
Suhail Rasool
Department of Laboratory Medicine,New Delhi, 110029, India
Madhuri Behari
Neurosciences Center, Department of Neurology, All India Institute of Medical Sciences, New Delhi, 110029, India
M. Irshad
Department of Laboratory Medicine,New Delhi, 110029, India
Vinay Goyal
Neurosciences Center, Department of Neurology, All India Institute of Medical Sciences, New Delhi, 110029, India
B.L. Jailkhani
Department of Laboratory Medicine,New Delhi, 110029, India
Trends in Medical Research
Year: 2008 | Volume: 3 | Issue: 2 | Page No.: 64-71







ABSTRACT
Using α-bungarotoxin (α -Bgtx) and β -bungarotoxin (β -Bgtx) to capture their corresponding proteins from preparation of crude human muscle receptor (triton extract), antibodies against these corresponding proteins were detected in Myasthenia gravis patients. α-Bgtx binds to AChR and β -Bgtx binds to presynaptic membrane receptor (PsmR) of triton extract. The captured proteins were used as antigen in indirect ELISA to detect the IgG antibodies against AChR and PsmR in sera from 146 Indian patients with MG and in 30 controls. Out of 146, 130 were generalised myasthenic patients (GM), 16 ocular Myasthenic patients (OMG). In addition to AChR antibodies, Myasthenia Gravis (MG) patients were also showing antibodies against PsmR. The frequency of anti-AChR antibodies was 71% and anti-PsmR antibodies was 65%. The OD values of GMG for AChR and PsmR were (0.76 ± 0.34) and (0.75 ±0.37), for OMG patients (0.32 ± 0.1) and (0.72 ± 0.42) and for healthy controls (0.16 ±0.06) and (0.18 ±0.04), respectively. These results showed that MG is not only due to the damage of the postsynaptic membrane, but it could concurrently be a consequence of antibody mediated damage of the presynaptic membrane, resulting particularly the damage of its β-Bgtx binding sites and could play an important role in the pathogenesis of MG. Therefore, it is also necessary to detect antibodies against PsmR in addition to AChR antibody.
PDF Abstract XML References Citation
How to cite this article
Suhail Rasool, Madhuri Behari, M. Irshad, Vinay Goyal and B.L. Jailkhani, 2008. Antibodies Against Postsynaptic Acetylcholine Receptor and Presynaptic Membrane Receptor in Myasthenia Gravis. Trends in Medical Research, 3: 64-71.
URL: https://scialert.net/abstract/?doi=tmr.2008.64.71
URL: https://scialert.net/abstract/?doi=tmr.2008.64.71
INTRODUCTION
Myasthenia Gravis (MG) is an autoimmune disease that results from antibody mediated damage of Acetylcholine receptor (AChR) at the neuromuscular junction (Patrick and Lindstrom, 1973). Recently autoantibodies against the nAChR alpha or beta subunits were isolated from four MG sera by affinity chromatography on columns carrying immobilized recombinant extracellular domains of human nAChR expressed in the yeast Pichia pastoris (Sideris et al., 2007). Much evidence has been presented supporting this hypothesis, including the induction of Experimental Autoimmune MG (EAMG) by immunization with AChR (Patrick and Lindstrom, 1973). Anti-AChR antibodies acting on the AChR may thereby cause the abnormal muscular fatigue and other signs of MG (Lindstrom et al., 1976; Lefvert, 1978). The autoimmune character of MG and pathogenic role of AChR antibodies have been established by several observations e.g., the demonstration of anti-AChR antibodies in about 90% of MG patients (Lindstrom, 1977; Brenner et al., 1978; Dwyer, 1992; Vincent et al., 2003), passive transfer of disease with IgG of MG patient to mouse (Toyka et al.,1977), localization of immune complexes (IgG and complement) on the postsynaptic membrane. Antibodies against acetylcholine receptor (AChR) can be detected in most patients with Myasthenia gravis and are known to be involved in the immunopathogenesis of this disease. It has been demonstrated that patients with MG have also antibodies against second protein named presynaptic receptor, which is identified by utilizing β-Bgtx, a ligand which binds to presynaptic receptor (Link, 1992). The antibodies against pre-synaptic membrane receptor (PsmR) and acetylcholine receptor (AChR) in serum samples of Myasthenia Gravis (MG) patients and healthy donors were tested by enzyme-linked immunosorbent assays (ELISA) (Xu et al., 2003). Presynaptic receptor (β-Bgtx binding proteins) represents another antigen besides AChR relevant for development of MG.
Presynaptic receptor can be defined as receptors at or near the nerve terminal that can positively or negatively modulate transmitter release directly or indirectly influence the probability of an action potential resulting in exocytosis (Wonnacott, 1997). It has been earlier shown that antibodies directed against β-Bgtx binding protein occur in MG (Xiao et al., 1991). This protein named presynaptic receptor has been isolated from electroplax tissue of Torpedo californica (Qiao et al., 1994), bovine diaphragm muscle (Link, 1992) and fetal bovine diaphragm muscle (Chaun-Zhen Lu et al., 1991) by utilizing β-Bgtx. In the presynaptic terminal, voltage-gated potassium channels and voltage-gated calcium channels are subtly altered in genetic disorders and mutations in the enzyme that synthesises acetylcholine have been demonstrated in a particular form of hereditary myasthenia syndrome (McConville and Vincent, 2002).
The present research is aimed to detect the antibody against presynaptic membrane receptor (PsmR) in addition to postynaptic AChRs in patients of Myasthenia Gravis by using triton extract of human cadaver skeletal muscle as a source of antigen.
MATERIALS AND METHODS
Alpha bungarotoxin (α-Bgtx), Beta bungarotoxin (β-Bgtx), tween-20, orthophenylenediamine, Protease inhibitors were from Sigma Chemical Co., St. Louis; microtitration ELISA plates from Costar Cambridge, UK and antihuman IgG from Dakopats, Denmark. Other reagents were of analytical grade. Sera from 130 patients of generalized myasthenia, 16 of Ocular myasthenia gravis and 30 healthy controls were stored at -20°C, until use.
Receptor Solubiliztaion Cadaver skeletal muscle and liver was obtained from our hospital mortuary. The receptors (postsynaptic acetylcholine receptor and presynaptic membrane receptor) were extracted by the method of Stephenson et al. (1981) and Jailkhani et al. (1986). Briefly the tissue was homogenised in four volumes of chilled Homogenising Buffer (HB) (0.01 Phosphate buffer, pH 7.4) containing 1 M NaCl, 0.02% NaN3, 0.001 M EDTA, 0.1 M benzethonium chloride 0.002 M benzemidene hydrochloride and 0.0001 M phenylmethyl sulphonyl fluoride and 0.5 mg mL-1 bacitracin). The homogenate was centrifuged at 20,000 g for 60 min at 4°C. The pellet obtained at 20,000 g centrifugation was extracted for 3 h at 4°C in two volumes of HB containing 2% (v/v) triton X-100. The supernatant (triton extract) obtained on centrifugation at 20,000 g for 60 min at 4°C was finally filtered through glass wool, aliquoted and stored as solubilised receptor (Triton Extract). The triton extracts from human cadaver skeletal muscle was used as a source for AChR and PsmR.
ELISA for Postsynaptic and Presynaptic Receptor Antibodies
Anti nAChR Antibody (Post Synaptic) In the indirect assay as described by Jailkhani et al. (1986) the receptor was bound by adding tissue triton extracts diluted in PBS containing 0.05% tween-20 and 0.1% skimmed milk powder on 96 Well ELISA plates (Costar Cambridge, UK) previously coated with α-bungarotoxin (1.0 μg mL-1) at pH 9.6 for 16-18 h at 4°C. The reaction with receptor extract was performed at 37°C for 2 h. Following receptor adsorption, the plates were washed 5 times with PBST and wells reacted with suitable dilution of myasthenic or control sera for 2 h at RT. After washing (x5) with PBST, the plates were reacted for 2 h with 1:10,000 dilution of monospecific rabbit anti-human IgG conjugated to HRP (Dako A/S of Denmark). The substrate o-phenyldiamine (OPD) (Sigma) 40 mg/100 mL of substrate buffer (0.1 Citric acid 0.2 M Sodium hydrogen phosphate pH 5.0, containing 40 μL of 30% H2O2) was added for 30 min and the reaction stopped with 2.5 N H2SO4. Absorbance at 492 nm was read in an ELISA reader (Biorad).
Anti PsmR Antibody (Pre Synaptic Membrane Receptor) The PsmR was captured using β-bungarotoxin. All other steps were same as for AChR ELISA described earlier.
RESULTS
Immunoreactivity Profile of Human Cadaver Skeletal Muscle with Myasthenic Sera Fresh skeletal muscle was obtained from amputation and used as a source of receptors (AChR and PsmR). Using indirect ELISA (Jailkhani et al., 1986), the receptors in detergent solubilize extract (triton extract) was added on ELISA plates precoated with α and β-Bgtx. It was then incubated with pooled myasthenic sera. As evident from Fig. 1, the triton extract gave a concentration dependent immunoreactivity with MG pooled sera, while no immunoreactivity was seen with control sera pool. Fresh tissue from human is not found to be ethical or feasible to obtain, therefore, the use of cadaver tissues was preferred which were obtained from the morturary of AIIMS.
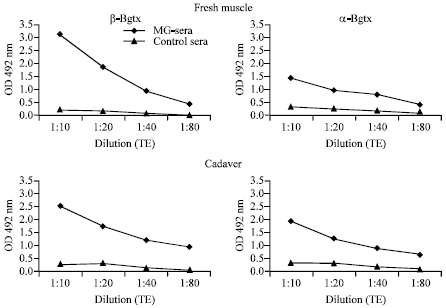 | |
| Fig. 1: | Immunoreactivity profile of triton extract of fresh muscle and different human cadaver muscle. Indicated dilutions of triton extracts from fresh and cadaver skeletal muscle were reacted with α-Bgtx and β-Bgtx pre coated plates for 2 h at 37°C, to trap the postsynaptic (α-Bgtx binding proteins) and presynaptic (β-Bgtx binding proteins) receptors which were then sequentially reacted with pooled myasthenic sera/control sera (1:200), anti IgG-HRP conjugate and substrate (OPD), the OD obtained at 492 nm at each input triton extracts dilutions are shown in figure |
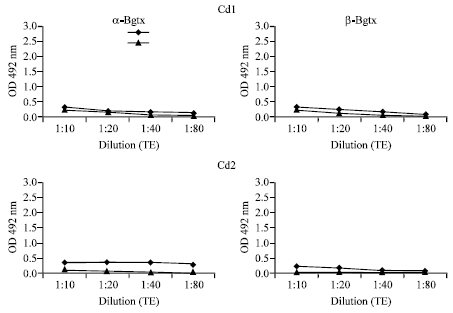 | |
| Fig. 2: | Immunoreactivity profile of triton extract of different human cadaver liver. Indicated dilutions of triton extracts from different human liver were reacted with α-Bgtx and β-Bgtx pre coated plates for 2 h at 37°C, to trap the postsynaptic (α-Bgtx binding proteins) and presynaptic (β-Bgtx binding proteins) receptors, which were then sequentially reacted with pooled myasthenic sera/control sera (1:200), anti IgG-HRP conjugate and substrate (OPD) the OD obtained at 492 nm at each input triton extracts dilutions are shown in figure |
It is evident by Fig. 1 that cadaver skeletal muscle gave comparable immunoreactivity with MG pool sera same as given by the fresh skeletal muscle, thereby justifying its use as a source for the AChR and PsmR. Since the concentration of the receptors varies from one sample to another, dilutions instead of protein concentrations were used for representing the immunoreactivity in triton extracts. Liver triton extracts showed no immunoreactivity with myasthenic sera (Fig. 2), which indicated that liver can been used as negative control.
Specificity In order to assess specificity of α and β-Bgtx for capturing AChR and PsmR an indirect ELISA was performed. Both α and β-Bgtx were coated on the ELISA Plates and incubated at 4°C for 16 h. Biologically active receptor (triton extract of human muscle) was incubated with increasing concentrations of toxin (both α and β-Bgtx) at 4°C overnight. After adding on ELISA plates, it was seen that triton extract with increasing concentration of α-Bgtx effectively competes with α-toxin pre-coated on the ELISA plate, while β-Bgtx added with the triton extract did not show any competition. Similarly β-Bgtx added with the triton extract competes with the previously coated β-Bgtx but not with α-Bgtx (Fig. 3).
Stability of AChR and PsmR at -20°C Cadaver tissue obtained up to 24 h postmortem do contain both AChR and PsmR, which retain both biological activity i.e., ability to bind their respective toxins and immunoreactivity i.e., vis-a-vis reactivity with Myasthenic sera. The reactivity in the Triton Extract (TE) remains stable when stored at -20°C in aliquotes for up to 65 days (Fig. 4).
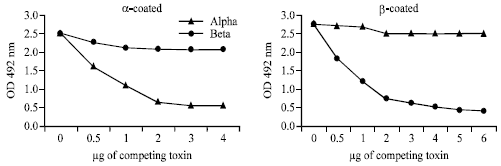 | |
| Fig. 3: | Specificity of α and β-Bgtx. α-Bgtx and β-Bgtx were coated (1 μg mL-1) on the ELISA plates in coating buffer (pH 9.6) overnight for 16-18 h at 4°C. The antigen (human muscle triton extract) was overnight incubated with increasing concentrations of α-Bgtx and β-Bgtx at 4°C. Plates were washed with PBST 5 times. Overnight incubated antigen with increasing concentration of toxin was added and put at 37°C for 2 h, followed by 5 washings with PBST. The test sera suitably diluted in PBST containing 0.1% skimmed milk were added and allowed to react for 2 h at RT. Following 5 washings with PBST, appropriate dilution of anti-IgG HRP conjugate was added for 2 h at 37°C. After 5 washings 0.2 mL of substrate O-phenylene diamine was added for 30 min at RT. The reaction was stopped with 50 μL of 2.5 N H2SO4 and the absorbance was taken at 492 nm in ELISA reader |
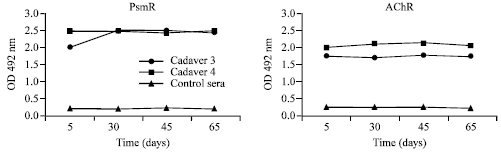 | |
| Fig. 4: | Stability of triton extract of human muscle at -20°C. Triton extracts were prepared from cadaver (No. 5) and stored and aliquoted at -20°C. The effect of immunoreactivity was assessed by indirect ELISA through α-Bgtx and β-Bgtx using a dilution of 1:10 of TE |
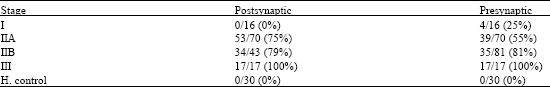
IgG Class of Antibodies Against Postsynaptic and Presynaptic Receptors in GMG, OMG and Healthy Controls One hundred and forty six sera from confirmed MG patients, out of which 130 were from Generalized Myasthenia Gravis (GMG), 16 from Ocular Myasthenia Gravis (OMG) and 30 from healthy controls were screened by indirect ELISA for IgG class of antibodies to postsynaptic nAChR (α-Bgtx binding proteins) and Presynaptic membrane receptor (β-Bgtx binding) captured using α-Bgtx and β-Bgtx as shown in Fig. 5. Scattergrams, which gives an overview of the antibody levels in confirmed myasthenic patients along with different stages and in healthy controls for AChR and PsmR, respectively. An OD value of more than 0.5 was taken as positive. The IgG class of antibodies against postsynaptic nAChR were present in 88/130 in GMG (67% positivity), 0/16 in OMG and in healthy controls no antibody was found. The IgG class of antibody against PsmR were present in 83/130 in GMG (63% positivity), 4/16 in OMG (25% positivity) and in healthy controls very low titre of IgG class of antibody were found (Table 1).
| Table 1: | Percentage of positivity in MG patients along with different stages |
 | |
.
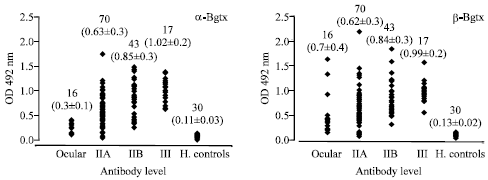 | |
| Fig. 5: | IgG class of antibodies against postsynaptic AChRs and presynaptic receptor along with different stages. Biologically active receptor (human muscle triton extract) was added to α-Bgtx and β-Bgtx a precoated plates and was incubated at 37°C for 2 h. The sera of different stages of myasthenic patents was used for screening for the IgG class of antibodies |
DISCUSSION
β-Bgtx is a 23 kD presynaptic neurotoxin that possess phospholipase A2 activity and bids to a high affinity site on the synaptosomal plasma membrane. The β-Bgtx induced depolarization of synaptosomal plasma membrane is Ca2+ dependent (Rugolo et al., 1986). In the present study we utilized two neurotoxins (α and β-Bgtx) to capture their specific proteins using triton extract of human cadaver muscle a source of antigen for both AChR and PsmR. Antibodies against α-Bgtx binding protein represents anti-AChR antibodies, while antibodies against β-Bgtx binding protein stands for anti-PsmR antibodies. The ELISA procedure described by Jailkhani et al. (1986) was adopted in our study. The specificity test was performed as from Fig. 3. When α-Bgtx was coated on to the ELISA plates, the α-Bgtx incubated with the antigen showed a consistent decrease in ELISA OD. Thereby proving that the incubated toxin effectively competes with toxin coated on the ELISA plate for the antigen whereas no such effect is seen with β-Bgtx. When β-Bgtx was coated and competed with α and β-Bgtx, only β-Bgtx showed competition. Hence indicating that α and β-Bgtx are specific for the AChR and PsmR respectively and don`t show any significant cross-reactivity. Autoantibodies against the nAChR alpha or beta subunits were isolated from four MG sera by affinity chromatography (Sideris et al., 2007).
Antibodies in Myasthenia Gravis The pathogenesis of MG involves the antibodies against postsynaptic AChR. The prevalence of anti-AChR antibodies in MG is influenced by a number of variables including variation of assay system used, cut off for reference range (we take >=0.5 as positive) and differences related to race. The frequency of anti-AchR antibodies was 71% and anti-PsmR antibodies was 65% (Table 2). We have screened 146 MG patients and 30 healthy controls for IgG class of antibodies against AChR and PsmR. However there are 15 MG patients who had only anti-PsmR antibodies and 12 patients had only anti-AChR antibodies. Seventy six patients had both anti-PsmR and anti-AChR antibodies. None of the 30 healthy controls was positive for these antibodies.
| Table 2: | Frequencies of antibodies against AChR and presynaptic membrane receptor in MG subgrouped according to Osserman scale |
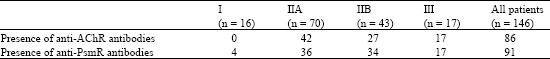 | |
AchR are not only distributed at the postsynaptic neuromuscular junction, but also at presynaptic membrane as well (Wilson, 1982). Presynaptic receptor has been confirmed by electrophysiological and pharmacological methods to belong to a subclass of voltage gated K+ channels (Rehn and Betz, 1984; Schmidt and Betz, 1988). It has also been demonstrated that antibodies (IgG) are produced against AChR in paralleled to PsmR. In addition to that we have seen that α-Bgtx is specific for postsynaptic AChR whereas β-Bgtx is specific for presynaptic membrane receptor and there is no cross reactivity using human cadaver muscle as source of antigen. Human cadaver liver was used as a negative control which does contain AChR or PsmR. Therefore, it is found that Myasthenia gravis occurs not only due to formation of antibodies against postsynaptic AChRs, but also against PsmR. We consider that in MG, primary pathogenic involvement is antibodies against presynaptic membrane receptor, which induces an initial damage to presynaptic membrane (Chaun Zen Lu et al., 1991). On the other hand, anti-AChR antibodies might also have a negative feedback on presynaptic membrane (Takamori et al., 1986).
In conclusion, MG is not only due to the damage of the postsynaptic membrane, but it could concurrently be a consequence of antibody mediated damage of the presynaptic membrane, resulting particularly the damage of its β-Bgtx binding sites and could play an important role in the pathogenesis of MG. It is also necessary to detect antibodies against PsmR in addition to AChR in myasthenia gravis and correlate it with the disease stages.
REFERENCES
- Brenner, T., B. Zweiman and O. Abramsky, 1978. Radio-Immunoassay of antibodies to acetlycholine receptor in serum of myasthenia gravis patients. Isr. J. Med. Sci., 14: 986-989.
PubMed - Chaun-Zhen, L., L. Hans, M. Xu-An, X. Biao-Guo, Z. Yu-Lin and Q. Zhen, 1991. Anti-presynaptic membrane receptor antibodies in myasthenia gravis. J. Neurol. Sci., 102: 39-45.
CrossRef - Dwyer, J.M., 1992. Manipulating the immune system with immune globulin. N. Engl. J. Med., 326: 107-116.
CrossRefDirect Link - Jailkhani, B.L., D. Asthana, N.F. Jaffery, R. Kumar and G.K. Ahuja, 1986. A simplified ELISA for anti-receptor antibodies in Myasthenia Gravis. J. Immunol. Meth., 86: 115-118.
CrossRef - Lefvert, A.K., 1978. Determination of acetylcholine receptor antibody in myasthenia gravis: Clinical usefulness and pathogenesis implications. J. Neurol. Neurosurg. Psychiatry, 41: 394-403.
Direct Link - Lindstrom, J.M., M.E. Seybold, V.A. Lennon, S. Whittingham and D.D. Duane, 1976. Antibody to acetylcholine receptor in myasthenia gravis. Prevalence, clinical correlates and diagnostic value. Neurology, 26: 1054-1059.
PubMed - Lindstrom, J., 1977. An assay for antibodies to human acetylcholine receptor in serum from patients with myasthenia gravis. Clin. Immunol. Immunopathol., 7: 36-43.
CrossRefDirect Link - Link, H., 1992. Myasthenia gravis: T and B cell reactivities to the β-bungarotoxin binding protein presynaptic membrane receptor. J. Neurol. Sci., 109: 173-181.
CrossRefDirect Link - McConville, J. and V. Vincent, 2002. Diseases of the neuromuscular junction. Curr. Opin. Pharmacol., 2: 296-301.
Direct Link - Patrick, J. and J. Lindstrom, 1973. Autoimmune response to acetylcholine receptor. Science, 180: 871-872.
CrossRefDirect Link - Qiao, J., Z.Y. Wang and H. Link, 1994. β-Bungarotoxin binding protein is immunogenic but lacks myasthenogenicity in rats. J. Neurol. Sci., 121: 190-193.
CrossRefDirect Link - Rehm, H. and H. Betz, 1984. Solubilization and characterization of -Bgtx binding protein of chicken brain membrane. J. Biol. Chem., 259: 6865-6869.
PubMed - Rugolo, M., O.J. Dolly and Nicholis, 1986. The mechanism of action of β-bungarotoxin at the presynaptic plasma membrane. Biochem. J., 233: 519-523.
CrossRefDirect Link - Sideris, S., G. Lagoumintzis, G. Kordas, K. Kostelidou, A. Sotiriadis, K. Poulas and S.J. Tzartos, 2007. Isolation and functional characterization of anti-acetylcholine receptor subunit-specific autoantibodies from myasthenic patients: Receptor loss in cell culture. J. Neuroimmunol., 189: 111-117.
Direct Link - Takamori, M., S. Okumura and A. Yasuda, 1986. Presynaptic modification of neuromuscular transmission by antiacetylcholine receptor antibody. Neurology, 36: 942-947.
Direct Link - Toyka, K.V., D.B. Drachman, D.E. Griffin, A. Pestronk, J.A. Winkelstein, K.H. Fishbeck and I. Kao, 1977. Myasthenia gravis. Study of humoral immune mechanisms by passive transfer to mice. N. Engl. J. Med., 296: 125-131.
PubMed - Vincent, A., J. Bowen, J. Newsom-Davis and J. McConville, 2003. Seronegative generalized myasthenia gravis: Clinical features, antibodies and their targets. Lancet Neurol., 2: 99-106.
Direct Link - Wilson, D.F., 1982. Influence of presynaptic receptors on neuromuscular transmission in rat. Am. J. Physiol., 242: C366-C372.
PubMed - Wonnacott, S., 1997. Presynaptic nicotinic ACh receptors. Trends Neurosci., 20: 92-98.
CrossRefDirect Link - Xiao, B.G., C.Z. Lu, B. Höjeberg and H. Link, 1991. Immunological specificity and cross-reactivity of anti-acetylcholine receptor and anti-presynaptic membrane receptor antibodies in myasthenia gravis. J. Neurol. Sci., 105: 118-123.
Direct Link - Xu, T.H., J. Ding, Y.L. Shi, M.F. Li, C.Z. Lu and J. Qiao, 2003. Effects of myasthenia gravis patients' sera with different autoantibodies on slow K+ current at mouse motor nerve terminals. Neurol. Res., 25: 58-62.
Direct Link