
S. Issaka
National Institute for Agronomic research of Niger (INRAN), BP. 60 Kollo Niger
A. Onasanya
Plant Pathology Unit, Africa Rice Center (AfricaRice), P.O. Box 33581, Dares-Salaam, Tanzania
A. Basso
National Institute for Agronomic research of Niger (INRAN), BP. 60 Kollo Niger
F. Sorho
University of Cocody-Abidjan, 22 BP. 582, Abidjan 22, Cote d�Ivoire
A. Haougui
National Institute for Agronomic research of Niger (INRAN), BP. 60 Kollo Niger
A.Y. Sido
National Institute for Agronomic research of Niger (INRAN), BP. 60 Kollo Niger
S. Ake
University of Cocody-Abidjan, 22 BP. 582, Abidjan 22, Cote d�Ivoire
D. Fargette
IRD, BP 6450, 34394 Montpellier cedex 5, France
Y. Sere
Pathology Unit of Africa Rice Center (Africa Rice), 01 Bp. 2031 Cotonou, Benin
Trends in Applied Sciences Research
Year: 2012 | Volume: 7 | Issue: 1 | Page No.: 32-45







ABSTRACT
This study has been conducted in screen house with an aim to asses the Rice yellow mottle virus pathogenic diversity and the level of resistance of released varieties in Niger republic. Sixty RYMV isolates from 23 Niger rice perimeters were inoculated mechanically to nine rice cultivars. The disease symptoms were scored at 42 days after inoculation. Analysis of Variance (ANOVA) and Additive Main effect and Multiplicative Interaction (AMMI) analysis were performed on the percentage of severity. The reaction of the rice cultivars to the virus isolates was significantly different. The interaction between isolates and rice cultivars was also significant. AMMI cluster analysis revealed the existence of four major pathotypes (Path 1 to 4) of Rice Yellow Mottle Virus (RYMV) in Niger republic. Path 4 pathotype included 12 resistance breaking isolates (20%). Path 3 and Path 2 pathotypes consist of 15 and 26 isolates respectively and were typical of wild type isolates with moderate level of pathogeny, including none aggressive (path 3 = MP) and aggressive isolates (Path 2 = MPA). The fourth pathotype Path 1 was made of 7 isolates and typical of particular isolates which have a moderate pathogenic level (FP). Resistance Breaking (RB) isolates occupied 30% of Niger rice ecologies in variable proportion. The rice varieties (Bassiroumo, IR15-29-690-3-1 and Kassoumo) released in Niger were highly susceptible to RYMV and therefore constituted a favorable condition for the rice yellow mottle disease propagation. This information is useful in rice breeding programs in the development and deployment of RYMV resistant cultivars to different rice perimeters in Niger Republic.
PDF Abstract XML References Citation
Received: June 30, 2011;
Accepted: September 19, 2011;
Published: November 17, 2011
How to cite this article
S. Issaka, A. Onasanya, A. Basso, F. Sorho, A. Haougui, A.Y. Sido, S. Ake, D. Fargette and Y. Sere, 2012. Existence of Several Pathotypes among Rice Yellow Mottle Virus (RYMV) Isolates Collected in Niger Republic. Trends in Applied Sciences Research, 7: 32-45.
URL: https://scialert.net/abstract/?doi=tasr.2012.32.45
URL: https://scialert.net/abstract/?doi=tasr.2012.32.45
INTRODUCTION
Rice Yellow Mottle Virus (RYMV), genus Sobemovirus, is the major constraint to rice production in Africa (Kouassi et al., 2005), causing yield losses comprised between 17-100% (Onwughalu et al., 2011). The virus colonizes all Africa rice ecosystems (SÁrÁ et al., 2008). The host range was restricted to Gramineous species, mainly Oryzae and Eragrostida genus. The RYMV is mechanically transmissible by insects, genus Chrysomelidea (Abo et al., 2000; Nwilene et al., 2009). These insects play a major role in the virus transmission. An abiotic transmission (by soil, seedbed, cultural practices etc.) of the RYMV is also reported (Traore et al., 2008, 2006a). The disease is characterized by mottle and yellowing symptoms of varying intensities depending on genotype and time after infection. These symptoms could therefore be mistaken for iron or nitrogen deficiency (Onasanya et al., 2006; Gnanamanickam, 2009). Others symptoms observed on infected plants are pale yellow mottle leaves, stunted, reduced tillering, non-synchronous flowering, poor panicle exertion and grains discoloration (Gnanamanickam, 2009).
The Rice yellow mottle disease was first described in Niger Republic in 1986 by Reckhaus and Adamou (Kouassi et al., 2005). However, no significant research has been undertaken to understand the interaction between RMYV strains and rice varieties in the country. In other African countries, major progress had been made in knowledge of the virus population’s structure and diversity (N’Guessan et al., 2000; Fargette et al., 2004; Sorho et al., 2005; Onasanya et al., 2006; Rakotomalala et al., 2008). Several strains of the virus with different geographical distribution were described (Abubakar et al., 2003; Traore et al., 2005). Natural and transgenic sources of resistance were reported (Sorho et al., 2005) and two (2) to three (3) different pathogroups of rymv1 had been defined in different rice spaces (N’Guessan et al., 2001; Onasanya et al., 2006; Amancho et al., 2009). Moreover many resistance genes were described. The major gene of resistance against rymv, rymv1, identified in the O. sativa resistant variety Gigante. The gene rymv1 encodes a translation initiation factor called eIF (iso) 4G (Albar et al., 2006). This factor is also responsible of the resistance of resistant cultivars O. glaberrima, accessions Tog5681 and Tog5672 linked respectively to the alleles rymv1-3 and rymv1-4. These 2 alleles are distinct from the Gigante resistance allele rymv1-2 (Albar et al., 2006). A new resistance gene was recently reported as RYMV2 in Tog5672 and Tog5691 (Thiemele et al., 2010). However, the emergence of Resistance-breaking (RB) isolates (Fargette et al., 2002; Traore et al., 2006b; Amancho et al., 2009) is a matter of concern. The existence of this kind of RYMV isolate has been proved in most rice ecologies (Ochola and Tusiime, 2011). Therefore the deployment of resistant varieties in a rice growing environment requires a good knowledge of the virus diversity.
The current study investigated the existence and distribution of different pathotype of Rice yellow mottle virus isolates in Niger republic. A preliminary propagation of the collected samples (Unpublished) revealed the existence of different types of isolates including some RB isolates. The current work aimed to clarify the effective presence and proportion of RB isolates in Niger RYMV’s populations. Their implications in rice improvement in Niger are also discussed.
MATERIALS AND METHODS
Rice cultivars: Nine rice cultivars (Table 1) were used for this study.
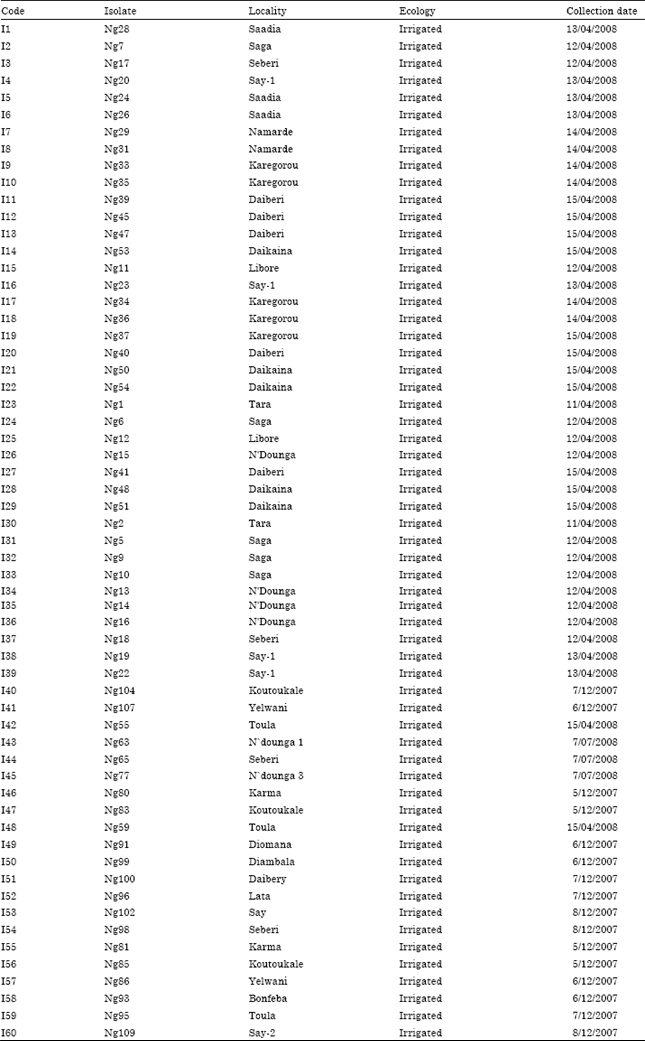
Virus isolates and inoculation procedure: Sixty RYMV isolates (Table 2) used for this study were collected from rice and weeds in 23 different localities in Niger republic. Before use, each isolate was first propagated in the susceptible rice variety IR 64, by mechanical inoculation of 21 old plants in the screen house. Four weeks after inoculation, leaves from each RYMV isolate with typical symptoms of RYMV disease were harvested and ground with 0.01 M phosphate buffer pH 7.0 at the ratio of 1:10 (w/v). By this way, the inoculum of the isolates was standardised. The resulting homogenate filtered through cheesecloth.
| Table 1: | Identity of varieties used for this study |
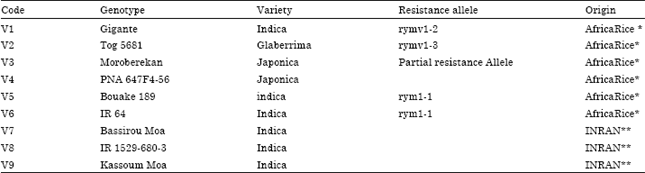 | |
| * Varieties obtained from Plant Pathology Unit, Africa Rice Center. ** Varieties obtained from National Institute of Agricultural Research of Niger (INRAN) | |
| Table 2: | List of isolates used for the study |
 | |
Carborundum powder (600 mesh) was added to the inoculum to aid the penetration of the virus into leaf tissues. The 21 days old seedlings of the 9 rice cultivars were mechanically inoculated in the screen house with the inoculum of the 60 isolates.
Experimental design: For each isolate, the experimental design was a randomized complete block with 3 replications. A fourth replicate was inoculated with 0.01 M phosphate buffer pH 7.0 and used as control. All the young rice plants were inoculated by the same way.
Data collection: Each leave was scored 42 days after inoculation according to a standard evaluation system where 1 characterized a lack of symptom and 9 a destroyed leave (Sorho et al., 2005) Disease severity was then evaluated as :
S = {(n1*1 + n3*3 + n5*5 + n7*7 + n9*9)*100} / {(n1 + n3 + n5 + n7 + n9)*9}
where, n1, n3, n5, n7 and n9 represented the number of plants scored 1, 3, 5, 7 and 9, respectively (Fininsa, 2003).
Data analysis: Analysis of Variance (ANOVA) and mean comparison of percentage disease severity were performed, using IRRISTAT version 4.3 statistical software. Genotype (cultivar) by environment (isolate) interaction effects on percentage disease incidence was carried out using additive main effect and Multiplicative Interaction (AMMI) analysis (Ebdon and Gauch, 2002b; Onasanya et al., 2006).
RESULTS
The ANOVA performed on disease severity indicated a significant difference between the 60 RYMV isolates collected in Niger Republic and the nine rice varieties (Table 3). Moreover, a significant interaction was found between the varieties and the isolates.
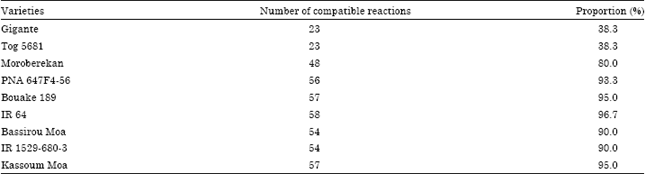
The mean comparison (full data not showed) allowed describing the interaction between the RYMV isolates and the rice varieties. An average severity of 25% is a threshold under which the response reflected the non-pathogenicity of the isolates and the resistance of the varieties. An example of a qualitative description of the relationship between virus isolates and rice varieties was given in the Table 4a and b. The mean comparison (Table 4a) allowed identifying the severity value that was statistically under or above the threshold of 25%. Therefore, the rice cultivars were described either Resistant (R) or Susceptible (S) to the RYMV isolate (Table 4b). A highly compatible interaction was found on almost all susceptible and partial resistant genotypes, as the majority of isolates (80%) were pathogenic on them. Then the resistant varieties showed a fairly compatible interaction (38.3%) with only few isolates (Table 5).
Cultivars Gigante and Tog 5681 are characterized by their resistance to more RYMV strains than the other varieties (Fig. 1; Table 5).
| Table 3: | ANOVA for Severity (SEV), at 42 days after inoculation |
 | |
| *HS = Highly significant at 1% | |
| Table 4a: | Example of a qualitative description of the relationship between RYMV isolates and rice varieties 4a: Mean comparison |
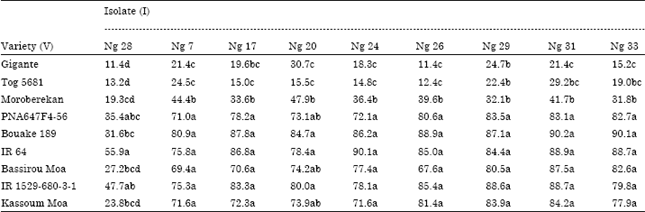 | |
| In the same column, means followed by the same letter are not significantly different at 5% level | |
| Table 4b: | Qualitative description |
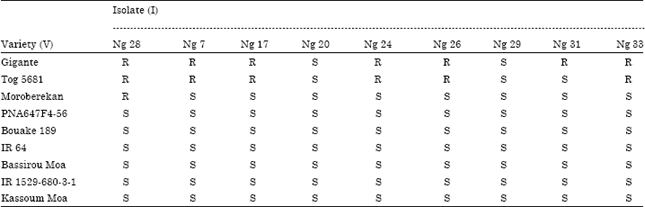 | |
| R: resistant; S: susceptible | |
| Table 5: | Compatible reaction between 9 rice genotypes and 60 Niger RYMV isolates |
 | |
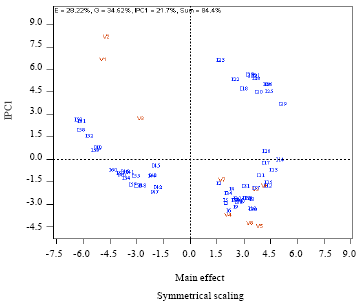 | |
| Fig. 1: | Main effects genotype (cultivar) by environment (isolate) interaction on percentage RYMV disease severity using additive main effects and multiplicative interaction (AMMI) analysis V1: Gigante, V2: Tog5681, V3: Moroberekan, V4: PNA647F4-56, V5: Bouake 189, V6: IR64, V7: IR1529-680-1, V8: Bassiroumo, V9: Kassoumo, I1, I2……I60: Isolate 1, 2….60 (Completed Table 2) |
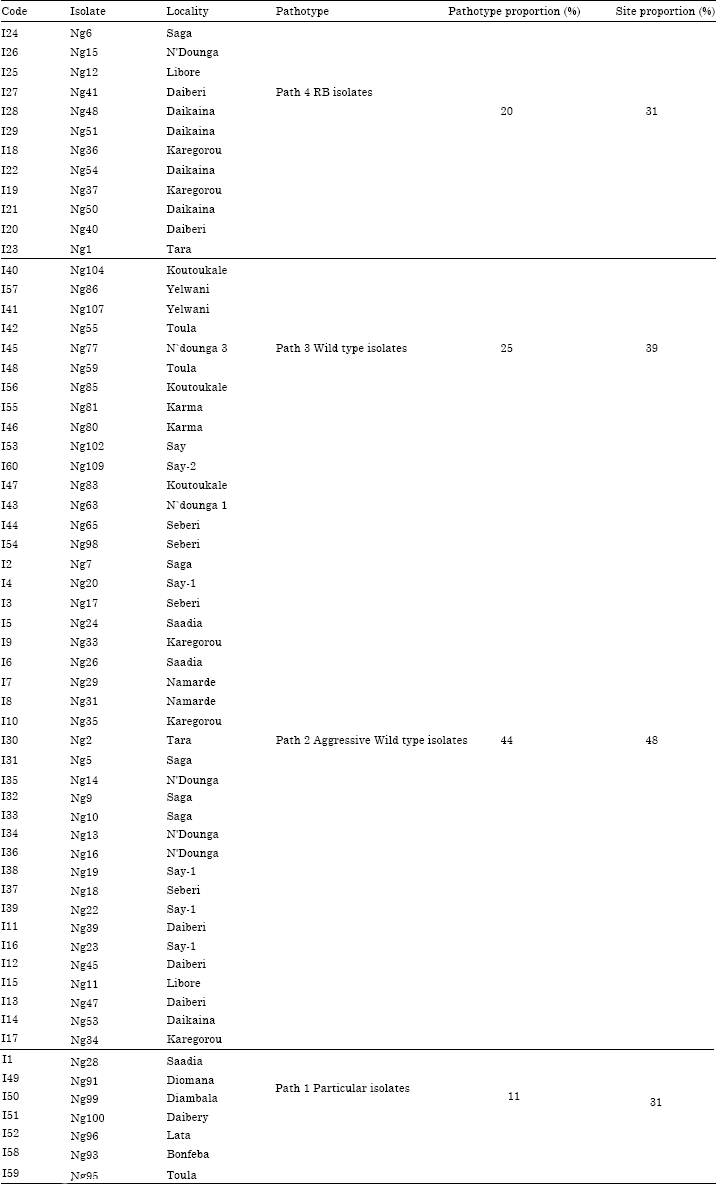
Indeed, their resistance alleles (rymv1-2 and rymv1-3, respectively) are broken by 38.3% of the Niger RYMV isolates (Table 5). However, according to the principal component analysis and pathotypes distribution (Fig. 1 and Table 6), the resistance breaking isolates (RB)’s proportion in Niger rice ecologies did not exceed 20%. Moreover, some isolates showed a particular pathogenicity but represented only 8% of the pathogen population (Table 6).
| Table 6: | Distribution of RYMV pathotypes in the irrigated rice ecologies of Niger Republic revealed by AMMI |
 | |
Genotypes x Environment interactions (environment represented by various isolates) were examined through the AMMI model. This model combines the additive model of ANOVA and the interaction displayed by the principal component analysis. This analysis (Fig. 2) showed that severity is more correlated with PC1 axis (52.8%), showing the aggressiveness of RYMV isolates and the susceptibility of rice cultivars. PC2 axis (25.3%) expressed the virulence of RYMV isolates and the resistance level of rice varieties. Finally, 4 types of interaction could be described (Fig. 2).
The AMMI cluster analysis (Fig. 3) classified the RYMV isolates in four major pathotypes, Path 1 to Path 4 (Table 6). Moreover the rice varieties were categorized into three groups (Fig. 4). Path 4 pathotype was made of 12 RYMV isolates with typical virulent isolates (20%) called resistance breaking isolates (HP). Path 3 and Path 2 pathotypes consisted of 15 and 26 RYMV isolates respectively and were typical of wild type isolates with moderate level of pathogeny including non- aggressive (path 3 = MP) and aggressive isolates (path 2 = MPA). The fourth pathotype was made of 7 RYMV isolates and was typical of particular isolates with average pathogenic level (FP). RB isolates occupied 30% of irrigated perimeter of Niger in variable proportions according to locations. In some perimeters like Daibery, Daikaina and Kareygorou, they coexisted with wild type isolates characterized by variable pathogenic level (Table 6). According to AMMI analysis, Path 2 pathotype isolates were responsible mainly for favourable interactive conditions, leading to significant increase in percentage disease severity in all the rice cultivars used (Fig. 2).
Variety resistant group (R) was made up of two highly resistant cultivars (V1 = Gigante and V2 = Tog 5681) and one moderately resistant cultivar (V3 = Moroberekan). Two susceptible cultivars (V4 = PNA 647F4-56 and V5 = Bouake 189) made up the S group while HS group included four highly susceptible cultivars (V6 = IR 64, V7 = Bassiroumo, V8 = IR15-29-690- 3-1 and V9= Kassomumo). These results have shown that the three widely cultivated genotypes in Niger republic (Bassiroumo, IR15-29-690-3-1 and Kassomumo) were highly susceptible to RYMV.
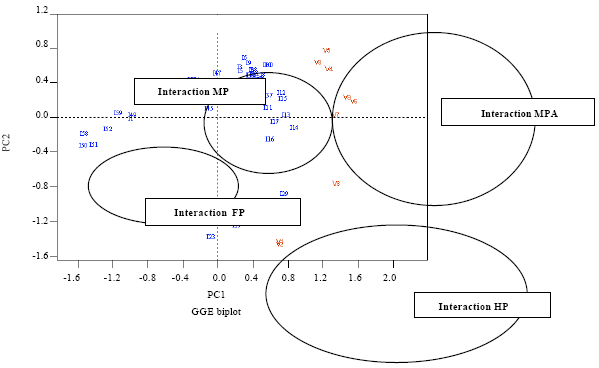 | |
| Fig. 2: | Relationship among rice variety and RYMV isolate as revealed by genotype (cultivar) by environment (isolates) interaction effects on percentage RYMV disease severity using additive main effects and Multiplicate Interaction (AMMI) analysis. HP: Highly pathogenic (virulent); MP: Moderately pathogenic; MPA: Aggressive and moderately pathogenic and FP: Fairly pathogenic |
DISCUSSION
This study revealed the pathogenic diversity of RYMV population in Niger. The existence of interaction indicated that varieties were different by their vertical and complete resistance (Adugna, 2004) and therefore, the isolates by their virulence.
This diversity can be described both by the isolates ability to develop compatible reaction (virulence) with the rice cultivars and the intensity of disease (aggressiveness). The four pathotypes revealed by statistical analysis consist of one resistance breaking isolates group (Path 4), two wild types isolates groups with variable aggressiveness level (Path 3 and Path 2) and a particular pathogenic isolate group (Path 1). In earlier studies, the existence of 2 to 3 pathotypes in different rice environments of Africa has been observed (N’Guessan et al., 2001; Ndjiondjop et al., 2001; Onasanya et al., 2006). The present results reported, for the first time, the presence of RB isolates (20%) in Niger republic. They are coexisting with a high proportion of wild type isolates non RB but aggressive. Such a high proportion of avirulent RYMV isolates in Niger rice ecology has already been reported elsewhere in Africa (Sorho et al., 2005; Onasanya et al., 2006; Amancho et al., 2009; Ochola and Tusiime, 2011). This information underlines the risk of a control of RYMV based only on the use of resistant genes.
The additive main effect and Multiplicative Interaction (AMMI) analysis appeared to be very effective in understanding and explaining complex genotype by environment (GxE) interactions between the rice genotypes and RYMV isolates (Ebdon and Gauch, 2002a; Onasanya et al., 2004, 2006). Previous studies revealed that such interactions could generate complex data difficult to understand with ordinary analysis of variance (Onasanya et al., 2004, 2006).
 | |
| Fig. 3: | RYMV isolate pathotype as revealed by Additive Main effects and Multiplicate Interaction (AMMI) cluster analysis. Path 1: particular isolates pathotype, Path 2: aggressive wild isolates pathotype, Path 3: Wild type isolates pathotype and Path 4 = resistance breaking isolates pathotype |
In the current study, the 60 RYMV isolates used covered major rice ecologies from different localities in Niger republic leading to very high RYMV interactions among rice cultivars. The existence of four pathotypes obtained in this study (Path 1 to Path 4) has led to differential reactions among genotypes with heavy implications on the cultivar resistance stability (Onasanya et al., 2004, 2006; N’Guessan et al., 2000). The presence of different Niger RYMV pathotypes in all rice areas indicated a high RYMV pathogenic variability in fields. Infestation of more than 30% perimeters by RB isolates suggested their wide distribution.
The cultivars Gigante and Tog 5681 has showed fair compatibility with Niger RYMV strain while the other cultivars present high compatibility. In fact, the resistance genes rymv1-2 (of Gigante) and rym1-3 (of Tog 5681) were useful against several RYMV pathotypes (Albar et al., 2006; Rakotomalala et al., 2008; Thiemele et al., 2010). These resistance sources are exploitable for varietal control of Rice yellow mottle disease (Ndjiondjop et al., 2001; Sorho et al., 2005).
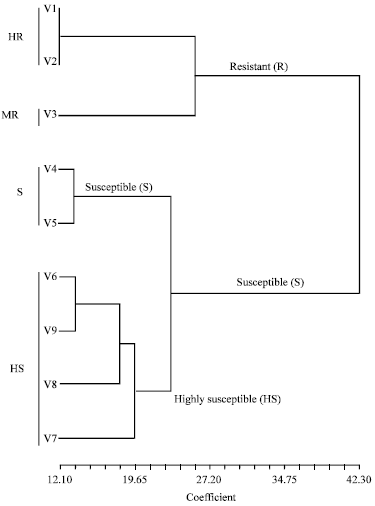 | |
| Fig. 4: | Resistance status of nine rice varieties to RYMV isolates from Niger Republic as revealed by Additive Main effects and Multiplicate Interaction (AMMI) cluster analysis. MR: moderately resistant; HRHighly resistant; S: Susceptible; HS: Highly susceptible |
However, the presence of a new virulent pathotype (Path 4) in different rice ecologies of Niger Republic constitutes major obstacles for rice cultivar resistance stability. Indeed, this pathotype can overcome the high resistance of the two resistance allele rymv1-2 and rymv1-3. The rate of RB prevalence (20%) obtained in the present study is relatively similar to the prevalence rates previously observed in South and Central Africa. These studies have reported 16.4% (Traore et al., 2006b) and 20% (Allarangaye et al., 2006) of symptomatic Resistance Breaking isolates. The high resistance of rice cultivars was first overcome by wild types isolates (Konate et al., 1997). Then their emergence was confirmed in laboratory studies, during serial inoculations of wild type avirulent isolate to resistant genotypes (Fargette et al., 2002). Besides, the occurrence of RYMV RB isolates is reported in african rice fields with contrasted ecologies (Traore et al., 2006b; Rakotomalala et al., 2008).
With the current RYMV isolates pathogenic population structure in Niger republic and the high susceptibility of cultivars widely cultivated, proportion of wild type RYMV aggressive isolates can increase in the future. There is a probability that Path 3 and Path 2 pathotype prevalence become high, if Niger’s farmers continue to use susceptible varieties. Path 4 pathotype isolates can also increase during its possible interactions with other pathotypes. The possible interactions between different pathogenic RYMV strains could lead to emergence of new virulent strains (Pinel-Galzi et al., 2009). The use of highly resistant cultivars (Gigante and Tog5681) will potentially reduce Path 3, Path 2 and Path 1 pathotypes population and their interactions. There is a probability that the highly resistant cultivars (Gigante and Tog5681) will survive through combinations of gene present in the cultivars population. Indeed, the population resistance is enhanced by genes polymorphism that may result in short-term selection of more tolerant cultivars in stressful viral environments (Ebdon and Gauch, 2002a; Onasanya et al., 2006). This information could be useful in rice breeding programs in the development and deployment of RYMV resistant cultivars in different rice perimeters in Niger Republic.
The compatible relation among rice cultivars and RYMV strains is due to gene to gene interaction between rice eIF(iso) 4G protein coded by gene rymv1 and RYMV viral protein genomic (VPg) (Albar et al., 2003; Hebrard et al., 2008). The resistance breakdown is associated to mutations of certain amino acid (48, 52 etc.) of the RYMV’s VPg (Hebrard et al., 2006; Pinel-Galzi et al., 2007; Poulicard et al., 2010; Traore et al., 2010). Thus, the VPg of RYMV RB isolates identified here must be sequenced to assess the molecular basis of their genetic and pathogenic determinism. Last, the presence of particular isolates (Path 1 isolates) which affect less susceptible cultivars indicate either genotype tolerance or negative interaction with the isolates. The susceptibility of these cultivars against ELISA negative RYMV isolates was also reported, indicating that the biological diagnosis is more sensitive than serological tests (Traore et al., 2008). Molecular studies on the VPg of Path 1 pathotype isolates could help to understand the determinism of their particular pathogeny.
CONCLUSION
The high genotypes by environment interactions in the reactions of rice cultivars to RYMV revealed the existence of four pathotypes of RYMV and three cultivar groups in Niger republic. A new RYMV pathotype (Path 4) able to break down the highly rice resistance is now reported in this country. Two rice cultivars (Gigante and Tog5681) identified in this study possess heterogeneous viral resistance characteristics that made them more stable, adaptable and resistant to stress induced by RYMV pathotypes from different localities. This information is useful in rice breeding programs for the development and deployment of RYMV resistant cultivars in different rice ecologies and localities in Niger Republic and over all sub-tropical Africa countries. As the impact of the inoculum in resistance breaking mechanism is preponderant, it is critical to support the deployment of resistant cultivars with sanitation measures. These measures involving isolation of nurseries and removal of infected weeds and rice ratoons could reduce the risk of resistance breaking isolates emergence.
ACKNOWLEDGMENTS
The International Foundation of Science (IFS) and the government of Japan had supported financially the researches. We thank them for their contribution. We thank also M. George SOTON, support staff at AfricaRice, for his technical aid.
REFERENCES
- Abubakar, Z., F. Ali, A. Pinel, O. Traore and P. N'Guessan et al., 2003. Phylogeography of rice yellow mottle virus in Africa. J. Gen. Virol., 84: 733-743.
CrossRefDirect Link - Albar, L., M. Ndjiondjop, Z. Esshak, A. Berger and A. Pinel et al., 2003. Fine genetic mapping of a gene required for rice yellow mottle virus cell-to-cell movement. Theor. Applied Genet., 107: 371-378.
PubMed - Albar, L., M. Angratz-Reyser, E. Hebrard, M.N. Ndjiondjop, M. Jone and Ghesquiere, 2006. Mutations in the eIF(iso)4G translation initiation factor confer high resistance of rice to rice yellow mottle virus. Plant J., 47: 417-426.
CrossRef - Allarangaye, M.D., O. Traore, E.V.S. Traore, R.J. Millogo and G. Konate, 2006. Evidence of non-transmission of rice yellow mottle virus through seeds of wild host species. J. Plant Pathol., 88: 309-315.
Direct Link - Ebdon, J.S. and H.G. Gauch, 2002. Additive main effect and multiplicative interaction analysis of National Turfgrass performance trials. Crop Sci., 42: 489-496.
CrossRefDirect Link - Ebdon, J.S. and H.G. Gauch, 2002. Additive main effect and multiplicative interaction analysis of national turfgrass performance trials: II. Cultivar recommendations. Crop Sci., 42: 497-506.
Direct Link - Fargette, D., A. Pinel, Z. Abubakar, O.Z. Traore and C. Brugidou et al., 2004. Inferring the evolutionary history of rice yellow mottle virus from genomic, phylogenetic and phylogeographic studies. J. Virol., 78: 3252-3261.
CrossRefDirect Link - Fininsa, C., 2003. Relationship between common bacterial blight severity and bean yield loss in pure stand and bean-maize intercropping systems. Int. J. Pest Manage., 49: 177-185.
Direct Link - Hebrard, E., A. Pinel-Galzi, A. Bersoult, C. Sire and D. Fargette, 2006. Emergence of a resistance-breaking isolate of rice yellow mottle virus during serial inoculations is due to a single substitution in the genome-linked viral protein Vpg. J. Gen. Virol., 87: 1369-1373.
CrossRef - Konate, G., O. Traore and M. Coulibaly, 1997. Characterization of rice yellow mottle virus isolates in Sudano-Sahalian areas. Arch. Virol., 142: 1117-1124.
CrossRefDirect Link - Kouassi, N.K., P. N'Guessan, L. Albar, C.M. Fauquet and C. Brugidou, 2005. Distribution and characterization of Rice yellow mottle virus: A threat to African farmers. Plant Dis., 89: 124-132.
CrossRefDirect Link - N'Guessan, P., A. Pinel, M. Caruana, R. Frutos, A. Sy, A. Ghesquiere and D. Fargette, 2000. Evidence of the presence of two serotypes of rice yellow mottle Sobemovirus in Cote d'Ivoire. Eur. J. Plant Pathol., 106: 167-178.
CrossRefDirect Link - N'Guessan, P., A. Pinel, A.A. Sy, A. Ghesquiere and D. Fargette, 2001. Distribution, pathogenicity and interactions of two strains of Rice yellow mottle virus in forested and savannah zones of West Africa. Plant Dis., 85: 59-64.
Direct Link - Ndjiondjop, M.N., C. Brugidou, Z. Shipping, D. Fargette, A. Ghesquire and C.M. Fauquet, 2001. High resistance to Rice yellow mottle virus in two cultivated rice cultivars is correlated with the failure of cell-to-cell movement. Physiol. Mol. Plant Pathol., 59: 309-316.
CrossRef - Nwilene, F.E., A.K. Traore, A.N. Asidi, Y. Sere, A. Onasanya and M.E. Abo, 2009. New records of insect vectors of Rice Yellow Mottle Virus (RYMV) in Cote d'Ivoire, West Africa. J. Entomol., 6: 189-197.
CrossRefDirect Link - Ochola, D. and G. Tusiime, 2011. Pathogenicity of rice yellow mottle virus and the potential sources of resistance against the disease in Eastern Uganda. Asian J. Plant Pathol., 5: 1-15.
CrossRef - Onasanya, A., Y. Sere, F. Nwilene, M.E. Abo and K. Akator, 2004. Reactions and resistance status of differential rice genotypes to Rice yellow mottle virus, genus Sobemovirus in Cote d'Ivoire. Asian J. Plant Sci., 3: 718-723.
CrossRefDirect Link - Onasanya, A., Y. Sere, M. Sie, K. Akator, M. M. Coulibaly and A. Hamadoun, 2006. Existence of two pathotypes of rice yellow mottle virus, genus Sobemovirus, in Mali. Plant Pathol. J., 5: 368-372.
CrossRefDirect Link - Pinel-Galzi, A., M. Rakotomalala, E. Sangu, F. Sorho and Z. Kanyeka et al., 2007. Theme and variations in the evolutionary pathways to virulence of an RNA plant virus species. PLoS Pathogen, 3: e180-e180.
Direct Link - Pinel-Galzi, A., A. Mpunami, E. Sangu, M. Rakotomalala and O. Traore et al., 2009. Recombination, selection and clock-like evolution of rice yellow mottle virus. Virology, 394: 164-172.
CrossRefDirect Link - Sere, Y., A. Onasanya, F.E. Nwilene, M.E. Abo and K. Akator, 2008. Potential of insect vector screening method for development of durable resistant cultivars to rice yellow mottle virus disease. Int. J. Virol., 4: 41-47.
CrossRefDirect Link - Sorho, F., A. Pinel, O. Traore, A. Bersoult and A. Guesquiere et al., 2005. Durability of natural and transgenic resistances in rice to rice yellow mottle virus. Eur. J. Plant Pathol., 112: 349-359.
CrossRefDirect Link - Traore, M.D., V.S.E. Traore, A. Galzi-Pinel, D. Fargette, G. Konate, A.S. Traore and O. Traore, 2008. Abiotic transmission of Rice yellow mottle virus through soil and contact between plants. Pak. J. Biol. Sci., 11: 900-904.
CrossRefPubMedDirect Link - Traore, O., F. Sorho, A. Pinel, Z. Abubakar and O. Banwo et al., 2005. Processes of diversification and dispersion of rice yellow mottle virus inferred from large-scale and high-resolution phylogeographical studies. Mol. Ecol., 14: 2097-2110.
CrossRefDirect Link - Traore, O., M. Traore D. Fargette, and G. Konate, 2006. Rice seedbeds as a source of primary infection by rice yellow mottle virus. Eur. J. Plant Pathol., 115: 181-186.
Direct Link - Traore, O., A. Pinel, E. Hebrard, M.Y.D. Gumedzoe, D. Fargette, A.S. Traore and G. Konate, 2006. Occurrence of resistance-breaking isolates of rice yellow mottle virus in West and central Africa. Plant Dis., 90: 259-263.
Direct Link - Adugna, A., 2004. Alternate approaches in deploying genes for disease resistance in crop plants. Asian J. Plant Sci., 3: 618-623.
CrossRefDirect Link - Onwughalu, J.T., M.E. Abo, J.K. Okoro, A. Onasanya and Y. Sere, 2011. Rice yellow mottle virus infection and reproductive losses in rice (Oryza sativa Linn.). Trends Applied Sci. Res., 6: 182-189.
CrossRef - Rakotomalala, M., A. Pinel-Galzi, L. Albar, A. Ghesquiere, Y. Rabenantoandro, P. Ramavovololona and D. Fargette, 2008. Resistance to rice yellow mottle virus in rice germplasm in Madagascar. Eur. J. Plant Pathol., 122: 277-286.
CrossRef - Fargette, D., A. Pinel, H. Halimi, C. Brugidou, C. Fauquet and M. van Regenmortel, 2002. Comparison of molecular and immunological typing of the isolates of rice yellow mottle virus. Arch. Virol., 147: 583-596.
CrossRefDirect Link