
P. Srinivasan
Department of Biotechnology, Life Sciences Building, University of Madras, Guindy Campus, Chennai-600 025, Tamil Nadu, India
P. Ramasamy
Department of Biotechnology, Life Sciences Building, University of Madras, Guindy Campus, Chennai-600 025, Tamil Nadu, India
G. P. Brennan
School of Biology and Biochemistry, Medical Biology Centre, The Queens University of Belfast, Belfast, BT9 7BL, Northern Ireland, UK
R.E.B. Hanna
Pathology Department, Veterinary Sciences Division, Agri-Food and Biosciences, Stormont, Belfast Bt43SD, Northern Ireland, UK
Asian Journal of Animal and Veterinary Advances
Year: 2007 | Volume: 2 | Issue: 4 | Page No.: 166-183







ABSTRACT
Bacteriophages of Vibrio harveyi from a shrimp farm located at Tuticorin, Tamilnadu, India, were isolated, characterized and identified. The phages, when examined by electron microscopy, were found to have icosahedral morphology. The phages belong to the Siphoviridae family and were designated VH1 to VH8. They produced circular clear plaques of growth inhibition on Vibrio sp., plate cultures. The growth inhibitory effects of the bacteriophages on isolates of V. harveyi were screened by a one-step agar bioassay method and the results suggest that the infectivity of the V. harveyi phages was not affected by ribonuclease, protease, trypsin, chloroform, acetone, ethyl alcohol, methyl alcohol and Triton-X100 treatment at 37°C. Even after their storage in chloroform or DMSO (dimethyl sulfoxide, 7%) at -40°C. They showed 100% infectivity and plaque-forming ability. In contrast, Infectivity of the V. harveyi phages was affected by lysozyme and SDS and in these cases; plaques were not formed on agar plate assays. The phages were found to be stable over the pH range 6-11 and exhibited optimal survival at 37°C while complete inactivation occurred at temperatures exceeding 55°C. All the isolates of bacteriophage (VH1-VH8) initiated an infection cycle and caused lysis of the host bacterial cells within 2 h from the start of incubation. The propagation curve for each phage shows a burst time started from 1-10 h. The results suggest that bacteriophages of Vibrio sp., might be effectively used in vivo as biological agents to control these pathogenic bacteria in aquaculture systems.
PDF Abstract XML References Citation
How to cite this article
P. Srinivasan, P. Ramasamy, G. P. Brennan and R.E.B. Hanna, 2007. Inhibitory Effects of Bacteriophages on the Growth of Vibrio sp., Pathogens of Shrimp in the Indian Aquaculture Environment. Asian Journal of Animal and Veterinary Advances, 2: 166-183.
DOI: 10.3923/ajava.2007.166.183
URL: https://scialert.net/abstract/?doi=ajava.2007.166.183
DOI: 10.3923/ajava.2007.166.183
URL: https://scialert.net/abstract/?doi=ajava.2007.166.183
INTRODUCTION
Bacteriophages represent a potential alternative to the use of antibiotics in the control of pathogenic bacteria. Bacteriophages have been administered to infected animals and humans in an effort to reduce populations of pathogenic bacteria through the natural ability of the phages to target and destroy bacterial cells. Interest in the ability of phages to control bacterial populations has extended from medical applications into the fields of agriculture, aquaculture and the food industry (Fuhrman and Suttle, 1993; Nakai and Park, 2002; Withey et al., 2005). Viruses of bacteria have been hypothesised to play a key role in controlling host bacterial population density and diversity, nutrient flow in marine food webs and the transfer of genes between hosts (Heldal and Bratbak, 1991; Fuhrman and Suttle, 1993). Due to their antibacterial activities, interest in the possibilities of bacteriophage therapy, as an alternative to antibiotics, is re-emerging (d’Herelle, 1917). Bacteriophage-typing schemes have been developed for most groups of pathogenic bacteria and more recently their host specificity has been applied in bacterial detection and diagnostic strategies (Marks and Sharp, 2000).
The present study was undertaken to investigate the antibacterial potential of bacteriophages isolated from cultures of Vibrio spp. found in shrimp aquaculture systems and in the marine environment of South India. This information may help in the development of specific phage therapy applicable to local aquaculture systems, which is considered to represent an interesting emerging alternative to antibiotic therapy and vaccination.
MATERIALS AND METHODS
Bacteria, Phages, Media and Culture Methods
V. harveyi isolates were obtained from shrimp farms located along the coast of the Bay of Bengal at Thulukenkulam (30 kms from Tuticorin, 08°40’ 05 to 08° 48’05 N; 78° 10’ 05 to78° 14’10 E), Chennai (Tamil Nadu) and Nellore (Andhra Pradesh), India (Fig. 1) during 2001-2002 and grown on TCBS (Thio-sulphate Citrate Bile Salts) agar. The bacteria were cultured in LB liquid media for 24-48 h at 37°C and the cultures were then centrifuged at 10000 rpm for 5 minutes to sediment the bacterial cells. The supernatant was decanted and saved. Four drops of chloroform were added to the supernatant to kill any residual bacteria.
Phage Isolation and Purification
V. harveyi phages (3x103 plaque forming units/mL) were mixed with an equal volume of overnight Vibrio bacterial cell culture and incubated at room temperature for 30 min to allow the phage particles to adsorb to the bacteria. The incubated sample was mixed with pre-cooled (45°C) 0.6% Luria Bertaini top agar and evenly poured over pre-set 1.5% (w/v) Luria Bertaini agar. The top agar mixture was allowed to set for 5-15 min at bench temperature (25°C) and the plate was incubated at 37°C for 24-48 h.
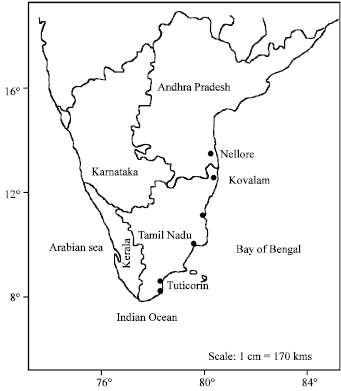 | |
| Fig. 1: | South India map showing location of (collection sites) shrimp farms and hatcheries |
When confluent lysis occurred, bacteriophages were harvested by adding 5 ml of SM solution [NaCl (5.8 g), MgSO4 (2 g), 1 M Tris.Cl (pH 7.5 - 50 mL), 2% (w/v) Gelatin solution (5 mL) and Milli Q water (made up to 1000 mL)] and leaving the plate at room temperature (25°C) for 30 min with gentle shaking. This allowed the phages to diffuse out of the agar and the resulting suspension of phages was termed the ‘lysate’. The lysate was removed from the culture plate and treated with chloroform at ratio [3 Lysate: 1 chloroform] for 30 min at room temperature. It was then centrifuged at 4°C for 15 min at 5000 rpm to remove the pellet containing unlysed, uninfected and infected bacterial cells and the supernatant containing phages was recovered. The supernatant was further filtered through a 0.22 -μm pore-size syringe filter to remove any remaining unlysed, bacterial cells and the filtrate contained only phages and thus the phage purification was carried out repeatedly by following the method of Moebus and Nattkember (1981) to ensure the purity of the phage stock. The phage suspension was stored at -20°C prior to further use.
Long-term Storage of V. harveyi Bacteriophage
The following procedure was used for long-term storage of Vibrio bacteriophages in the archival collections. The phage isolate was treated with 7% DMSO (Dimethyl sulfoxide) (v/v) or with an equal volume of chloroform and frozen at -40°C (30 days). The stocks of each phage isolate were sub-divided into 100 mL prior to storage. For use, the phage stocks were removed from storage and thawed. The remaining stock if any was recapped and returned immediately to -40°C. The contents of tubes containing aliquots of phages were mixed with the appropriate host bacterial cell isolates and streaked on to Nutrient/Luria Bertaini agar medium Petri dishes and in each case a single phage plaque was isolated and used subsequently.
Infectivity and Inhibitory Effects of Bacteriophages on the growth of Isolates of Vibrio sp.
The agar bioassay method and one step growth experiments were carried out to determine the infectivity and inhibitory effect of Vibrio bacteriophages.
Agar Bioassay
Inhibitory effects of eight bacteriophages of V. harveyi were determined against 40 different isolates of Vibrio sp., Sterilised Nutrient/Luria Bertaini agar medium was poured into Petri dishes and after solidification, two 3 mm diameter wells were punched in each plate with a gel puncher. One hundred μL of target Vibrio cell culture was spread on each agar plate and 100 μL aliquots of phages was pipetted into one of the wells. The remaining empty well was inoculated with sterile Nutrient broth/Luria Bertaini broth alone (without phages) to serve as a negative control. A zone of clearance around the well indicated phage infectivity and lysis of the cells of Vibrio sp.
One-step Bacteriophage Growth Experiment
The one-step bacteriophage growth experiment was developed by Ellis and Delbruck (1939) and was later described by Adams (1959). It was used to determine the lytic burst size and burst time for Vibrio bacteriophages. One ml of cells of the Vibrio isolate in LB broth was mixed with 1 ml of Vibrio bacteriophage suspension (containing 3x103 plaque-forming units). Optical density readings of the mixture of bacteriophage and Vibrio host cells were taken every 1 h up to 10 h at 600 nm (OD600) using a UV1601 visible spectrophotometer (Shimadzu corporation, Japan).
Electron Microscopy
A suspension of V. harveyi phage infected V. damsela was centrifuged at 5000 rpm for 30 min and the resulting pellet was fixed with 2% EM grade glutaraldehyde buffered with 0.1 M sodium cacodylate containing 3% sodium chloride and 5% sucrose. After 24 h fixation, the pellet of V. harveyi phage infected V. damsela was washed repeatedly with sodium cacodylate buffer, post fixed with 0.1% OsO4 for 1 h at 4°C, dehydrated through a graded ethanol series (from 30% ethanol with 30 min. in each change) and embedded in Epon 812. Sixty to ninety-nanometer (i.e., gold-and silver-colored) sections were cut with a diamond or glass knife using a reichert Ultracut E ultramicrotome, stained with aqueous uranyl acetate and concentrated lead citrate and viewed using a Philips E10 JEOL 100B transmission electron microscope at an accelerating voltage of 80 kV. Electron micrographs of random bacterial cell clusters were taken.
Effects of Enzymes, Organic Solvents and Detergents on Infectivity, Survival and Multiplication of V. harveyi Bacteriophages
Purified Vibrio bacteriophage lysate samples (3x103 PFU mL-1) were treated with enzymes (RNAse, protease, lysozyme and trypsin at a final concentration each of 100 μg mL-1) organic solvents (chloroform, acetone, ethyl alcohol and methyl alcohol at a final concentration each of 50%) and detergents (SDS and Triton-X 100 at a final concentration each of 50%) and were incubated at 37°C for 1 h. A purified Vibrio bacteriophage lysate sample incubated without enzymes, organic solvents or detergents served as control. The contents of tubes containing aliquots of treated and untreated phages were mixed with the appropriate host bacterial cell isolates and streaked on to Nutrient/Luria Bertaini agar medium. The samples were incubated for 16-24 h at 37°C and the number of plaques formed on a lawn of Vibrio cells in the treated and untreated samples were recorded and compared.
Effect of pH on the Stability and Infectivity of V. harveyi Phages
Growth media (LB broth/Nutrient broth) with a range of different pH values (4-12) were prepared using 5 N NaOH or 1 N HCl and were used to test their effect on the stability and infectivity of V. harveyi phages. Reaction mixtures incubated without the V. harveyi bacteriophage served as control. Samples of V. harveyi phage lysate were used to infect uninfected cells of Vibrio sp., by mixing and incubating for 1 h and subsequently the suspension of phage adsorbed/infected cells was spread on the agar plates and incubated for 16-24 h at 37°C and the plaques formed were counted and compared.
Effect of Temperature on the Survival and Infectivity of V. harveyi Phages
Samples of V. harveyi phage lysate (3x103 PFU mL-1) were incubated at temperatures ranging from 20°C to 90°C for 1 h and were used to test their effect on the stability and infectivity of Vibrio phages. Samples of treated and untreated Vibrio phage lysate were used to infect cells of Vibrio sp., by mixing and incubating them for 1 h and subsequently the suspension of phage adsorbed/infected cells was spread on the agar plates and incubated for 16-24 h at 37°C and the plaques formed were counted and compared.
RESULTS
Detection of V. harveyi Bacteriophages
Phages of V. harveyi designated as VH1-VH8 were isolated from aquaculture environment in India. They produced circular plaques on isolates of V. harveyi, V. parahaemolyticus, V. anguillarum and V. mimicus. The plaques formed were about 1.9 mm in diameter after 24 h of incubation at 37°C. Isolates of phages of V. harveyi were collected from P. monodon egg sample, Artemia nauplii, Artemia nauplii rearing tank water; MBV-infected post-larval P. monodon, source sea water and from WSSV-infected shrimp tissues viz., hepatopancreas, intestine and gills of P. monodon (Table 1 and Fig. 2).
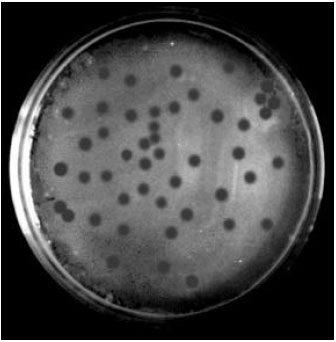 | |
| Fig. 2: | Photographs showing clear circular plaques (1.9 mm) produced by bacteriophages of Vibrio harveyi |
| Table 1: | Sources and total number of Vibrio sp., isolates recovered from the aquaculture environments |
 | |
Long-term Storage of V. harveyi Bacteriophages
The plaque-forming experiments revealed that the ability of infectivity of V. harveyi bacteriophages was not affected even after their storage in chloroform or DMSO (dimethyl sulfoxide, 7%) for 30 days at -40°C. All the stored samples of bacteriophages in chloroform or DMSO (dimethyl sulfoxide, 7%) for 30 days at -40°C showed 100% infectivity and plaque-forming ability.
Growth Inhibitory Effects of V. harveyi Bacteriophages on Isolates of Vibrio spp. Determined by the Agar Bioassay Method
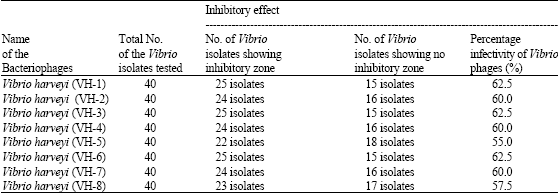
All the isolates of bacteriophages were found to inhibit the growth of several different isolates of Vibrio spp. when they were assayed in LB/Nutrient agar medium. The inhibition of growth of Vibrio isolates by V. harveyi bacteriophages was found to be a maximum at a temperature of 37°C. Three of the V. harveyi phage isolates namely VH-1, VH-3 and VH-6 showed 62.5% of inhibition of growth of Vibrio sp., while the other bacteriophage isolates showed inhibition of growth in the order VH-2, VH-4 and VH-7 (60%), VH-8 (57.5%) and VH-5 (55%) (Table 2 and Fig. 3a-d).
One-step Bacteriophage Growth Experiment
The propagation curve for each phage shows a burst time started from 1-10 h. The rate (percentage) of absorption of phages indicated that more than 80% of the bacteriophages adsorbed to the host cells during 10 h incubation. All the isolates of bacteriophage (VH-1-VH-8) initiated an infection cycle and caused lysis of the host bacterial cells within 2 h from the start of incubation. The absorbance (A600) for all the phage-treated experimental samples of the isolates of Vibrio spp. was 0.05 at the beginning and reached a maximum value of 0.168 (VH-1), 0.062 (VH-2), 0.168 (VH-3), 0.76 (VH-4), 0.117 (VH-5), 0.076 (VH-6), 0.07 (VH-7) and 0.201 (VH-8) after 2 h of incubation and then, in all cases, the absorbance declined during the subsequent period, reaching values of 0.125 (VH-1), 0.028 (VH-2), 0.011 (VH-3), 0.024 (VH-4), 0.011 (VH-5), 0.024 (VH-6), 0.008 (VH-7) and 0.12 (VH-8) after 10 h of incubation.
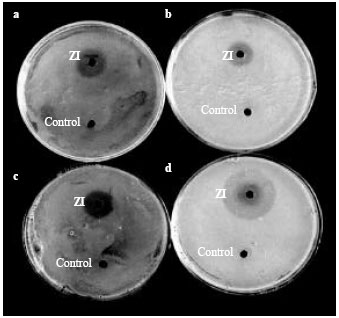 | |
| Fig. 3a-d: | Petriplates consisting of Vibrio isolates exhibiting a zone of inhibition around the well of Vibrio harveyi bacteriophage against (a) Vibrio mimicus, (b) Vibrio furnissi, (c) Vibrio parahaemolyticus and (d) Vibrio damsela. ZI- Zone of Inhibition |
| Table 2: | Infectivity and inhibitory effects of Vibrio bacteriophages on the growth of Vibrio isolates recovered from aquaculture environments using agar bioassay |
 | |
In contrast, the absorbance value of the control samples, incubated in the absence of bacteriophage, was 0.05 at the beginning of incubation and subsequently increased to reach a maximum value of 2.08 (VH-1), 2.0 (VH-2), 2.078 (VH-3), 0.321 (VH-4), 0.458 (VH-5) and (VH-6), 0.278 (VH-7) and 9.1 (VH-8) after 10 h of incubation at pH 7 indicating the proliferation and growth of uninfected bacterial cells (Fig. 4a-h).
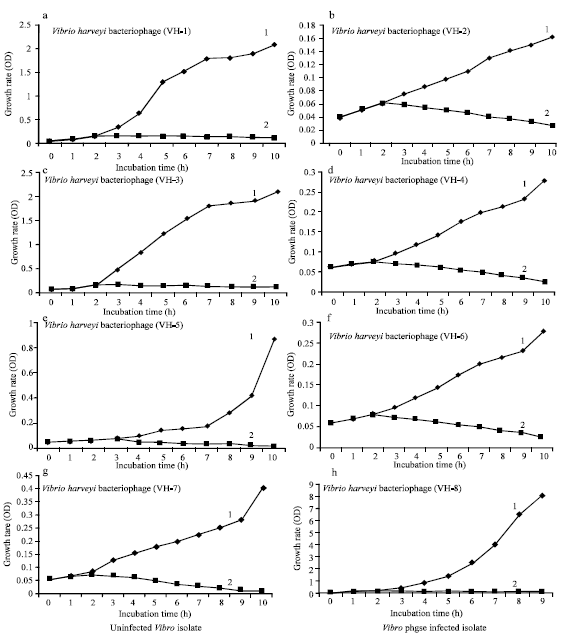 | |
| Fig. 4: | One step growth experiment of Vibrio isolates infected with Vibrio harveyi bacteriophages (VH1-VH8, a-h, respectively) compared with uninfected controls. The incubation time was 0-10 h at 37°C; 1 = Uninfected Vibrio isolate; 2 = Vibrio harveyi phage-infected isolate |
Transmission Electron Microscopy of V. harveyi Bacteriophages
Transmission electron microscopy of bacteriophages and infected bacteria was carried out to determine the occurrence, structure, site and mode of infection of the phages. The eight distinct V. harveyi phage isolates were examined. Phages were commonly encountered on the cell wall of the infected V. harveyi cells examined. Each phage consisted of a double envelope enclosing the nucleic acid and proteins and also featured a tail. The head displayed icosahaedral symmetry and the tail, when visible on the electron micrographs, was seen to be short. Considering the morphological features and the host genus, the phages can be classified as belonging to the order Caudovirales of the family Siphoviridae. Phages from each and every isolate examined (VH-1-8) were found to conform to this description and identity (Fig. 5a-h).
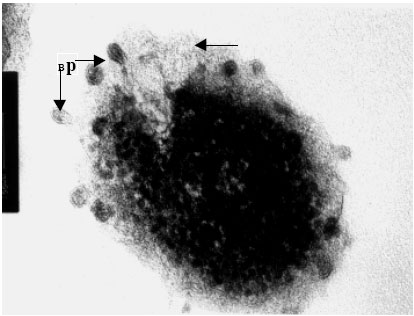 | |
| Fig. 5a: | Transmission electron micrograph showing the surfacing of Vibrio harveyi (VH-1) phage from the cell wall of the host cell bacterium Vibrio sp., Note the presence of partially assembled phage (Bp) particles within the cytoplasm while those completely assembled particles are seen on the ruptured cell surface (unlabelled arrow) |
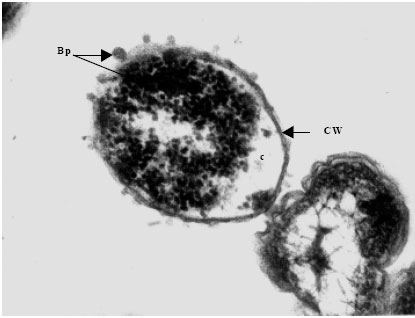 | |
| Fig. 5b: | Transmission electron micrograph showing the surfacing of Vibrio harveyi (VH-2) phages (Bp) from the Cell Wall (CW) of the host cell bacterium Vibrio sp., Note the presence of partially assembled phage particles within the cytoplasm (c) while those completely assembled particles are seen on the cell wall |
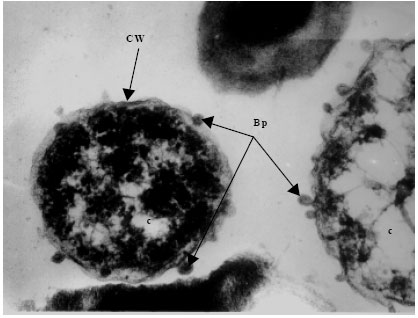 | |
| Fig. 5c: | Transmission electron micrograph showing the surfacing of Vibrio harveyi (VH-3) phages (Bp) from the Cell Wall (CW) of the host cell bacterium Vibrio sp., Note the presence of partially assembled phage particles within the cytoplasm (c) while those completely assembled particles are seen on the cell wall |
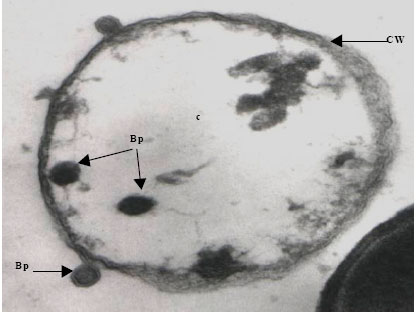 | |
| Fig. 5d: | Transmission electron micrograph showing the surfacing of Vibrio harveyi (VH-4) phages (Bp) from the Cell Wall (CW) of the host cell bacterium Vibrio sp., Note the nature of the lysed cytoplasm (c) |
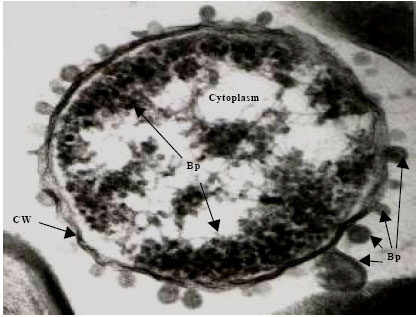 | |
| Fig. 5e: | Transmission electron micrograph showing the surfacing of Vibrio harveyi (VH-5) phages (Bp) from the Cell Wall (CW) of the host cell bacterium Vibrio sp., Note the presence of partially assembled phage particles within the cytoplasm while those completely assembled particles are seen on the cell |
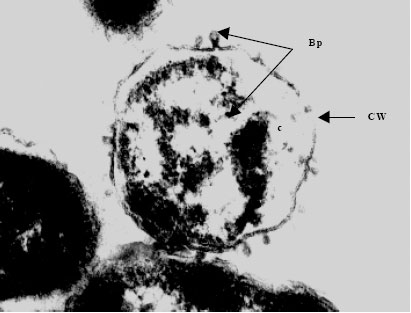 | |
| Fig. 5f: | Transmission electron micrograph showing the surfacing of Vibrio harveyi (VH-6) phages (Bp) from the Cell Wall (CW) of the host cell bacterium Vibrio sp., Note the presence of partially assembled phage particles within the cytoplasm (c) while those completely assembled particles are seen on the cell |
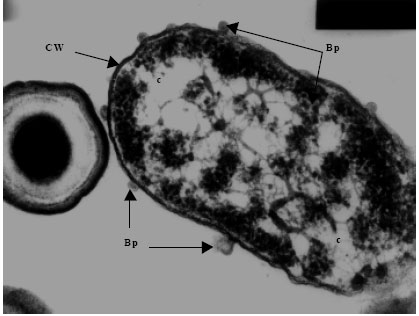 | |
| Fig. 5g: | Transmission electron micrograph showing the surfacing of Vibrio harveyi (VH-7) phages (Bp) from the Cell Wall (CW) of the host cell bacterium Vibrio sp., Note the presence of partially assembled phage particles within the cytoplasm (c) while those completely assembled particles are seen on the cell |
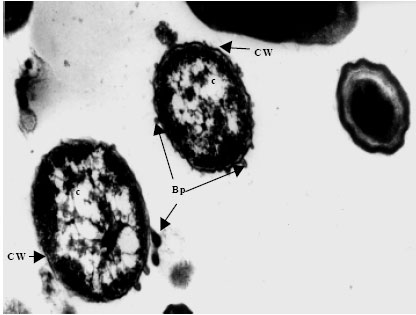 | |
| Fig. 5h: | Transmission electron micrograph showing the surfacing of Vibrio harveyi (VH-8) phage (Bp) from the Cell Wall (CW) of the host cell bacterium Vibrio sp., Note the presence of partially assembled phage particles within the cytoplasm (c) while those completely assembled particles are seen on the cell |
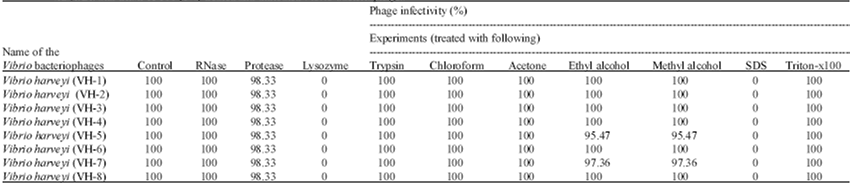
| Table 3: | Effects of enzymes viz., RNase, protease, lysozyme, trypsin (100 μg mL-1); organic solvents viz., chloroform, acetone, ethyl alcohol and methyl alcohol (50%); and detergents viz., SDS and triton-X 100 (1%) treated and untreated Vibrio bacteriophages |
 | |
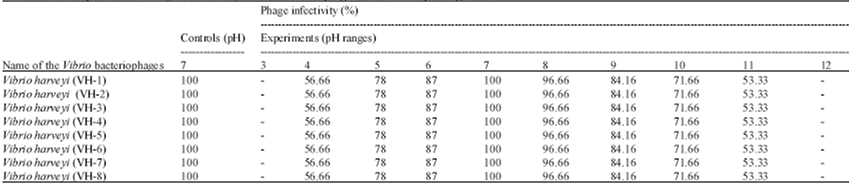
| Table 4: | Effect of pH on the stability and infectivity of Vibrio bacteriophages with different pH ranges 3-12 |
 | |
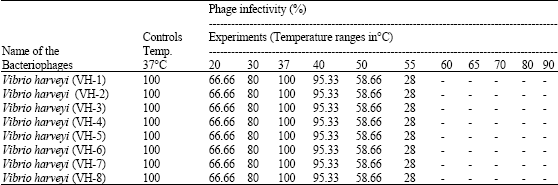
| Table 5: | Effect of temperature on the stability and infectivity of Vibrio bacteriophages at different temperature from 20-90°C |
 | |
The cytoplasm of phage-infected bacterial cells was less electrons dense than that of uninfected cells. Within the cytoplasm of some bacterial cells, various stages in the multiplication and assembly of viral particles were identified. This was apparently associated with ‘clearing’ of the cytoplasm, presumably the result of lytic activity induced by phage infection (Fig. 5d).
Effects of Enzymes, Organic Solvents and Detergents on the Phages of V. harveyi
The effects of enzymes (ribonuclease, protease, lysozyme and trypsin) organic solvents (chloroform, acetone, ethyl alcohol and methyl alcohol) and detergents (SDS and Triton-X 100) on the survival, infectivity, growth and multiplication of the V. harveyi phages (VH1-VH8) were determined. Infectivity of the V. harveyi phages was not affected by ribonuclease, protease, trypsin, chloroform, acetone, ethyl alcohol, methyl alcohol and Triton-X100 treatment at 37°C as determined by the ability of the phages to form plaques similar to those formed by untreated control phages on agar plate assays. Infectivity of the V. harveyi phages was however affected by lysozyme and SDS. In these cases, plaques were not formed on agar plate assays.
The percentage survival and infectivity of phages of V. harveyi (VH1 to VH8) after treatment with ribonuclease, trypsin, chloroform, acetone, ethyl alcohol and methyl alcohol (except VH5 and VH7) and Triton-X 100 were seen to be relatively higher (100%) when compared to treatment with other chemicals and the phages infectivity were found to be complete (100%). In contrast, the percentage infectivity of V. harveyi phages (VH1 to VH8) was observed to be 98.33% after protease treatment. 97.36 and 95.47% infectivity of V. harveyi phages (VH5 and VH7) was observed after ethyl alcohol and methyl alcohol treatment (Table 3).
Effect of pH and Temperature on the Stability and Infectivity of V. harveyi Bacteriophages
V. harveyi bacteriophages were stable and exhibited infectivity over the pH range 4-11, but there was complete loss of bacteriophages infectivity at pH 3 and 12. All the isolates of bacteriophages showed 100% survival and infectivity at pH 7, with uniform reduction of survival and infectivity in the ranges pH 6-4 and pH 8-11 (Table 4). V. harveyi bacteriophages were found to be stable at temperatures in the range 20-50°C but were inactivated at temperatures above 55°C (Table 4 and 5).
DISCUSSION
V. harveyi bacteriophages, designated VH1-VH8, were isolated from diverse components of the aquaculture environment and from various tissues of infected shrimp. Experimental infectivity studies, plaque assays and transmission electron microscopy have conclusively demonstrated the ubiquitous occurrence and distribution of Vibrio-specific phages in Vibrio sp., from the Indian aquaculture environment. This is the first report of marine Vibrio-specific phages from India. A bacteriophage of V. parahaemolyticus isolated from seawater is known to have a broad host range infecting various Vibrio species (Paul et al., 1997; Matsuzaki et al., 2000; Miller et al., 2003) and nine phages of V. vulnificus have been isolated from estuarine waters of Louisiana (Pelon et al., 1995). Four marine phage-host systems have been described from Mamala Bay, Hawaii Jiang et al. (1998). In the present study it was found that phages of V. harveyi formed clear plaques 1.9 mm in diameter in agar assays. Matsuzaki et al. (1998a, b) showed that vibriophage KVP20 exhibited a much narrower host range than KVP40 and formed plaques in only some V. parahaemolyticus strains. Bacteriophage VPP97 infects many strains of V. parahaemolyticus isolated from marine environments and forms clear plaques 0.6-1.0 mm in diameter (Pirisi, 2000). V. alginolyticus phages form plaques 0.5-0.9 mm in diameter (Ju et al., 1997).
Whilst the present account deals specifically with the inhibitory effects of bacteriophages on the growth of shrimp-pathogenic bacteria, other agents that inhibit bacterial growth in the aquaculture environment have been described in the literature. More than 400 bacterial isolates from the intestine and the external surface of farmed Scophtalmus maximus as well as from fish food and hatchery water were screened for inhibitory effects against the fish pathogen V. anguillarum HI 11345 and seven other fish pathogens (Westerdahl et al., 1991). Of the total number of isolates from the intestine, 28% were found to be inhibitory against V. anguillarum H 11345 and 60% had an inhibitory effect on five other fish-pathogenic serotypes of V. anguillarum. Pseudomonas fluorescens AH2 has been reported to reduce the growth of V. anguillarum (Gram et al., 1999). A marine bacterial strain, Pseudomonas I-2 is reported to produce inhibitory compounds against shrimp pathogenic Vibrio spp. in aquaculture systems especially V. harveyi, V. fluvialis, V. parahaemolyticus, V. damsela and V. vulnificus (Chythanya et al., 2001).
The one step growth studies carried out in the current investigation revealed relatively slow lytic activity for most of the vibriophages tested. Wu et al. (1981) demonstrated the formation of AH1 (Aeromonas hydrophila) phage particles after 50 min of infection and bursting occurred after100 min when approximately 160 phages were present in each cell. Phage pA1 (P. aerunginosa) has been shown to exhibit a latent period of 60 min, a rise period of 60 min and a burst size of 130 particles per cell (Hirayama et al., 1993). Jiang et al. (1998) reported that four phages viz., T-φD0, T-φD2S, T-φHSIC and T-φD1B exhibited a latent period ranging from 90 to 180 min. Eisenstark (1967) investigated the duration of the phage replication cycle and the relative amounts of progeny phage released using one-step replication experiments.
In the present study it has been shown that V. harveyi phage isolates were stable in long-term storage and they showed undiminished survival and infectivity even after their long-term storage. Similarly, Catherine et al. (2004) found no decrease in titre over a 6 -month period storage at 4°C and -70°C and phage stability remained unaffected by freeze/thawing. In contrast, Pelon et al. (1995) reported that whilst phage isolates were found to be stable on storage at 4°C they exhibited a rapid loss of infectivity with repeated freezing and thawing at -70°C and similar results were observed by Gantzer et al. (1998).
Transmission electron microscopy revealed the presence of bacteriophages on the surface of infected bacterial cells. Attachment was by means of the short tail structure. Intra-cellular multiplication of the phages and lysis of the host cell following the phage multiplication were also visualized, providing evidence of the mode of infection and multiplication of phage particles in the host cell. These vibriophages may be unique to the Indian aquaculture environment and have not yet been reported elsewhere. Pelon et al. (1995) described nine phage isolates infectious for V. vulnificus. These were detailed as follows: (A) small, tailless icosahedral capsids (S1); (B) icosahedral heads with thin flexible tails (P13, P38, P53, P65, P108 and P111); (C) prolate icosahedral heads with relatively short (130 nm) rigid contractile tails (P68); and (D) icosahedral heads with long (233 nm) rigid tails consisting of contractile sheaths surrounding rigid tubes (P147). Morphological groups of Vibriophages similar to those isolated in the study of Pelon et al. (1995) were also seen in our study. Shaffer et al. (1999) have shown that the morphology of theUNL-1 virion (P. aerunginosa) resembles most closely of that of members of the Myoviridae family, which are large and structurally complex bacterial viruses that possess contractile tails with fibers.
Bacteriophage VPP97 infects strains of V. parahaemolyticus and electron microscopy studies have revealed the presence of a hexagonal head and a relatively long tail (Yoon et al., 1999). A vibriophage, KVP 241 and six of its relatives were isolated independently from seawater using V. parahaemolyticus as the host. Each of these phages has a hexagonal head and a tail with a contractile sheath. The host range is limited to some strains of V. parahaemolyticus (Matsuzaki, et al., 2000). Vibriophages from V. alginolyticus, V. fluvialis and V. furnissii were found to possess a hexagonal head and a relatively long tail (Pirisi, 2000). Ackermann (1996) reported that over 4500 bacterial viruses have been examined by electron microscopy since 1959 and 4400 of these (96%) have been found to be tailed whilst only 162 phages (4%) are cubic, filamentous, or pleomorphic. They are classified by morphotype and host genus. Siphoviridae is one of the families that contains phages with long, non-contractile tails and includes about 60% of tailed phages. On the bases of morphology and host genus, the V. harveyi bacteriophages isolated in the present study are considered to fit best in the order Caudovirales of the Family Siphoviridae.
The present study has shown that the infectivity of V. harveyi phages was not inhibited by ribonuclease, trypsin and protease. In contrast, V. harveyi phages infectivity was inhibited by lysozyme perhaps indicating that the enzyme interfered with adsorptions of the phages to the bacterial cell wall. The effects of enzymatic treatment on several bacteriophages have been reported by Valyasevi et al. (1990) and Callegari et al. (1998). Binetti et al. (2002) reported that proteinase-K treatment did not change the adsorption ability of phage particles. MAV1 phage infectivity was found to be unaffected by treatment with RNAase, trypsin, or proteinase K (Voelker and Dybvig, 1998).
It has been shown here that the infectivity of Vibrio bacteriophages was unaffected by the treatment with chloroform, acetone, methyl alcohol and ethyl alcohol at 37°C for 1 hr. Kakimoto and Nagatomi (1972) reported that all six bacteriophages from the host bacteria Aeromonas, Vibrio and related organisms isolated from Kinko Bay, Japan, were fairly resistant to chloroform. One of the isolates was found to be stable in saturated chloroform, whilst the ability of another of the isolates to form plaques was reduced to 10% and two further isolates were completely inactivated by the treatment (Hidaka and Tokushige, 1978; Nakamura et al., 2003). Bacteriophage BCJA1 (Bacillus clarkii) was found to be stable to exposure to chloroform indicating that it probably does not contain lipid (Jarrell et al., 1997). In contrast, polyhedral, filamentous and pleomorphic (PFP) phages, comprising about 190 of the 4500 viruses studied by Ackermann (2003), were shown to be highly sensitive to chloroform and ether.
The results of the present study suggest that the Vibrio bacteriophages survived treatment with Triton X 100 whereas they did not survive treatment with 1% SDS. SDS might have hydrolyzed peptide bonds and removed membrane-bound proteins, thus affecting the infectivity, growth, survival and multiplication of the bacteriophages. These studies suggest that the phages contain proteins and that some detergents such as SDS present in the environment can affect bacteriophage adsorption and infectivity. They might therefore have a role in the control of phage infectivity. On the other hand, Binetti et al. (2002) found that SDS 1% treatment did not affect the adsorption ability of phage particles. Further studies on the proteins and lipids of vibriophages may help us to understand their roles in the life cycle and infectivity of bacteriophages.
pH has been shown to be an important control factor, influencing attachment, infectivity, intracellular replication and multiplication of phages. Generally, studies on the lytic activity of bacteriophages of Vibrio spp. have shown them to be most sensitive to pH values less than 5 and over 10 (Hirayama et al., 1993; Jarrell et al., 1997; Nakai et al., 1999; Pirisi, 2000; Jepson and March, 2004). In the present study, the phages of V. harveyi were found to be inactivated at pH values of 3 or less and at pH 12 and greater. In the range pH 4 to 11 the lytic activity of the phages varied, with the maximum at pH 7. Lytic activity was preserved for a greater range of pH values above pH 7 than below pH 7.
All the vibriophage isolates in the present study exhibited optimal survival at 37°C while complete inactivation occurred at a temperatures exceeding 55°C. Thus the present study has shown that temperature may be an important regulating factor on the infectivity of Vibrio bacteriophages. The VPP97 phage of V. parahaemolyticus, has been shown to be stable up to a temperature of 65°C but was totally inactivated at 70°C (Pirisi, 2000). Jepson and March (2004) reported that the phages were highly stable at 42°C, 37°C and at 4°C for up to 181 days. The current study has clearly underlined the fact that physico-chemical parameters are very important for the survival and infectivity of bacteriophages of Vibrio sp., in the aquaculture environment.
In conclusion, the present study highlights the occurrence of bacteriophages of Vibrio sp., in the shrimp aquaculture environment of South India. The study may assist in the early diagnosis of pathogenic Vibrio sp., that infect the crop and may point the way to non-chemotherapeutic control and preventative measures based on bacteriophage therapy.
ACKNOWLEDGMENTS
PR is thankful for DST-FIST. PS thanks the State Government, Tamilnadu, for the award fellowship.
REFERENCES
- Ackermann, H.W., 1996. Frequency of morphological phage descriptions in 1995. Arch. Virol., 141: 209-218.
CrossRefDirect Link - Ackermann, H.W., 2003. Bacteriophage observations and evolution. Res. Microbiol., 154: 245-251.
CrossRefPubMedDirect Link - Binetti, A.G., A. Quiberoni and J.A. Reinheimer, 2002. Phage adsorption to S. thermophilus. Infuence of environmental factors and characterization of cell-receptors. Food Res. Int., 35: 73-83.
Direct Link - Callegari, M.L., B. Riboli, J.W. Sanders, P.S. Cocconcelli, J. Kok, G. Venema and L. Morelli, 1998. The S-layer gens of Lactobacillus helveticus CNRZ 892: Cloning, sequence and heterologous expression. Microbiology, s144: 719-726.
Direct Link - Catherine, S.B. and M.D. Susan, 2004. Talking about a revolution: The impact of site-specific recombinases on genetic analyses in mice. Dev. Cell, 6: 7-28.
Direct Link - Chythanya, R., I. Karunasagar and I. Karunasagar, 2002. Inhibition of shrimp pathogenic vibrios by a marine Pseudomonas I-2 strain. Aquaculture, 208: 1-10.
CrossRefDirect Link - Ellis, E.L. and M. Delbruck, 1939. The growth of bacteriophage. J. Gen. Physiol., 22: 365-384.
CrossRefDirect Link - Fuhrman, J.A. and C.A. Suttle, 1993. Viruses in marine planktonic systems. Oceanography, 6: 51-63.
Direct Link - Gantzer, C., A. Maul, M. Audic and L. Schwartzbrod, 1998. Detection of infectious enteroviruses, enterovirus genomes, somatic coliphages and Bacteroides fragilis phages in treated wastewater. Applied Environ. Microbiol., 64: 4307-4312.
Direct Link - Gram, L., J. Melchiorsen, B. Spanggaard, I. Huber and T.F. Nielsen, 1999. Inhibition of Vibrio anguillarum by Pseudomonas fluorescens AH2, a possible probiotic treatment of fish. Applied Environ. Microbiol., 65: 969-973.
Direct Link - Heldal, M. and G. Bratbak, 1991. Production and decay of viruses in aquatic environments. Mar. Ecol. Prog. Ser., 72: 205-212.
Direct Link - Jarrell, K.F., T. Vydykhan, P. Lee, M.D. Agnew and N.A. Thomas, 1997. Isolation and characterization of bacteriophage BCJA1, a novel temperate bacteriophage active against the alkaliphilic bacterium, Bacillus clarkii. Extremophiles, 1: 199-206.
Direct Link - Jepson, C.D. and J.B. March, 2004. Bacteriophage lambda is a highly stable DNA vaccine delivery vehicle. Vaccine, 22: 2413-2419.
Direct Link - Jiang, S.C., C.A. Kellogg and J.H. Paul, 1998. Characterization of marine temperate phage-host systems isolated from Mamala Bay, Oahu, Hawaii. Applied Enviorn. Microbiol., 64: 535-542.
Direct Link - Marks, T. and R. Sharp, 2000. Bacteriophages and biotechnology: A review. J. Chem. Technol. Biotechnol., 75: 6-17.
CrossRefDirect Link - Matsuzaki, S., T. Inoue and S. Tanaka, 1998. A vibriophage, KVP40, with major capsid protein homologous to gp23 of coliphage T4. Virology, 242: 314-318.
Direct Link - Matsuzaki, S., T. Inoue, M. Kuroda, S. Kimura and S. Tanaka, 1998. Cloning and sequencing of major capsid protein (mcp) gene of a vibriophage, KVP20, possibly related to T-even coliphages. Gene, 222: 25-30.
Direct Link - Matsuzaki, S., T. Inoue, S. Tanaka, T. Koga, M. Kuroda, S. Kimura and S. Imai, 2000. Characterization of a novel Vibrio parahaemolyticus phage, KVP241 and its relatives frequently isolated from seawater. Microbiol. Immunol., 44: 953-956.
Direct Link - Miller, E.S., J.F. Heidelberg, J.A. Eisen, W.C. Nelson and A.S. Durkin et al., 2003. Complete genome sequence of the broad-host-range vibriophage KVP40: Comparative genomics of a T4-related bacteriophage. J. Bacteriol., 185: 5220-5233.
Direct Link - Nakai, T., R. Sugimoto, K.H. Park, S. Matsuoka, K. Mori, T. Nishioka and K. Maruyama, 1999. Protective effects of bacteriophage on experimental Lactococcus garvieae infection in yellow tail. Dis. Aquat. Organ., 37: 33-41.
Direct Link - Nakai, T. and S.C. Park, 2002. Bacteriophage therapy of infectious diseases in aquaculture. Res. Microbiol., 153: 13-18.
Direct Link - Nakamura, M., K. Tsumoto, I. Kumagai and K. Ishimura, 2003. A morphologic study of filamentous phage infection of Escherichia coli using biotinylated phages. FEBS Lett., 536: 167-172.
Direct Link - Paul, J.H., J.B. Rose, S.C. Jiang, P. London, X. Xhou and C. Kellogg, 1997. Coliphage and indigenous phage in Mamala Bay, Oahu, Hawaii. Applied Environ. Microbiol., 63: 133-138.
Direct Link - Pelon, W., R.J. Siebeling, J. Simonson and R.B. Luftwig, 1995. Isolation of bacteriophage infections for Vibrio vulnificus. Curr. Microbiol., 30: 331-336.
Direct Link - Shaffer, J.J., L.M. Jacobsen, J.O Schrader, K.W. Lee, E.L. Martin and T.A. Kokjohn, 1999. Characterization of Pseudomonas aeruginosa bacteriophage UNL-1, a bacterial virus with a novel UV-A inducible DNA damage reactivation phenotype. Applied Environ. Microbiol., 65: 2606-2613.
Direct Link - Valyasevi, R., W.E. Sandine and B.L. Geller, 1990. The bacteriophage kh receptor of Lactococcus lactis subsp. cremoris KH is therhamnose of the extracellular wall polysaccharide. Applied Environ. Microbiol., 56: 1882-1889.
Direct Link - Westerdahl, A., J.C. Olsson, S. Kjelleberg, P.L. Conway, 1991. Isolation and characterization of turbot (Scophtalmus maximus)-associated bacteria with inhibitory effects against Vibrio anguillarum. Applied Environ. Microbiol., 57: 2223-2228.
Direct Link - Withey, S., E. Cartmell, L.M. Avery and T. Stephenson, 2005. Bacteriophages-potential for application in wastewater treatment processes. Sci. Total Environ., 339: 1-18.
Direct Link