
Hui-Yu Liu
College of Geography Science, Nanjing Normal University, Nanjing 210097, Peoples Republic of China
Zhen-Shan Lin
Jiangsu Key Laboratory of Environmental Change and Ecological Construction, Nanjing 210097, Peoples Republic of China
Hong-Yu Liu
Northeast Institutes of Geography and Agricultural Ecology,
CAS, Changchun 130012, Peoples Republic of China
Trends in Applied Sciences Research
Year: 2006 | Volume: 1 | Issue: 4 | Page No.: 341-349







ABSTRACT
The scaling of environmental capacity of single species (K) with the number of patches (P) has been introduced by applying scaling theory in this paper based on the investigated data of Oriental White Storks in Naoli River basin in China. The maximum capacity of Oriental White Storks scales the number of patches to power 0.71, i.e., Ka P0.71. In the meantime, by incorporating the scaling relation into non-autonomous population model for single species with Allee effect under the accumulative effects of human activities, we simulated the population dynamics of the Oriental White Stork. The simulation results show that the Oriental White Stork is a living dead species; there is about 100 year time debt for its extinction to respond to the past habitat loss and fragmentation. To avoid the extinction of Oriental White Storks, habitat quality must be improved instead of holding current quality. By comparisons of different cases of improving habitat, we find that the changes of average patch area have more significant effects on the persistence of Oriental White Storks than the changes of the number of patches on condition that the changes of total areas are equal. Decreasing the number of patches is favorable to the persistence of the Oriental White Stork. At the same time, a larger reserve is more propitious to the Oriental White Stork`s long-term persistence than several smaller reserves with the same area.
PDF Abstract XML
How to cite this article
Hui-Yu Liu, Zhen-Shan Lin and Hong-Yu Liu, 2006. Response of Single Species with Allee Effect to Accumulative Effects of Human Activities and its Forecast a Case Study of the Oriental White Stork. Trends in Applied Sciences Research, 1: 341-349.
URL: https://scialert.net/abstract/?doi=tasr.2006.341.349
URL: https://scialert.net/abstract/?doi=tasr.2006.341.349
Instroduction
Habitat destruction involving downright loss of habitat, degrading habitat quality and fragmentation of the remaining habitat is the leading cause of population and species extinction (Hanski, 1998). Fragmented habitat affects the environmental capacity of a certain species mainly through average patch area and the number of patches. So the study of the scaling relation among environmental capacity, average patch area and the number of patches is very important in landscape and population dynamics (Lin et al., 2005). Direct observations and experiments are hardly sufficient to decipher all the long-term and large-scale biological consequences of habitat loss and fragmentation, the dynamical simulation is an effective tool to study the population dynamics. The previous models to simulate population dynamics of single species do not refer to the accumulative effects of human activities and mostly are autonomous dynamical models. Lin et al. (2005) establish Non-autonomous Population Dynamics Model for Single Species Affected by Human activities at the first time by incorporating the scaling of environmental capacity with the number of patches and take a case study of red-crowned cranes. They pointed out: the red crowned crane is a living dead species. The question we will propose is whether the Oriental White Stork meets the same question. The Oriental White Stork is also listed as the national first-grade wildlife of China for protection on China's List of Wildlife Under State Important Protection (Zheng and Wang, 1998) and classified as threatened on the IUCN Red list of Threatened Animals (WCMC, 1994). The population size decrease quickly since 1970’s due to habitat deterioration following marsh reclamation for cultivation and disafforestation and overkill.
In this paper, we firstly identified the scaling of environmental capacity of the Oriental White Stork to the number of patches; secondly, we forecast the population dynamics of the Oriental White Stork by Non-autonomous Population Dynamics Model for Single Species Affected by Human activities. We intend to reveal the mechanism of the response of population dynamics of Oriental White Storks to accumulative effects of human activities and to answer whether Oriental White Stork is a living dead species by numerical simulations; finally, we have simulated different cases of improving habitat quality, which provide some scientific implication to preserve the long-term persistence of Oriental White Stork.
MATERIALS AND METHODS
Study Area
The study was conducted in Naoli River basin (131°31'2E, 45°43'N-134°10'E, 47°35'N), where is the hinterland of Three River Plain in Heilongjiang Province of China. It borders on the watershed of Wanda Mountain in southeast and adjoins Wusuli River on east with a total area of 241.67x104 hm2, which is close to 1/4 of the total area of Three River Plain and the total wetland area of it is more than 1/4 of that of Three River Plain. There are four wetland nature reserves in Naoli River basin and all of which have some representativeness in preserving the biodiversity of Three River Plain and holding the ecosystem function and process. The agricultural exploitation is so intense in this basin that there are four great-scale agricultural developments since 1949, especially for the period of rapid economic growth since 1980.This study area mainly consists of six counties and seven modern farms, and has 1.25 millions of people, of which peasant population account for about 65.6%.This area is one of the most important productive bases of commodity grains in China. Many wetlands change into farmland due to the population explosion and economy growth. Wetlands shrank to shore and lose 80%. The remained wetland only account for 7.1% of total area of the wetland in this basin and11% in Tree River Plain.
Data Source
Two data sets are adopted to analyze the landscape changes of the wetland ecosystem in 1967 and 2000, respectively (Fig. 1).The dataset in 1967 is derived from the relief map scaling 1:200,000 in 1960s (referencing to the relief map scaling 1:100,000 in 1967 and 1968), While that in 2000 is derived from ETM sensor data acquired from June to July in 2000(four panoramic images and five semi-panoramic images) and detailed field data (Liu et al., 2004a).
| Table 1: | The changes of the number of Grus japonensis, habitat area and patches number in Naoli River basin |
| Table 2: | The extinction time when total area is not changed with r = 0.065 and a = 10 |
 | |
| Table 3: | The extinction time when total area increasing with r = 0.065 and a = 10 |
 | |
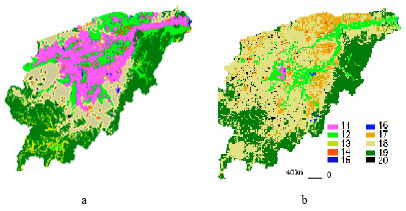 | |
| Fig. 1: | The landscape structure of wetlands at 1967 (a) and 2000 (b) in Naoli River. 11. Marsh, 12. meadow wetland ,13 .scrub wetland, 14. island forest wetland, 15. lake wetland, 16. river ,17. paddy field ,18. dry land ,19. forest, 20. other land |
To improve the precision of above data and to reflect the status of land use and wetland landscape during different periods in history, we adopt the same system of landscape classification, map projection and minimum cell of cartography.
According to the natural properties and seeper condition, the wetland landscape has been classified into widen water surface (also sorted into lake, rive, paddy field), meadow wetland, scrub wetland, island forest wetl and and marsh wetland and the rest has been classified into dry land, forest and other land.
Observation Data
Table 1 shows the changes of the number of Oriental White Storks, habitat area and the number of patches in Naoli River basin in China from 1967 to 1999 (Zeng et al., 2003; Liu 2004).
As shown from Fig. 1 and Table 1, habitat loss and fragmentation in Naoli River basin have been intensified as a result of population explosion and reinforced activities of human being since 1967, which results in the population size of Oriental White Storks decreasing rapidly. Therefore, it is propitious to the conservation of Oriental White Storks to study the relations among the environmental capacity, average patch area and the number of patches.
Non-autonomous Population Dynamics Model for Single Species with Allee effect under accumulative effects of Human Activities
The scaling relation between environmental capacity and the number of patches
It is well known that for any species or population, its environmental capacity has a close relation with average patch area and the number of patches. Assuming s and P denoting average patch area and the number of patches respectively, then the relations among environmental capacity (k, i.e., the number of Oriental White Storks habitat can hold), average patch area and the number of patches can be expressed as:
| (1) |
If we take S0, K0 as the total area of all patches and environmental capacity at the starting time and ST, kT as the total area and environmental capacity after human have destroyed landscape for T year (T = 33 from 1967 to 2000 in this study) and then
| (2) |
Based on Eq. (2), we get the following:
| (3) |
From Eq. (3), we get the index of the number of patches (n):
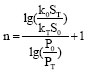 | (4) |
In this paper, we substitute corresponding data in Table1 into Eq. (4), then
| (5) |
Therefore, the scaling of environmental capacity of Oriental White Storks with the number of patches can be expressed as
| (6) |
From Eq. (3) and (5), the environmental capacity of Oriental White Storks at any time can be expressed as:
| (7) |
Non-autonomous Population Dynamics Model for Single Species under Accumulative Effects of Human Activities
| (8) |
Equation 8 is the well known Logistic growth model. Where r is the intrinsic growth rate, N is the population size and K is the maximum capacity that environment and resources can hold.
Obviously the right-hand side of Eq. (8) does not include time explicitly andthe model belongs to autonomous Population Dynamics Models. At the meantime, other early models as Prey-Predator model (Lotka, 1925; Nicholson and Bailey, 1935) and cotemporary model as Levin’s (1969), Hanski’s (1997), Tilman’s (1994) and so forth, they also do not include time explicitly. Therefore, they can not be used to study the accumulative effects of human activities. Human-caused habitat destruction and its effects on population dynamics are different during different periods in history. That is, the interaction of population dynamics and human-caused habitat destruction is a function of time. So, we think it is needed to build and apply non-autonomous, Population Dynamics Model to describe and study the time accumulative effects of human activities. We introduce f(t) to represent the effects of human-caused habitat destruction on population dynamics.
| (9) |
where t is the time for the simulation of population dynamics after human activities stop. After human-caused habitat destruction stops, the effects of human activities on population fade down over time. Then the accumulative effects of human activities on environmental capacity can be expressed as:
| (10) |
If kT represents the environment capacity when human activities stop and K(t) represent the environment capacity in consideration of accumulative effects of human activities, then we can get the following:
| (11) |
We substitute Eq. 11 and 7 into Eq. 12, then we can get the non-autonomous population dynamics model for single species under the accumulative effects of human activities:
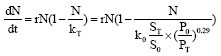 | (12) |
The right-hand of model (12) we proposed includes time explicitly; it belongs to non-autonomous dynamical systems in mathematics. Herein, it can be used to study the time accumulative effects of human activities.
Allee effects
Populations at very low numbers may go extinct due to Allee effects (Allee, 1931). Too low population size will lead to low fecundity (McCarthy et al., 1997), when population density is under the threshold density, population will decline and go extinct at end. Because the Oriental White Stork is an endangered species, it is more prone to Allee effect. So it is necessary to introduce Allee effect into model (12).
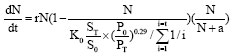 | (13) |
where N/N+a is the Allee effect and a is the Allee effect constant. The bigger a is, the greater the Allee effect is. We take 9≤a≤35 for the Oriental White Stork, i.e., when the number of Oriental White Storks ranges between 9 and 35, species will go extinct.
In contrast to the classical Logistic Model, the model we have advanced here involves not only time t, but also the accumulative effects of human activities, the landscape fragmentation factor which imposes the most significance effects on population dynamics as well as the habitat area factor. Moreover, it also holds the merits of Logistic Model that sufficiently consider both the environmental capacity and the Allee effect.
RESULTS
In order to study the future trend of the Oriental White Stork and further to provide scientific applications to protect the Oriental White Stork, we have simulated population dynamics of the Oriental White Stork under the accumulative effects of human activities since 1967. Figure 2 is the simulation results of population dynamics of the Oriental White Stork according to model (13) with r = 0.065 (Xu et al., 1993), a = 10(a), a = 20(b).
As shown from Fig. 2a, when a = 10, population size will increase transitorily after human activities stop andthen decrease quickly. During the first decades, the declining speed is very high and becomes slowly later, which indicates the effects of human activities become more and more weak. In 2097, the population density is lower than 10, which means the Oriental white stork is doomed to extinction. Shown from Fig. 2b, when a = 20, the Oriental white stork also experiences a transient recovery firstly andthen decline.
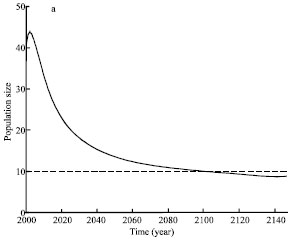 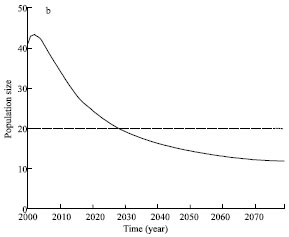 | |
| Fig. 2: | The population dynamic of Oriental white storks in the case of a = 10(a), a = 20(b), r = 0.065 |
It will go extinct in 2029.At the meantime, shown from Fig. 2, if not recovering and improving the present habitat quality, the Oriental white stork will be doomed to extinction in this area due to the past 30 year continuous habitat destruction caused by human activities. That is to say, there is a time delay of 100 year for species extinction to respond to habitat deterioration. The simulation results also justify the extinction debt advanced by Tilman et al. (1994), i.e., following habitat loss, the threshold condition for survival is no longer met for some species, but these species have not yet gone extinct because of the time delay in their response to environmental change. Such species are called the dead-living (Hanski, 1997).To protect Oriental White Stork, we must improve habitat quality sufficiently before species go extinct.
DISCUSSION
From above simulation, we can get the following conclusions:
| • | The maximum capacity of Oriental White Storks scales the number of patches to power 0.71, i.e., K ∝ P0.71. |
| • | If not recover and improve the landscape quality, the Oriental white stork will go extinct in 100 year in this area, that is, the Oriental white stork is a living dead species. It provides some implications for biology conservation that we must recover and improve habitat quality so as to avoid extinction of Oriental white storks instead of solely protecting the present landscape. |
How can we improve habitat quality and further to protect the Oriental white stork on the premise that the total area of habitat being unchanged? If we improve habitat quality, how can we protect the Oriental white stork from extinction more effectively? To solve such question, we discuss how we conserve the Oriental white stork on condition that the total area is not changed and increases respectively. Then, we designed 4 cases and respectively give the extinction time in these cases as shown in Table 2 with r = 0.065 and a = 10.
Shown from Table 2, on condition that the total area is not changed with r = 0.065 and a = 10, to reduce the number of patches (i.e., to increase average patch area) is propitious to species persistence. When decreasing 50% of the number of patches, the extinction time will be postponed for 88 year in contrast to 2097 in current landscape; when decreasing 90% of the number of patches, the Oriental white stork can escape extinction. However, to reduce the average patch area (i.e., to increase the number of patches) is disadvantage to species persistence. When decreasing 50 and 90% of average patch area, the extinction time will be advanced for 34 and 65 year, respectively. Moreover, we can find that the effects of decreasing the number of patches are more significant than those of increasing the number of patches. Therefore, a bigger reserve is more favorable to persistence than several smaller ones provided that the total areas are the same.
The above mentioned is solely to improve the structure of current landscape. If we would optimize landscape by means of increasing habitat area, which effect will be more prominent, to increase average patch area or to increase the number of patches? We suppose the situations that average patch area doesn’t change and the number of patches increases, or average patch area increases and the number of patches doesn’t change. Then, we designed 4 cases and respectively give the extinction time in these cases as shown in Table 3 with r = 0.065 and a = 10.
As shown from Table 3, we suppose that habitat area increases by 50%, in case 1 the extinction time of the Oriental White Stork is in 2269, being postponed for 172 year in contrast to 2097 in current landscape with average patch area unchanged and the number of patches increasing by 50%; in case 2 the Oriental White Stork will die out in 2499, being postponed for 402 year. In case 3 and case 4, total areas both increase by 100%. The extinction time is in 2849 year in case 3, being postponed for 748 year.While in case 4, the Oriental White Stork will escape from extinction. It also reveals that it is more propitious to species persistence to increase average patch area than to increase the number of patches when increasing the same proportion of habitat.
As shown above, on condition that the changes of total habitat area are equal, the effect of the change of average patch area on extinction time is more significant than the change of the number of patches. Therefore, when recovering and improving the habitat to avoid Oriental White Stork extinction, we should give first priority to the enlargement of the average patch area or to bigger patches instead of increasing the number of patches.
Liu et al. ( 2004b) find that linear corridors are the major factor leading to the fragmentation of the wetland landscape in Naoli River basin, especially for the construction of irrigation works. The number of penstocks increases from 1234 to 13546. By comparing different cases on the conditions of improving habitat quality; we find that average patch area has a more significant influence on species than the number of patches when the changes of total areas are the same. In addition, the Oriental White Stork requires very big domain for the propagation (Su, 1993). So to preserve the Oriental White Storks, it is very important to reduce the number of penstocks to make several smaller patches change into bigger patches. Moreover, as for the design of the reserve of Oriental White Storks, the larger the reserve area is, the more beneficial it is to species persistence. A bigger reserve is better than several smaller ones though the total areas are equal, which agrees with some principles of biogeography island theory (Diamond, 1975).
Although nowadays many complex spatial explicit metapopulation models have been developed to forecast the population dynamics, with a common shortage that they are too complicated with lots of parameters difficult to estimate and assumptions difficult to test. While the model we have put forward hereinto is much easier to operate and test. Similarly with other models, there are lots of uncertainties in forecasting the extinction risk due to the variability of landscape structure. In short, the contribution of the model to the biology conservation lies in the comparison between different landscape structures and different management cases (Hanski, 1998).
ACKNOWLEDGMENTS
This research has been supported by China NSF (No. 40371108), China National “211” Key Project: The Environmental evolution and Ecological Construction on Multi-spatial-temporal Scales andthe Foundation of Jiangsu Key Laboratory of Environmental Change and Ecological Construction. We also thank the Anonymous Reviewers for their helpful comments.
REFERENCES
- Lin, Z.S., H.Y. Liu and H.Y. Liu, 2005. Non-autonomous population dynamics model for single species with Allee effect affected by human activities and its application: A case study of red-crowned cranes Grus japonensis. Acta Ecol. Sin., 25: 945-951.
Direct Link - Liu, H.Y., X.G. Lv and S.K. Zhang, 2004. Landscape biodiversity of wetlands and their changes in 50 years in watersheds of the Sanjiang Plain. Acta Ecol. Sin., 24: 1472-1479.
PubMed - Liu, H.Y., S.K. Zhang and X.G. Lv, 2004. Wetland landscape structure and the spatial-temporal changes in 50 years in the Sanjiang Plain. Acta Geogr. Sin., 59: 391-400.
Direct Link - McCarthy, M.A., D.B. Lindenmayer and M. Dreschler, 1997. Extinction debts and risks faced by abundant species. Conserv. Biol., 11: 221-226.
CrossRefDirect Link - Tilman, D., R.M. May, C.L. Lehman and M.A. Nowak, 1994. Habitat destruction and the extinction debt. Nature, 371: 65-66.
Direct Link - Zeng, Z.W., L. Chen and X.M. Li, 2003. The numerical distribution and conservation of oriental white stork in China. Territory Nat. Resour. Study, 25: 71-72.
Direct Link