
Nesreen Moustafa Omar
Department of Histology and Cell Biology, Faculty of Medicine, Mansoura University, Egypt
Trends in Medical Research
Year: 2014 | Volume: 9 | Issue: 1 | Page No.: 17-32







ABSTRACT
Myoepithelial cells are highly specialized cells which express cytoskeletal and contractile proteins such as α-smooth muscle actin (α-SMA). This study was performed in order to determine the immunohistochemical and ultra structural features of mammary myoepithelial cells during pregnancy and lactation. Fifteen adult female albino rats assigned into 3 groups; group I (virgin), group II (pregnant) and group III (lactating); were used in this study. Specimens from the mammary gland were obtained and processed for light and electron microscopy. Paraffin sections of the mammary gland were prepared and stained with H and E stain and immunohistochemical stain using anti α-SMA antibody. In the resting mammary gland, the myoepithelial cells (ME) formed a single continuous layer around the mammary ducts. During pregnancy, ME cells were more developed forming multilayered sheaths around the ducts which may protrude to the stroma. Around the acini, myoepithelial cells were branched forming a discontinuous layer around the acinar cells. During lactation, ME cell were stretched, faintly stained, discontinuous and acquired some ultra structural features of smooth muscles. It could be concluded that mammary myoepithelial cells undergo morphological alterations which correlate with the physiological requirements during pregnancy and lactation.
PDF Abstract XML References Citation
Received: September 05, 2013;
Accepted: January 20, 2014;
Published: March 29, 2014
How to cite this article
Nesreen Moustafa Omar, 2014. Immunohistochemical and Ultra Structural Study of Mammary Myoepithelial Cells in Pregnant and Lactating Rats. Trends in Medical Research, 9: 17-32.
URL: https://scialert.net/abstract/?doi=tmr.2014.17.32
URL: https://scialert.net/abstract/?doi=tmr.2014.17.32
INTRODUCTION
The mammary gland is considered as a modified sweat gland which is present in the subcutaneous connective tissue (Monteiro-Riviere and Stinson, 1998). The gland is formed of a branching tree-like network of duct system that ends in secretory alveoli, when fully developed. The mammary ducts and alveoli are lined with two cell layers, luminal epithelial and basal myoepithelial. The contractile myoepithelial cells surround the ducts and alveoli and the whole epithelial system is enclosed by a continuous basement membrane (Pitelka, 1983; Russo and Russo, 1987; Castro et al., 2006).
Myoepithelial cells are highly specialized cells which have a complex phenotype, combining epithelial and smooth muscle features. They are derived from ectoderm, differentiate toward smooth muscles and acquire smooth muscle-like ultra structure and contractility (Redman et al., 1980; Deugnier et al., 1995; Uematsua et al., 2005). They express cytoskeletal and contractile proteins such as α-smooth muscle actin (α-SMA) which has been long used as a phenotypic marker for mammary myoepithelial cells (Sapino et al., 1993; Deugnier et al., 2002; Santos et al., 2010).
Contraction of myoepithelial cells is responsible for ejection of milk under the control of the posterior pituitary hormone, oxytocin since specific oxytocin receptors are present in their plasma membranes (Redman et al., 1980; Adan et al., 1995). The function of myoepithelial cells in the mammary gland is more than just contractility (Adriance et al., 2005). They synthesize basement membrane components such as collagen IV, laminin-1, laminin-5 and fibronectin that regulate ductal growth (Dickson and Warburton, 1992; Ekblom et al., 2003). During development, myoepithelial cells synthesize the basement membrane component laminin-1 which is responsible for luminal cell polarity (Runswick et al., 2001; Gudjonsson et al., 2002). They affect the proliferation, survival and differentiation of luminal cells, modify stromal-epithelial interactions and play a major role in mammary morphogenesis (Niranjan et al., 1995). In addition, myoepithelial cells are responsible for proper coordination of all of epithelial cell activities to maintain normal breast function (Ronnov-Jessen et al., 1996; Sternlicht et al., 1997).
It has been reported that a number of type II tumor suppressor genes are expressed by myoepithelial cells (Sager, 1997; Bissell and Radisky, 2001) ME cells have many antitumorigenic properties, such as inhibition of tumor cell invasion and angiogenesis (Sternlicht et al., 1996; Nguyen et al., 2000). ME cells have been shown to function as natural tumour suppressors. These cells can stimulate apoptosis of luminal cells, supress angiogenesis and make a natural barrier preventing metastasis (Deugnier et al., 2002). Therefore, the loss or change of myoepithelial cell function and immunoreactivity may have a possible role in the development of breast cancer (Adriance et al., 2005).
The main development of mammary glands occurs in the postnatal period; however, marked morphological changes occur in the mammary gland during pregnancy and lactation (Deugnier et al., 2002). The effect of pregnancy and lactation on the myoepithelial cell has not been fully demonstrated especially at the ultrastrural level. To discover the relationship between myoepithelial cell and hormones of pregnancy and lactation such as estrogen and progesterone is also important in order to understand the biological behaviour of myoepithelial celll which may be related to the development of breast neoplastic lesions. Therefore, the current study was performed in order to declare the immunohistochemical and ultrastructural features of mammary myoepithelial cells during pregnancy and lactation.
MATERIALS AND METHODS
Fifteen adult female albino rats with average age (12-16 weeks) were used in this study. The animals were housed in separate cages under constant environmental conditions and were allowed free access to food and water.
The rats were assigned into three groups (5 rats each):
| Group I | : | Virgin female rats which were bred separately from males |
| Group II | : | Pregnant rats on the 14th day of pregnancy. The animals were bred with males overnight and the next day was considered as the first day of pregnancy |
| Group III | : | Lactating rats on the 7th day of lactation |
The animals from each group were anesthetized with intraperitoneal thiopental sodium (10-15 mg kg-1). From each rat, the left first abdomino-inguinal mammary gland was excised and processed for light and electron microscopy.
Light microscopic study: Paraffin sections (5 μm thick) were prepared and stained with:
| • | Haematoxylin and Eosin (H and E) stain (Gamble and Wilson, 2002) |
| • | Immunohistochemical stain: By using anti-α smooth muscle actin. (Hsu et al., 1981) |
Immunohistochemical staining: Immunocytochemical reaction was performed by using avidin biotin peroxidise complex (ABC) technique. Specific mouse monoclonal antibody (Monoclonal Mouse Anti-human, smooth muscle actin, clone 1A, code M0581) was applied at a dilution of 1:100. Cross reactivity of this antibody with other species has been reported.
Technique: Endogenous peroxidase activity was inhibited by 3% H2O2 in distilled water for 30 min. Sections were then washed in tap water for 30 min and in distilled water for 10 min. Non specific binding of antibodies was blocked by incubation with normal goat serum (DAKO X 0907, Carpinteria, CA) and Phosphate Buffer Solution (PBS) at a dilution of 1:4. Then, sections were incubated with monoclonal mouse antisera against α-SMA (Dako, Hamburg, Germany) which was applied at a dilution of 1:100. The incubated sections were then left at room temperature for 1 h. The sections were then washed in PBS 3x3 min. and incubated with biotinylated anti-mouse IgG (DAKO LSAB 2 Kit) followed by washing in PBS 3x3 min. and then incubation with (ABC) complex solution (DAKO LSAB 2 Kit). The antibody bound to sections was visualized by treating with 0.05% (W/V) 3, 3-diaminobenzidine (DAB) tetrachloride (Sigma Chemicals Co., St. Louis, MO, USA) in 10 mmol Tris-buffered saline. Then, the sections were stained by Mayer’s haematoxylin as counter stain. Negative control sections were put under the same conditions after omitting primary antibodies. Positive cytoplasmic controls were included within the tissue sections and positive cytoplasmic reaction was seen in the wall of the gut.
Electron microscopic study: (Woods and Stirling, 2008). Small fragments, about 1 mm 3 of tissues, were obtained and processed to prepare semithin and ultrathin sections for transmission electron microscopic study.
Procedure: Specimens were first fixed in 2.5% gluteraldehyde in phosphate buffer at 4°C for 2 h followed by washing with Phosphate Buffer Solution (PBS) three times (10 min each). The samples were then post fixed in 1% osmium tetroxide for 30 min then washed with PBS for three times (10 min each) followed by dehydration in ascending grades of ethyl alcohol. Then, the specimens were infiltrated with acetone for 1 h followed by embedding in Araldite 502 resin and polymerized at 60°C for 24 h. Semi-thin and ultrathin sections were obtained by using LEICA Ultracut (UCT) ultramicrotome. Ultrathin sections were stained by a double staining technique of 2% uranyl acetate for 10 min, followed by Reynold's lead citrate solution for another 10 min.
Ultrathin sections were then examined by using JEOL-JEM-100 SX transmission electron microscope in the electron microscopy unit, Tanta University, Egypt.
RESULTS
Group I: in H and E stained sections, the parenchyma of the resting mammary gland consisted of lobules which were predominantly formed of adipose tissue and branched duct system. This duct system comprised small intralobular ducts within the lobules and larger interlobular ducts in the interlobular connective tissue septa. The ductal epithelium consisted of an inner layer of cubical luminal epithelial cells and an outer layer of myoepithelial cells lying on the basement membrane (Fig. 1). In immunostained sections, positive reaction against α-SMA was detected in myoepithelial cells in the wall of the mammary ducts (Fig. 2a). The myoepithelial cells appeared spindle shaped forming a single continuous layer surrounding the luminal epithelial cells in the wall of intralobular (Fig. 2b) and interlobular ducts (Fig. 2c). In the rudimentary acini, ME cells appeared branched with many cytoplasmic processes forming a continuous layer around the acinar cells (Fig. 2d). In the electron micrographs, the myoepithelial cells appeared spindle shaped and located parallel to the wall of the duct. The cytoplasm of ME cell contained bundles of tonofilaments, cytoplasmic vacuoles and scanty organelles (Fig. 3).
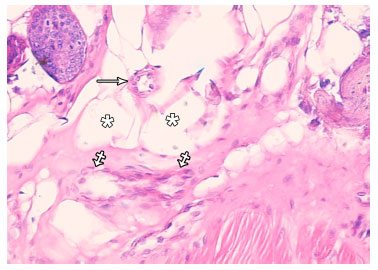 | |
| Fig. 1: | Photomicrograph of resting mammary gland showing a small intralobular duct (arrow), an interlobular duct (crossed arrows) and adipose tissue (asterisks). (H and EX400) |
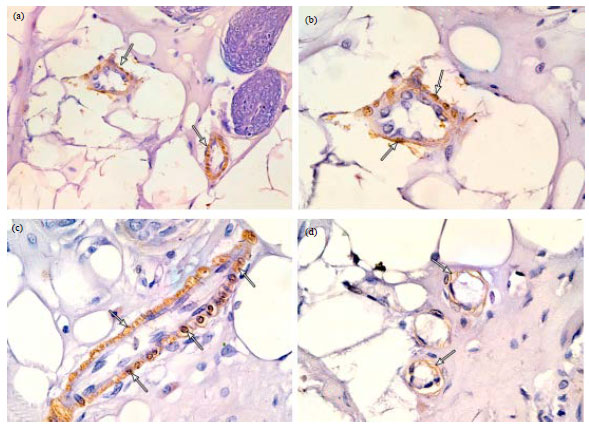 | |
| Fig. 2(a-d): | Photomicrographs of immunostained sections of resting mammary gland showing (a) Positive α-SMA immunoreaction in the ME cells in the wall of two small intralobular ducts (arrows), (b) A higher magnification of the previous figure showing a small intralobular duct surrounded by a single continuous layer of ME cells (arrows), (c) An interlobular duct surrounded by a single and continuous layer of ME cells (arrows) and (d) Vestigial acini surrounded by branched ME cells (arrows). (α-SMA immunostaining; a X400, b, c,d X1000) |
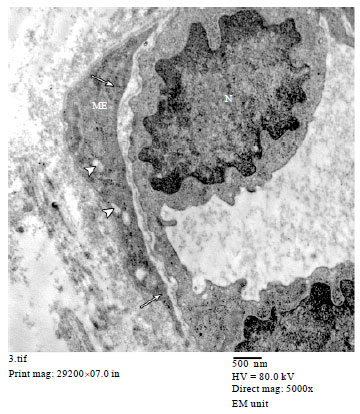 | |
| Fig. 3: | An electron micrograph of resting mammary gland showing a small duct lined with luminal epithelial cells with rounded to oval nucleus (N). A spindle shaped myoepithelial cell (ME) containing small cytoplasmic vacuoles (arrow heads) and bundles of tonofilaments (arrows) is also seen. (TEMX5000) |
In group II rats, marked developmental changes occurred in the mammary gland. Lobules of developing acini lined with cubical luminal epithelial cells and numerous proliferating ducts were observed (Fig. 4a, b). In immunostained sections, positive reaction against α-SMA was detected in the myoepithelial cells, wall of blood vessels and in bundles of smooth muscles scattered in the stroma (Fig. 5a). The ME cells formed a well developed sheath around the ducts which may protrude the surrounding connective tissue (Fig. 5b). Around the acini, ME cells were branched and their processes formed a discontinuous layer around the acinar cells (Fig. 5c). In electron micrographs, ductal myoepithelial cells showed well developed bundles of tonofilaments. A multilayered sheath formed of myoepithelial cell processes could be seen around the ducts, separating them from the periductal stroma (Fig. 6a, b). The alveolar myoepithelial cells showed many cytoplasmic processes and were attached to the acinar cells by desmosomes to the basal lamina by hemidesmosomes (Fig. 7, 8). However, in some areas, the ME cell layer was deficient and the acinar cells appeared in direct contact with the basal lamina (Fig. 8).
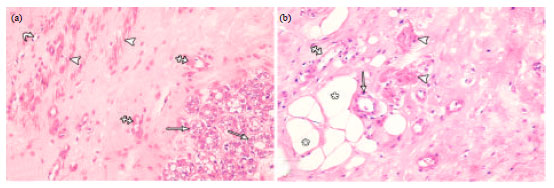
In group III rats, the mammary gland was fully developed and predominantly composed of large clusters of secretory alveoli. Some of the alveoli appeared small and lined with cubical acinar cells. Others appeared dilated, distended with secretions and lined with flattened acinar cells (Fig. 9a). The duct system comprised small and medium sized ducts lined with cubical luminal cells (Fig. 9b) and larger ducts distended with secretions and lined with flattened luminal epithelial cells (Fig. 9c).
 | |
| Fig. 4(a-b): | Photomicrographs of pregnant rat mammary gland. (a) Newly formed acini (arrows) lined with cubical luminal epithelial cells with rounded nuclei and vacuolated cytoplasm, proliferating ducts (crossed arrows), bundles of smooth muscle fibres (arrow heads) and a blood capillary (curved arrow) and (b)Proliferating ducts of different sizes; an intralobular duct (arrow) lined with luminal cubical cells and an interlobular duct (crossed arrow) lined with flattened luminal epithelial cells. Note adipose tissue (asterisks) and bundles of smooth muscle fibers (arrow heads) (H and EX400) |
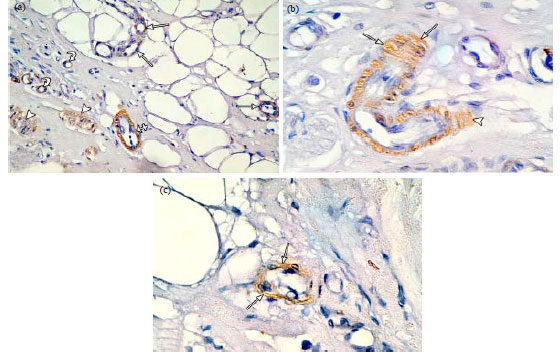 | |
| Fig. 5(a-b): | Photomicrographs of immunostained sections of pregnant rat mammary gland. (a) Positive α-SMA immunoreaction in the ME cells in the wall of proliferating intralobular ducts (arrows) and in the wall of interlobular duct (crossed arrow). Positive immunoreaction is also observed in the wall of blood vessels (curved arrows) and in bundles of smooth muscle fibres (arrow heads) in the stroma, (b) An interlobular duct surrounded by a continuous well developed sheath of ME cells (arrows) which protrude to the surrounding stroma (arrowhead) and (c) A newly formed acinus surrounded by a discontinuous layer of branched ME cells (arrows). (α-SMA immunostaining; a X400, b, c X1000) |
 | |
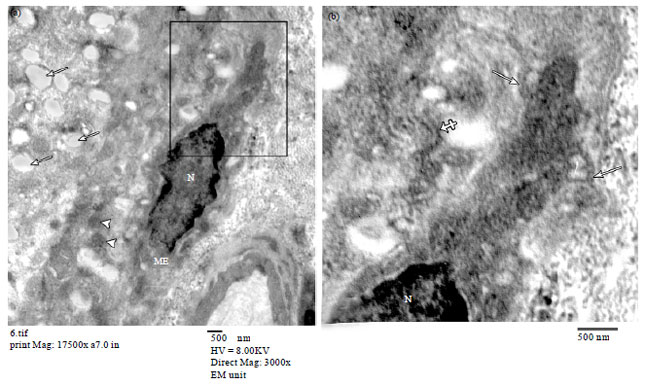
| Fig. 6(a-b): | Electron micrographs of pregnant rat mammary gland. (a) Small duct lined with luminal epithelial cells with rounded nucleus (N) and surrounded by a myoepithelial cell with oval flattened nucleus (n). Note cytoplasmic processes of ME cells rich in tonofilaments forming a multilayered sheath (arrows) around the duct and (b) A large duct lined by luminal epithelial cells with oval flattened nuclei (N) and surrounded by a multilayered sheath (arrows) formed by ME cell processes (TEM a X3000, b X2500) |
In immunostained sections, an almost continuous layer of stretched ME cells with elongated processes was observed around the intralobular and interlobular ducts (Fig. 10). In larger dilated ducts, a markedly discontinuous layer of faintly stained ME cells was observed (Fig. 11a). Occasional α-SMA positive fusiform cells with rounded nuclei were observed in close proximity to some large ducts (Fig. 11b). Around acini, ME cells formed a discontinuous layer around the acinar cells (Fig. 12a). This discontinuity was more obvious in larger acini distended with secretions (Fig. 12b). In electron micrographs, the cytoplasm of ME cells appeared abundant and contained bundles of microfilaments, dense bodies and more cellulalr organelles such as mitochondria. These cells showed many cytoplasmic processes and were attached to the acinar cells by intercellular connections and to the basal lamina by hemidesmosomes (Fig. 13).
DISCUSSION
Myoepithelial cells are highly specialized cells which have a complex phenotype, combining epithelial and smooth muscle features. They are derived from ectoderm, differentiate toward smooth muscles and acquire smooth muscle-like ultra structure and contractility (Redman et al., 1980; Deugnier et al., 1995; Uematsua et al., 2005). The contractile protein α-SMA is mainly found in cells having contractile functions and has been long used as a phenotypic marker for mammary myoepithelial cells (Deugnier et al., 2002; Santos et al., 2010).
 | |
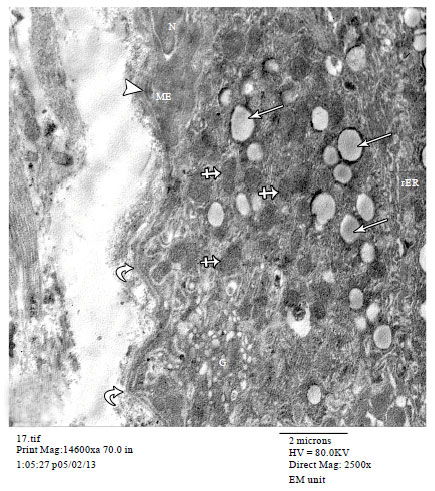
| Fig. 7(a-b): | Electron micrographs of pregnant rat mammary gland (a) showing an acinar cell containing secretory vesicles (arrows) and mitochondria (arrowheads). Under the acinar cell, a myoepithelial cell (ME) with oval flattened nucleus (N) is seen. (b) Zoomed in part of Fig. 7a showing a part of ME cells containing the nucleus (N) and many cytoplasmic processes (arrows). A desmosome (crossed arrow) between the acinar cell and ME cell is also observed. (a TEMX3000) |
Myoepithelial cells affect the proliferation, survival and differentiation of luminal cells, modify stromal-epithelial interactions and play a major role in mammary morphogenesis (Niranjan et al., 1995). The mammary gland undergoes marked morphological changes during pregnancy and lactation (Deugnier et al., 2002). Therefore, this study was performed in order to determine the immunohistochemical and ultra structural features of mammary myoepithelial cells during these physiological conditions.
The present work revealed that the resting mammary gland consisted mainly of adipose tissue and a branched duct system. Similarly, Morroni et al. (2004) mentioned that adipocytes comprised the most prominent component of the resting mouse mammary gland. As reported in this work, the duct system was lined by an inner layer of luminal epithelial cells and an outer layer of myoepithelial (ME) cells. Similar findings were described in canine (Santos et al., 2010) mice and rat resting mammary gland (Sangai et al., 2006; Kariagina et al., 2007).
Immunostained sections of the resting mammary gland revealed positive immunoreaction against α-SMA in the myoepithelial cells which formed a single continuous layer around the intralobular and interlobular ducts. Similar findings were reported by Jin et al. (2000).
 | |
| Fig. 8: | An electron micrograph of pregnant rat mammary gland showing an acinar cell containing mitochondria (crossed arrows), secroetory vesicles (arrows), rough endoplasmic reticulum (rER) and Golgi saccules (G). A part of myoepithelial cell (ME) with its nucleus (N) is also seen. Note the hemidesmosome (arrowhead) between the ME cell and the basal lamina. Areas where the acinar cell lies in direct contact (curved arrows) with the basal lamina are also seen. (TEMX2500) |
It was reported that ME cells express contractile and cytoskeletal proteins such as α-smooth muscle actin (Sapino et al., 1993). Actin microfilaments of myoepithelial cells provide contractility to facilitate milk ejection in response to the posterior pituitary hormone, oxytocin (Hamperl, 1970; Adan et al., 1995).
In the present work, the alveolar ME of resting mammary gland appeared branched with many cytoplasmic processes forming a continuous layer around the acinar cells of the vestigial acini. Similarly, few residual acini surrounded by a continuous layer of myoepithelial cells were reported by Santos et al. (2010) in canine resting mammary gland. On the other hand, Emerman and Vogl (1986) reported a discontinuous basket-like network formed of stellate shaped acinar myoepithelial cells in the mouse resting mammary gland.
The present ultra structural study of the resting mammary gland revealed spindle shaped myoepithelial cells which contained bundles of tonofilaments and cytoplasmic vacoules and located parallel to the wall of the duct. Emerman and Vogl (1986) mentioned that the myoepithelial cells, in the mouse resting mammary gland, were longitudinal in shape and arranged in parallel to the long axis of ducts.
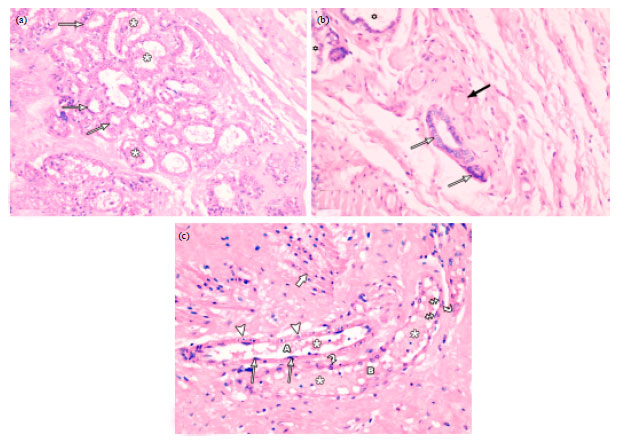 | |
| Fig. 9(a-c): | Photomicrographs of lactating mammary gland. (a) Lobules of secretory acini lined with low cuboidal cells (arrows) with pale vacuolated cytoplasm. Most of the acini are distended with secretions (asterisks) and lined with flattened epithelial cells. (b) Small and medium sized ducts (arrows) lined with cubical luminal epithelial cells and some small ducts (black arrow) distended with secretions and lined with flattened luminal epithelial cells. Some secretory acini (asterisks) are also seen. (c) A large duct (A) lined with flattened luminal epithelial cells (arrows) and surrounded by a continuous layer of ME cells (arrowheads). A dilated duct (B) lined with flattened luminal epithelial cells (crossed arrows) and surrounded by a discontinuous layer of ME cells (curved arrows) is also observed. Both ducts are distended with secretions (asterisks). Note bundles of smooth muscle fibres (thick arrow) in the stroma. (H and EX400) |
The presence of endocytotic vesicles and contractile microfilaments in myoepithelial cells was also previously reported by Gugliotta et al. (1988).
In pregnant rats, the mammary gland showed marked proliferative changes with numerous proliferating ducts and developing acini. It has been demonstrated that the proliferative activity in the rodent mammary gland reach a peak during pregnancy (Nagasawa and Vorherr, 1977). Progesterone and the pituitary hormone, prolactin coordinate mammary epithelial differentiation during pregnancy resulting in 100-fold increase in the cell number and development of secretory epithelium (Oakes et al., 2008). It has been known that estradiol and progesterone are required for the remodelling of mammary gland (Ruan et al., 2005). Estrogens promote end-bud development and duct elongation while progesterone is involved in lobuloalveolar development (Aupperlee and Haslam, 2007).
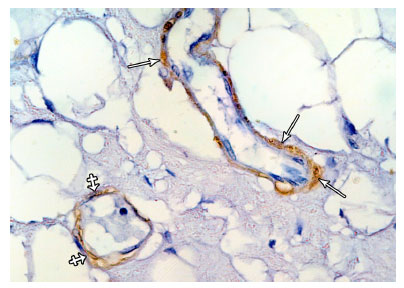 | |
| Fig. 10: | A photomicrograph of immunostained sections of lactating mammary gland showing an almost continuous layer of stretched ME cells with elongated processes around an intralobular (crossed arrows) and interlobular ducts (arrows). (α-SMA immunostaining X1000) |
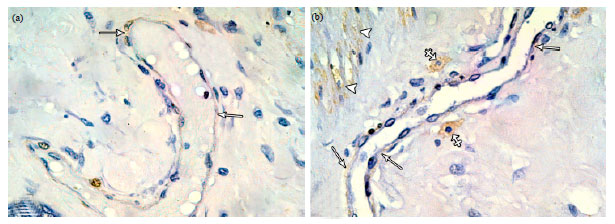 | |
| Fig. 11(a-b): | Photomicrographs of immunostained sections of lactating mammary gland (a) A large dilated duct surrounded by a discontinuous layer of faintly stained ME cells (arrows). (b) A large duct surrounded with a discontinuous layer of faintly stained ME cells (arrows). α-SMA positive fusiform cells (crossed arrows) are seen in close vicinity to the duct. Note smooth muscle bundles (arrowheads) showing α-SMA positive reaction in the stroma. (α-SMA immunostaining X1000) |
It has been also suggested that mammary epithelial cells are influenced by sex steroids resulting in modification of mammary gland development and function (Li et al., 2010).
In the present study, immunostained sections of pregnant rat mammary gland demonstrated a well developed sheath of ME cells around the ducts which may protrude to the periductal stroma. This was confirmed by the ultra structural study which showed a multilayered sheath formed by ME cell processes around the ducts. Jin et al. (2000) reported stratified ME cells around the ducts of pregnant rat mammary gland.
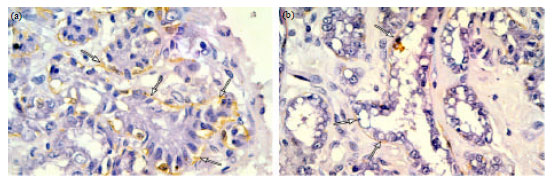 | |
| Fig. 12(a-b): | Photomicrographs of immunostained sections of lactating mammary gland. (a) Clusters of large secretory acini surrounded by a discontinuous layer of ME cells (arrows). (b) Clusters of dilated secretory acini distended with secretions and surrounded by a discontinuous layer of ME cells (arrows) with wider gaps in between. (α-SMA immunostaining X1000) |
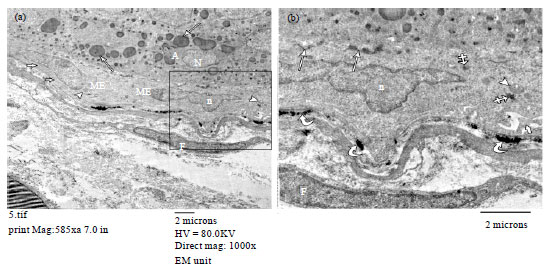 | |
| Fig. 13(a-b): | Electron micrographs of lactating mammary gland (a) Showing an acinar cells (A) containing rounded to oval nuclei (N) and secretory vesicles (arrows). Two myoepithelial cells (ME) with oval flattened nuclei (n), bundles of microfilaments (thick arrows) and mitochondria (arrowheads) are observed. A spindle shaped fibroblast (F) is also seen. (b) Zoomed in part of (a) showing a part of myoepithelial cell with oval flattened nucleus (n), mitochondria (arrowhead), cytoplasmic processes (thick arrow) and dense bodies (crossed arrows). Hemidesmosomes (curved arrows) between the ME cell and the basal lamina and intercellular connections (arrows) between ME cell and acinar cell are observed. A part of fibroblast (F) is also seen. (a, TEMX1000) |
Increased proliferative activity of the myoepithelial cells during pregnancy has also been proven as shown by bromodeoxyuridine (BrdU) and [3H]-thymidine uptake in the rat and mice mammary gland (Joshi et al., 1986; Sapino et al., 1990). Progesterone receptors have been demonstrated in myoepithelial cells suggesting that progesterone may directly stimulate proliferation and/or terminal differentiation of myoepithelial cells (Kariagina et al., 2007). It has also been shown that treatment with estrogen and progesterone increased recruitment of BM-derived myoepithelial cells and periductal fibroblasts to the mammary gland (Sangai et al., 2006). The presence of a well developed sheath of ME cells around the ducts may provide a potent barrier preventing the invasion of the mammary stroma by the highly proliferating epithelial elements during pregnancy.
In immunostained sections of pregnant rat mammary gland, the alveolar ME cells appeared branched and their processes formed a discontinuous layer around the acinar cells. Jin et al. (2000) reported that the cytoplasmic processes of the alveolar myoepithelial cells, in the pregnant rat mammary gland, were thin with a dotted appearance compared to the continuous arrangement in the resting phase. Emerman and Vogl (1986) reported stellate shaped ME cells forming a basket-like network around the luminal cells in pregnant mouse mammary gland. At the ultra structural level, the myoepithelial cells showed many cytoplasmic processes and appeared attached to the acinar cells by desmosomes and to the basal lamina by hemidesmosomes. Desmosomal connections between the ME cells and luminal epithelial cells were previously described by Gugliotta et al. (1988). The presence of hemidesmosomes between the myoepithelial cells and the basement membrane was also reported (Koukoulis et al., 1991; Bergstraesser et al., 1995).
In the current study, the lactating mammary gland was fully developed and predominantly composed of large clusters of secretory alveoli distended with secretions. Large ducts distended with secretions were also seen. Similar structure of rat lactating mammary gland was described by Deugnier et al. (1995). A complex milieu of hormones marked by a drop in the level of progesterone stimulates epithelial changes leading to lactation which is typically maintained by the suckling response of the newborn (Neville et al., 2002; Pang and Hartmann, 2007).
In the immunostained sections of lactating mammary gland, an almost continuous layer of stretched ME cells with elongated processes was seen around the ducts. However, marked discontinuity and weak immunoreactivity of ME cell layer were observed in the large dilated ducts. Alkafafy et al. (2012) demonstrated a continuous layer of spindle shaped myoepithelial cells oriented parallel to the long axis of ducts in bovine lactating mammary gland. Jin et al. (2000) reported a single layer of ME cells around markedly dilated ducts in lactating mammary gland. Upon contraction, the ductal myoepithelial cells oriented parallel to the long axis decrease the length and increase the diameter of the ducts to eject the milk (Emerman and Vogl, 1986). Bundles of smooth muscle fibres showing α-SMA positive immunoreaction were also observed in the stroma of lactating mammary gland. Similarly, Richardson (2009) reported smooth muscle forming scattered inter-lobular bundles in the goat lactating mammary gland and suggested a possible role of their contraction in the process of milk ejection.
In the present work, a discontinuous layer of ME cells was observed around the acini of the lactating mammary gland. This discontinuity was more evident in dilated acini distended with secretions. Similarly, Alkafafy et al. (2012) reported a discontinuous layer of stellate-shaped alveolar myoepithelial cells in bovine lactating mammary gland. Moreover, Jin et al. (2000) mentioned that the cytoplasmic processes of ME cells during lactation, were immunostained vaguely and weakly and the distance of the cytoplasmic processes came to be greater than that observed in the other phases. Emerman and Vogl (1986) reported that the alveolar epithelial cells of pregnant and lactating mammary gland are fully encompassed by the ME cell body and processes.
The current ultra structural study of lactating mammary gland revealed that the myoepithelial cells acquired some of the smooth muscle characters such as abundant cytoplasm, more mitochondria, cytoplasmic dense bodies and bundles of microfilaments. These characters may provide better contractility function needed for milk ejection during lactation. These cells showed many cytoplasmic processes and connections to the acinar cells and to the basal lamina. The cytoplasmic processes of ME cells seem to serve the milk ejecting function during lactation. The presence of dense bodies and cytoplasmic processes in ME was previously described by Gugliotta et al. (1988) and Emerman and Vogl (1986). Moreover it was reported ME cells form distinct desmosomes with luminal epithelial cells and hemidesmosomes with the basement membrane (Koukoulis et al., 1991; Bergstraesser et al., 1995). The connection of ME cells to the basement membrane and the intercellular connections with the luminal epithelial cells facilitate paracrine regulatory mechanisms and play a role in proper coordination of all activities to maintain normal breast function (Ronnov-Jessen et al., 1996, Sternlicht et al., 1997; Gudjonsson et al., 2002).
CONCLUSION
Mammary myoepithelial cells undergo morphological alterations which correlate with the physiological requirements during pregnancy and lactation; so as to provide a barrier function during the highly proliferative state of pregnancy and contractility function during the secretory state of lactation.
REFERENCES
- Adan, R.A., F.W. Van Leeuwen, M.A. Sonnemans, M. Brouns, G. Hoffman, J.G. Verbalis and J.P. Burbach, 1995. Rat oxytocin receptor in brain, pituitary, mammary gland and uterus: Partial sequence and immunocytochemical localization. Endocrinology, 136: 4022-4028.
Direct Link - Adriance, M.C., J.L. Inman, O.W. Petersen and M.J. Bissell, 2005. Myoepithelial cells: Good fences make good neighbors. Breast Cancer Res., 7: 190-197.
Direct Link - Alkafafy, M., R. Rashed and A. Helal, 2012. Immunohistochemical studies on the bovine lactating mammary gland (Bos taurus). Acta Histochem., 114: 87-93.
CrossRefDirect Link - Aupperlee, M.D. and S.Z. Haslam, 2007. Differential hormonal regulation and function of progesterone receptor isoforms in normal adult mouse mammary gland. Endocrinology, 148: 2290-2300.
CrossRefDirect Link - Bergstraesser, L.M., G. Srinivasan, J.C. Jones, S. Stahl and S.A. Weitzman, 1995. Expression of hemidesmosomes and component proteins is lost by invasive breast cancer cells. Am. J. Pathol., 147: 1823-1839.
PubMedDirect Link - Bissell, M.J. and D. Radisky, 2001. Putting tumours in context. Nat. Rev. Cancer, 1: 46-54.
CrossRefDirect Link - Castro, G.D., C.R. de Castro, M.E. Maciel, S.L. Fanelli, E.C. de Ferreyra, M.I.D. Gomez and J.A. Castro, 2006. Ethanol-induced oxidative stress and acetaldehyde formation in rat mammary tissue: Potential factors involved in alcohol drinking promotion of breast cancer. Toxicology, 219: 208-219.
CrossRefDirect Link - Deugnier, M.A., E.P. Moiseyeva, J.P. Thiery and M. Glukhova, 1995. Myoepithelial cell diffeentiation in the developing mammary gland: Progressive acquisition of smooth muscle phenotype. Dev. Dyn., 204: 107-117.
CrossRefDirect Link - Deugnier, M.A., J. Teuliere, M.M. Faraldo, J.P. Thiery and M.A Glukhova, 2002. The importance of being a myoepithelial cell. Breast Cancer Res., 4: 224-230.
Direct Link - Dickson, S.R. and M.J. Warburton, 1992. Enhanced synthesis of gelatinase and stromelysin by myoepithelial cells during involution of the rat mammary gland. J. Histochem. Cytochem., 40: 697-703.
CrossRefDirect Link - Ekblom, P., P. Lonai and J.F. Talts, 2003. Expression and biological role of laminin-1. Matrix Biol., 22: 35-47.
CrossRefPubMedDirect Link - Emerman, J.T. and A.W. Vogl, 1986. Cell size and shape changes in the myoepithelium of the mammary gland during differentiation. Anat. Rec., 216: 405-415.
PubMedDirect Link - Gudjonsson, T., L. Ronnov-Jessen, R. Villadsen, F. Rank, M.J. Bissell and O.W. Petersen, 2002. Normal and tumor-derived myoepithelial cells differ in their ability to interact with luminal breast epithelial cells for polarity and basement membrane deposition. J. Cell Sci., 115: 39-50.
PubMed - Gugliotta, P., A. Sapino, L. Macri, O. Skalli, G. Gabbiani and G. Bussolati, 1988. Specific demonstration of myoepithelial cells by anti-alpha smooth muscle actin antibody. J. Histochem. Cytochem., 36: 659-663.
PubMed - Hamperl, H., 1970. The myothelia (myoepithelial cells). Normal state, regressive changes, hyperplasia, tumors. Curr. Top. Pathol., 53: 161-220.
PubMed - Hsu, S.M., L. Raine and H. Fanger, 1981. Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques: A comparison between ABC and unlabeled antibody (PAP) procedures. J. Histochem. Cytochem., 29: 557-580.
CrossRefPubMedDirect Link - Jin, H.S., S. Umemura, T. Iwasaka and R.Y. Osamura, 2000. Alterations of myoepithelial cells in the rat mammary gland during pregnancy, lactation and involution and after estradiol treatment. Pathol. Int., 50: 384-391.
CrossRefDirect Link - Joshi, K., J.T.B. Ellis, C.M. Hughes, P. Monaghan and A.M. Neville, 1986. Cellular proliferation in the rat mammary gland during pregnancy and lactation. Lab. Invest., 54: 52-61.
PubMed - Kariagina, A., M.D. Aupperlee and S.Z. Haslam, 2007. Progesterone receptor isoforms and proliferation in the rat mammary gland during development. Endocrinology, 148: 2723-2736.
PubMed - Koukoulis, G.K., I. Virtanen, M. Korhonen, L. Laitinen, V. Quaranta and V.E. Gould, 1991. Immunohistochemical localization of integrins in the normal, hyperplastic, and neoplastic breast. Correlations with their functions as receptors and cell adhesion molecules. Am. J. Pathol., 139: 787-799.
PubMed - Li, S., B. Han, G. Liu, S. Li, J. Ouellet, F. Labrie and G. Pelletier, 2010. Immunocytochemical localization of sex steroid hormone receptors in normal human mammary gland. J. Histochem. Cytochem., 58: 509-515.
CrossRefPubMedDirect Link - Morroni, M., A. Giordano, M.C. Zingaretti, R. Boiani and R. De Matteis et al., 2004. Reversible transdifferentiation of secretory epithelial cells into adipocytes in the mammary gland. Proc. Natl. Acad. Sci., 101: 16801-16806.
PubMed - Nagasawa, H. and H. Vorherr, 1977. Rat mammary deoxyribonucleic acid synthesis during the estrous cycle, pregnancy and lactation in relation to mammary tumorigenesis: Its implication for human breast cancer. Am. J. Obstet. Gynecol., 127: 590-593.
PubMed - Neville, M.C., T.B. McFadden and I. Forsyth, 2002. Hormonal regulation of mammary differentiation and milk secretion. J. Mammary Gland Biol. Neoplasia, 7: 49-66.
CrossRefPubMedDirect Link - Nguyen, M., M.C. Lee, J.L. Wang, J.S. Tomlinson, Z.M. Shao, M.L. Alpaugh and S.H. Barsky, 2000. The human myoepithelial cell displays a multifaceted anti-angiogenic phenotype. Oncogene, 19: 3449-3459.
PubMed - Niranjan, B., L. Buluwela, J. Yant, N. Perusinghe and A. Atherton et al., 1995. HGF/SF: A potent cytokine for mammary growth, morphogenesis and development. Development, 121: 2897-2908.
PubMed - Oakes, S.R., R.L. Rogers, M.J. Naylor and C.J. Ormandy, 2008. Prolactin regulation of mammary gland development. J Mammary Gland Biol. Neoplasia, 13: 13-28.
CrossRefDirect Link - Pang, W.W. and P.E. Hartmann, 2007. Initiation of human lactation: Secretory differentiation and secretory activation. J. Mammary Gland Biol. Neoplasia, 12: 211-221.
CrossRefDirect Link - Redman, R.S., L.R. Sweney and S.T. McLaughlin, 1980. Differentiation of myoepithelial cells in the developing rat parotid gland. Am. J. Anatomy, 158: 299-320.
CrossRefDirect Link - Richardson, K.C., 2009. Contractile tissues in the mammary gland, with special reference to myoepithelium in the goat. J. Mammary Gland Biol. Neoplasia, 14: 223-242.
CrossRefDirect Link - Ronnov-Jessen, L., O.W. Petersen and M.J. Bissell, 1996. Cellular changes involved in conversion of normal to malignant breast: Importance of the stromal reaction. Physiol. Rev., 76: 69-125.
Direct Link - Ruan, W., M.E. Monaco and D.L. Kleinberg, 2005. Progesterone stimulates mammary gland ductal morphogenesis by synergizing with and enhancing insulin-like growth factor-I action. Endocrinology, 146: 1170-1178.
CrossRefDirect Link - Runswick, S.K., M.J. O'Hare, L. Jones, C.H. Streuli and D.R. Garrod, 2001. Desmosomal adhesion regulates epithelial morphogenesis and cell positioning. Nat. Cell. Biol., 3: 823-830.
CrossRefDirect Link - Sager, R., 1997. Expression genetics in cancer: Shifting the focus from DNA to RNA. Proc. Nat. Acad. Sci. USA., 94: 952-955.
Direct Link - Sangai, T., G. Ishii, H. Fujimoto, A. Ikehara and T. Ito et al., 2006. Hormonal stimulation increases the recruitment of bone marrow-derived myoepithelial cells and periductal fibroblasts into the mammary gland. Biochem. Biophys. Res. Commun., 346: 1173-1180.
CrossRefDirect Link - Santos, M., R. Marcos and A.M.R. Faustino, 2010. Histological study of canine mammary gland during the oestrous cycle. Reprod. Domestic Anim., 45: e146-e154.
CrossRefDirect Link - Sapino, A., L. Macri, P. Gugliotta, D. Pacchioni, Y.J. Liu, D. Medina and G. Bussolati, 1993. Immunophenotypic properties and estrogen dependency of budding cell structures in the developing mouse mammary gland. Differentiation, 55: 13-18.
CrossRefDirect Link - Sapino, A., L. Macri, P. Gugliotta and G. Bussolati, 1990. Immunocytochemical identification of proliferating cell types in mouse mammary gland. J. Histochemistry Cytochem., 38: 1541-1547.
CrossRefDirect Link - Sternlicht, M.D., P. Kedeshian, Z.M. Shao, S. Safarians and S.H. Barsky, 1997. The human myoepithelial cell is a natural tumor suppressor. Clin. Cancer Res., 3: 1949-1958.
PubMed - Sternlicht, M.D., S. Safarians, S.P. Rivera and S.H. Barsky, 1996. Characterizations of the extracellular matrix and proteinase inhibitor content of human myoepithelial tumors. Lab. Invest., 74: 781-796.
Direct Link - Uematsua, J., Y. Nishizawac, Y. Hirakoa, K. Kitamurad, J. Usukurac, T. Miyata and K. Owaribe, 2005. Both type-I hemidesmosomes and adherens-type junctions contribute to the cell-substratum adhesion system in myoepithelial cells. Eur. J. Cell. Biol., 84: 407-415.
CrossRefDirect Link