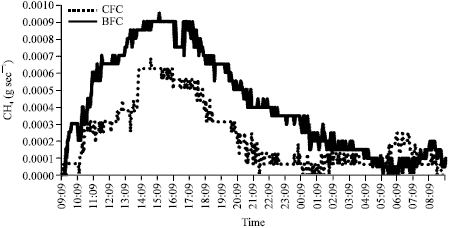 | |
| Fig. 1: | Diurnal CH4 (g sec-1) emission profiles from ewes receiving CFC and BFC diets recorded over experimental period |
 | |
| Fig. 2: | Diurnal CO2 (g sec-1) emission profiles from ewes receiving CFC and BFC diets recorded over experimental period |
Although emission of CH4 reflects the fermentation activity of the digestive tract, the output of CO2 is primarily an indicator of the respiratory activity of the ewes. For this reason, in the experiment, the tendency of CH4 emission was higher than that of CO2 (Fig. 2). A similar diurnal pattern of gas emission was observed by Kirchgessner et al. (1991). In this trial also, CH4 emissions increased after feeding and then declined until the next fresh feed was distributed.
Carbondioxide production is a measurement of heat production, activity and energy expenditure. Di Marco et al. (2001) reported that biting rate was positively correlated CO2 emission rates. The fact that in both groups (CFC, BFC), ewes under choice feeding system emitted similar CO2 (g/DMI and g day-1) was due to the similarity in feeding behavior and ration composition (forage: concentrate ratio) selected.
The ecosystem of the rumen microbial flora was not investigated in this study because the experiment focused especially on the measurement of methane emissions. However, it is well known that cellulolytic microbes tend to diminish in the presence of competitive substrates such as starch. The results showed that there was an important relationship between ruminal VFA and ruminal production of methane. McGinn et al. (2004) reported that adding sunflower oil to the diet clearly decreased ruminal fermentability of the fiber, as evidenced by lower acetate concentration, higher propionate concentration and a lower acetate:propionate ratio. In general, cellulolytic and methanogenic organisms are less tolerant of such changes and may decrease in number (Slyter, 1976). The methanogenic bacteria are the most sensitive to changes in the rumen environment and are affected by many dietary factors. Given that the methanogens are the principal utilizers of hydrogen, their welfare affects ruminal and enteric metabolism and carbon balance. Increased rumen turnover may also restrict methanogens through competition for sufficient generation time. Reduction of methanogenesis tends to promote hydrogen production and shifts the carbon balance toward propionate (Van Soest, 1994).
The results of this study are important and indicate that CFC systems have the potential to decrease methane production in the rumen. Results also indicate that corn selected as cereal source in this experiment modified ruminal fermentation. The factors that influence methanogenic bacteria include those that create a less favorable rumen environment for methanogenesis through an increased rate of passage and rate of digestion, depression of rumination and depression of rumen pH. In the CFC system, these conditions may favor propionate-producing bacteria over acetate producers, making less hydrogen available to methanogens (Van Soest, 1994), as seen in this experiment. Careful selection of different cereal source may allow the manipulation of methane production in the rumen.
REFERENCES
- Abijaoude, J.A., P. Morand-Fehr, J. Tesier, P.H. Schimidely and D. Sauvant, 2000. Diet effect on the daily feeding behaviour, frequency and characteristics of meals in dairy goats. Livest. Prod. Sci., 64: 29-37.
CrossRef - Casper, D.P. and D.J. Schingoethe, 1989. Lactational response of dairy cow to diets varying in ruminant solubilities of carbonhydrate and crude protein. J. Dairy Sci., 68: 928-941.
PubMed - Casper, D.P., H.A. Maiga, M.J. Brouk and D.J. Schingoethe, 1999. Synchronization of carbonhyrate and protein sources on fermentation and passage rates in dairy cows. J. Dairy Sci., 82: 1779-1790.
PubMed - Casper, D.P., D.J. Schingoethe and W.A. Eisenbeisz, 1990. Response of early lactation dairy cows fed diets varying in source of non structural carbohydrate and crude protein. J. Dairy Sci., 73: 1039-1050.
PubMed - Di Marco, O.N. and M.S. Aello, 2001. Energy expenditure due to the forage intake and walking of grazing cattle. Arq. Bras. Med. Vet. Zootec., 53: 105-110.
CrossRef - Fedele, V., S. Claps, R. Rubino, M. Calandrelli and A.M. Pilla, 2002. Effect of free choice and traditional feeding systems on goat feeding behaviour and intake. Livest. Prod. Sci., 74: 19-31.
CrossRef - Gorgulu, M., S. Yurtseven, H.R. Kutlu and U. Serbester, 2008. Effects of grain and protein sources on diet preferences, milk yield and milk composition of choice-fed german fawn x hair crossbred goats in mid lactation. J. Anim. Vet. Adv., 7: 1241-1251.
CrossRefDirect Link - Gorgulu, M., O. Guney, O. Torun O. Ozuyanyk and H.R. Kutlu, 2003. An alternative feeding system for dairy goats: Effect of free choice feeding on milk yield and milk composition in early lactation of Damascus goats. J. Anim. Feed Sci., 12: 33-44.
Direct Link - Hadjipanayiotou, M., 2004. Replacement of barley grain for corn in concentrate diets fed to dairy Damascus goats at different frequencies. Small Rumin. Res., 51: 229-233.
CrossRef - Johnson, K.A. and D.E. Johnson, 1995. Methane emissions from cattle. J. Anim. Sci., 73: 2483-2492.
PubMedDirect Link - McCarty, R.D., T.H. Klusmeyer, Jr., J.L. Vicini, J.H. Clark and D.R. Nelson, 1989. Effect of source of protein and carbonhydrate on ruminal fermentation and passage of nutrients to the small intestine of lactating cows. J. Dairy Sci., 72: 2002-2016.
PubMed - McGinn, S.M., K.A. Beauchemin, T. Coates and D. Colombatto, 2004. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast and fumaric acid. J. Anim. Sci., 82: 3346-3356.
PubMedDirect Link - Moe, P.W. and H.F. Tyrrell, 1979. Methane production in dairy cows. J. Dairy Sci., 62: 1583-1586.
CrossRefDirect Link - NRC, 2001. Nutrient Requirements of Dairy Cattle. 7th Edn., National Academy Press, Washington, DC, ISBN: 978-0-309-06997-7, Pages: 405.
CrossRefDirect Link - Poore, M.H., J.A. Moore, R.S. Swingle, T.P. Erc and W.H. Brown, 1993. Response of lactating Holstein cows to fibre source and ruminal starch degradability. J. Dairy Sci., 76: 2235-2243.
Direct Link - Provenza, F.D., 1995. Postingestive feedback as an elementary determinant of food preference and intake in ruminants. J. Range Manage., 48: 2-17.
Direct Link - Santini, F.J., C.D. Lu, M.J. Potchoiba and J.M. Fernandez, 1992. Dietary fiber and milk yield, mastication, digestion and rate of passage in goats fed alfalfa hay. J. Dairy Sci., 75: 209-219.
PubMed - Schimidely, P., M. Lloret Pujol, P. Bas, A. Rouzeau and D. Sauvant, 1999. Influence of feed intake and source of dietary carbohydrate on metabolic response to propionate and glucose challenges in lactating goats. J. Dairy Sci., 82: 738-746.
PubMed - van Soest, P.J., J.B. Robertson and B.A. Lewis, 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci., 74: 3583-3597.
CrossRefPubMedDirect Link - Wang, C.J., S.P. Wang and H. Zhou, 2009. Influences of flavomycin, ropadiar, and saponin on nutrient digestibility, rumen fermentation and methane emission from sheep. Anim. Feed Sci. Technol., 148: 157-166.
CrossRefDirect Link - Yurtseven, S. and M. Gorgulu, 2004. Effects of grain sources and feeding methods, free-choice vs. total mixed ration, on milk yield and composition of German Fawn x Hair crossbred goats in mid lactation. J. Anim. Feed Sci., 13: 417-428.
Direct Link - Jenkins, T.C., T. Gimenez and D.L. Cross, 1989. Influence of phospholipids on ruminal fermentation in vitro and on nutrient digestion and serum lipids in sheep. J. Anim. Sci., 67: 529-537.
PubMed - Toral, P.G., A. Belenguer, P. Frutos and G. Hervas, 2009. Effect of the supplementation of a high-concentrate diet with sunflower and fish oils on ruminal fermentation in sheep. Small Rumin. Res., (In Press).
CrossRef