
C.J. Ogugbue
School of Industrial Technology, Environmental Technology Unit, Universiti Sains Malaysia Penang, Malaysia
F. Akubuenyi
Department of Biological Sciences, Cross River University of Technology, Cross River State, Nigeria
A.A. Ibiene
Department of Microbiology, University of Port Harcourt, Port Harcourt, Nigeria
Singapore Journal of Scientific Research
Year: 2012 | Volume: 2 | Issue: 1 | Page No.: 1-13







ABSTRACT
The decolourization of Acid orange 10 in Synthetic Saline Wastewater (SSW) medium under static conditions was investigated using Bacillus firmus. The bacterium was able to effectively decolourize the azo dye in a wide range of salt (NaCl) concentrations (1 to 6%, w/v), temperature (20-40°C) and pH (6-10) after 24 h of incubation in static culture. Colour removal by this strain was enhanced in the presence of starch and considerable decolourization was obtained in medium with high salt concentration (6%) in the presence of SDS (0.5-50 mg L-1). Results suggest the potential of this halotolerant bacterium for use in effective treatment of dye-contaminated wastewaters containing high salt concentrations characteristic of typical textile effluents.
PDF Abstract XML References Citation
Received: November 04, 2011;
Accepted: March 01, 2012;
Published: June 25, 2012
How to cite this article
C.J. Ogugbue, F. Akubuenyi and A.A. Ibiene, 2012. Bacterial Decolourization of Acid Orange 10 in Synthetic Wastewater under Saline Conditions: Effect of Process Parameters. Singapore Journal of Scientific Research, 2: 1-13.
URL: https://scialert.net/abstract/?doi=sjsres.2012.1.13
URL: https://scialert.net/abstract/?doi=sjsres.2012.1.13
INTRODUCTION
Azo dyes are the class of dyes most widely used in textile finishing and paper-printing industries having a world market share of 60-70%. They are usually recalcitrant to conventional wastewater treatment (Kodam et al., 2005) and have become of concern because of their color, bio-recalcitrance and potential toxicity to animals and humans (Dubey et al., 2007; Yoo et al., 2000). Consequently, it has become necessary to search for and develop effective treatments and technologies for the decolourization of azo dyes in textile effluents. Currently, the implementation of physical/chemical methods (Igwe et al., 2007; Tchatchueng et al., 2009; Yasar et al., 2007) have the inherent drawbacks of being economically unfeasible (as they require more energy and chemicals), unable to completely remove the recalcitrant azo dyes and/or their organic metabolites, generating a significant amount of sludge that may cause secondary pollution problems and involving complicated procedures (Forgacs et al., 2004; Zhang et al., 2004). Thus, more studies are now focused on the use of biological methods for wastewater treatment (Omar, 2008; Rajendran et al., 2011a, b; Rajeswari et al., 2011), since it is an eco-friendly, cost-competitive alternative to chemical decomposition process that could help reduce water consumption compared to physicochemical treatment methods (Rai et al., 2005; Verma and Madamwar, 2003).
Textile wastewaters bearing azo dyes are characterized by their deep color and high salt concentration due to the presence of dyes and high concentrations (40 to 100 g L-1) of salts (especially sodium chloride) used in dye baths to ensure maximum fixation of dye to fibres (Carliell et al., 1994, 1998; EPA, 1997; Manu and Chauhari, 2003). It is estimated that during the mercerising process the weight of these salts can make up 20% of the fibre weight (EPA, 1997). Textile wastewaters are also usually slightly alkaline due to elevation of hydroxyl ions when sodium hydroxide is used in the dye bath to increase the pH to the alkaline range during fixation (Dos Santos et al., 2007). High salinities or wide salinity ranges in wastewater make dye degradation by microorganisms difficult and are, currently, limiting factors in the development of biotreatment methods due to the sensitivity of many of these bacteria to the high concentrations of salts that are released into the wastewater (Carliell et al., 1994; Manu and Chauhari, 2003).
The performance of the biological treatment process for saline wastewater usually has low colour removal due to adverse effects of salt on microbial flora (Kargi and Dincer, 1996). Generally, sodium chloride concentrations above 1% can cause moderate inhibition of most bacterial activities (De Baere et al., 1984; Guo et al., 2008) due to plasmolysis and/or recession of the cytoplasm which is induced by an osmotic difference across the cell wall. This causes an outward flow of intracellular water resulting in the loss of microbial activity and cell dehydration (Kargi, 2002; Ozalp et al., 2003). The implication of this is that degrading microorganisms will require a pretreatment of the effluent wastewater to dilute high salt concentrations before the biological treatment; there by generating even larger volumes of wastewater. Moreover, the use of physico-chemical processes such as reverse osmosis (El-Kady and E1-Shibini, 2001), ion exchange (Dincer and Kargi, 2000) or electro dialysis (Dincer and Kargi, 2000) for salt removal before biological treatment is rather expensive and of limited applicability.
Consequently, the identification and characterization of halotolerant and halophilic bacteria that can degrade azo dyes may facilitate the development of biological treatment methods for treatment of saline azo dye wastewater. Many microorganisms can thrive in saline environments. The potential for growth in different salt concentrations is a variable characteristic of microorganisms which greatly depends on their physiological properties. These properties have been used to classify different microorganisms. Nonhalophilic microorganisms are defined as organisms growing optimally below 0.2 M NaCl, while halophilic microorganisms need concentrations of more than 0.2 M NaCl to reach their optimum growth. In addition, bacteria able to grow in the absence of salt as well as in the presence of relatively high salt concentrations are designated as halotolerant (DasSarma and Arora, 2002).
Bioremediation in salty environments inevitably requires the application of these halotolerant and halophilic microorganisms which are able to grow under such harsh conditions. Halotolerant and halophilic bacteria have been utilized in the bioremediation of oil (Delille et al., 1998; Margesin and Schinner, 2001) and oxyanion pollution (Nriagu and Payna, 1988; Kinkle et al., 1994), but there is a dearth of information on their effectiveness in decolourization of solutions containing azo dyes and high concentrations of sodium chloride. The use of microorganisms able to degrade azo dyes in the presence of salt could help prevent costly dilution to lower the salinity, or the removal of salt by physico-chemical methods before biological treatment.
This study evaluates the biodegradation of Acid orange 10, an azo dye by a newly isolated halotolerant and pH versatile bacterium in media with sodium chloride concentrations ranging from 0 to 8%.
MATERIALS AND METHODS
Dye and chemicals: Acid orange 10 (C.I. 16230) was obtained from Huntsman Co. (formerly Ciba-Geigy). It was used as received without any further purification. Figure 1 shows the chemical structure of the dye.
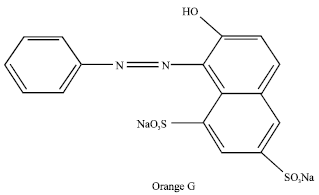 | |
| Fig. 1: | Chemical structure of acid orange 10 |
The dye was filter-sterilised on a 0.45 μm filter (Millipore, USA) prior to addition to sterile culture medium. All other reagents were of analytical grade and were purchased from Sigma chemical co. (USA).
Microorganism and culture medium: The bacterial strain used in this study was isolated from the sludge of a textile industrial treatment plant and was identified as Bacillus firmus as previously described (Ogugbue and Sawidis, 2011). The sludge was acclimatized in dye solution for eight weeks and then used for the isolation of the bacterium. The bacterium was able to decolorize a variety of azo dyes under anoxic conditions by azo bond reduction Determination of the halotolerance characteristics of the bacterial isolate was done by checking for growth in Synthetic Saline Wastewater (SSW) medium containing various salt concentrations (1-10%) and spiked with dye (100 mg L-1). The SSW prepared with de-mineralized water presented the following composition (g L-1); (NH4)2SO4 0.28, NH4Cl 0.23, KH2PO4 0.067, MgSO4.7H2O 0.04, CaCl2.2H2O 0.022, FeCl3.6H2O 0.005, NaCl 40.0, NaHCO3 1.0 and 1 mL L-1 of a trace element solution containing (g L-1); ZnSO4.7H2O 0.01, MnCl2.4H2O 0.1, CuSO4.5H2O 0.392, CoCl2.6H2O 0.248, NaB4O7.10H2O 0.177 and NiCl2.6H2O 0.02. The SSW medium was used for subsequent decolourization experiments.
Determination of the effect of different process parameters on dye biodecolourization under saline conditions: The effects of process parameters (salt concentration, dye concentration, pH, temperature, SDS concentration, additional carbon sources) on azo dye biodecolourization under saline conditions were investigated using batch static flask experiments. Different salt concentrations (1, 2, 4, 6 and 8%) were prepared using sodium chloride analytical grade as a source of salt in medium in order to check the salt tolerance potential of the bacterial isolate. Decolourization by the isolate was also investigated at different initial concentrations (5, 10, 50, 100, 500 mg L-1) of the dye in medium containing 6% salt concentration. A pH range of 4-11 was screened to determine the range of pH activity for the bacterium and temperature optimization was carried out at 15, 20, 25, 30, 35, 40 and 45°C. Thereafter, the effect of different concentrations (0.1, 0.5, 1.0, 5.0, 10 and 50 mg L-1) of Sodium Dodecyl Sulphate (SDS) on decolourization was investigated under best conditions of pH and temperature. The effect of the presence of additional carbon source (glucose, starch, cellulose and sodium carbonate) in the SSWM on the decolourization process was also determined. All experiments, except the effect of salt concentration, were carried out in medium containing 6% salt concentration in 24 h. During incubation, samples from each flask were taken at different times (0, 6, 12, 18 and 24 h) and analyzed for residual dye concentration. The same cell concentration of the isolate, adjusted at optical density 1.0 at λ = 620 nm (equivalent to ca. 4.87 log CFU mL-1), was used to decolourize all the dyes. Samples from each flask were used to determine the % color removal at the end of the experiment as described below. Control flasks consisting of cell-free medium (to indicate effect of medium components on decolourization) and culture medium poisoned with 1% HgCl2 (to show adsorption of dye on cells) were set up. All assays were conducted in triplicates.
Analytical methods: The UV/visible spectral signatures of the culture samples and control were determined using a scanning spectrophotometer (UV/visible, Shimadzu, Kyoto, Japan). A clarified sample from dye free medium was used as the baseline for absorbance calibration. Withdrawn samples were clarified before absorbance measurements in order to prevent absorbance interference from the cellular or other suspended debris. Sample clarification was carried out by centrifugation at 12,000 rpm for 10 min at 10°C and the absorbance of supernatants obtained determined at the maximum absorption wavelength (λmax) of the dye (475 nm). The calibration curve for absorbance versus dye concentration, obtained by plotting the corresponding maximum absorbance in the UV-visible spectra at different concentrations of dye, was used to obtain the residual dye concentration in each sample using the absorbance reading obtained.
The % color removal was calculated using the following equation:
| (1) |
where, C0 and C1 represent the initial and final concentrations of azo dye respectively. Average values of triplicate determinations were used in calculations. Data obtained were subjected to statistical analysis to determine means and standard deviations of means. Significant differences between means of experiments were determined by analysis of variance (ANOVA). A significance level of 0.05 was chosen.
RESULTS AND DISCUSSION
Effect of agitation on dye biodecolourization in SSW: The culture under static conditions demonstrated a better decolorization than that under agitated conditions in 24 h. Only slight colour removal (7%) was obtained with the agitated culture, whereas more than 92.7% decolorization was obtained in culture kept under static conditions (Fig. 2).
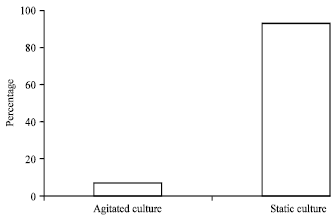 | |
| Fig. 2: | Effect of agitation on decolourization of acid orange 10 under saline conditions by Bacillus firmus |
Static or anoxic conditions were necessary for decolorization by the bacterium although higher cell growth was obtained under agitated conditions. Consequently, subsequently decolorization experiments were conducted under static conditions.
Aromatic compounds are susceptible to biological degradation under both aerobic and anaerobic conditions (Field et al., 1995; Olukanni et al., 2009). The enzymes, mono- and di-oxygenase, catalyse the ring fission of organic compounds by the incorporation of oxygen from O2 into the aromatic ring under aerobic conditions (Madigan et al., 2003). Nevertheless, azo dyes though aromatic compounds, are quite recalcitrant to aerobic bacterial degradation due to their substituents containing mainly nitro and sulfonic groups (Claus et al., 2002).
This is probably due to the electron-withdrawing nature of azo bonds and their resistance to oxygenases attack. Moreover, as decolorization occurs by the way of reduction of the azo dye which acts a final electron acceptor in the microbial electron transport chain, reducing equivalents might have a preference for more effective electron acceptors like oxygen in the medium which could then act as a limiting factor for dye removal (Cinar and Demiroz, 2010). It was speculated that under agitation conditions, the aerobic respiration of the strain might dominate the utilization of NADH and inhibit the azoreductase from obtaining electrons from NADH to decolorize azo dyes (Stolz, 2001). Similar results were obtained during the biodegradation of the azo dye C.I. Acid orange 7 by Pseudomonas sp. OX1 under a variety of operating conditions and it was found that the decolorization potential of the microorganism was fully exploited only in the absence of oxygen (Lodato et al., 2007). It was also reported in another study that the efficiency of aerobic treatment is inferior to that of anaerobic decolorization process (Forgacs et al., 2004).
Dye biodecolourization at different salt concentrations in SSW: The decolourization of Acid orange 10 by Bacillus firmus under high salt condition was examined while varying the saline conditions of the medium. In 24 h, colour removal in all culture flasks containing 0 to 6% (w/v) sodium chloride was more than 85% (Fig. 3). However, a gradual decline in decolorization was obtained with increase in sodium chloride concentration despite high decolorization extents obtained at salt concentrations below 6%. For example, complete colour removal at 1% sodium chloride concentration was obtained while decolorization extents obtained at 4 and 6% salt concentrations were 96.2 and 92.8%, respectively. Colour removal was inhibited at salt concentrations above 6%, albeit growth was observed at 8% salt concentration indicating the tolerance of the organism but the inhibition of decolourization at that concentration.
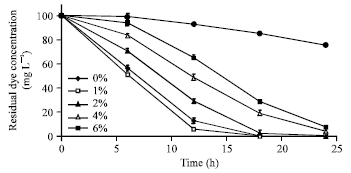 | |
| Fig. 3: | Time bound decolourization of Acid orange 10 by Bacillus firmus in media containing varying salt (NaCl) concentrations. Dye concentration 100 mg L-1; pH: 7; 30°C. Results are means of triplicate experiments (SD is indicated with error bars) |
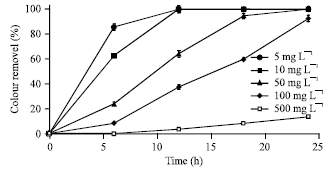 | |
| Fig. 4: | Time bound decolourization of different concentrations of Acid orange 10 by Bacillus firmus under high salt condition. NaCl concentration 6; pH: 7; 30°C. Results are means of triplicate experiments (SD is indicated with error bars) |
This inhibition effect may be attributed to the negative influence of salt on the permeability of cell membrane for dye ions and the relative competition between chloride and dye molecules for the active sites of Bacillus firmus at that concentration (8%) (Diniz and Volesky, 2005; Maurya et al., 2006). Usually, high salt concentrations (>1%) cause inhibition of dye decolourization due to the loss of microbial activity, cell dehydration and eventual plasmolysis of cells. As a result, low removal performance of dyes and the persistence of these environmental pollutants especially at high salt concentrations occur. However, results of this study demonstrates the halotolerance of the isolate and its capacity for growth and dye decolourization in a saline environment suggesting its potential for use in the treatment of wastewater with salt concentrations varying from 0-6%.
Dye biodecolourization at different dye concentration in SSW: Results obtained on experiments carried out to determine the effect of dye concentration on decolourization under saline conditions are presented in Fig. 4. The experiments were conducted consecutively from lower to higher (5-500 mg L-1) concentrations of the dye to encourage acclimatization and resistance of the bacterial cells to dye toxicity under saline conditions, besides achieving better decolourization results. Moreover, it has been stated that it is important to know whether microorganisms, for azo dye wastewater treatment applications, can bear high concentrations of dyes since dye concentration in a typical industrial effluent can vary between 10 and 50 mg L-1 (Padamavathy et al., 2003). The experimental data indicate that color removal was effective at dye concentrations below 100 mg L-1. Exposure of the bacterial cells to concentrations above 100 mg L-1 resulted in a decrease in color removal and cellular growth (data not shown). More than 92% colour removal at dye concentrations below 100 mg L-1 was obtained within 24 h (Fig. 4) whereas increase in dye concentration up to 500 mg L-1 showed a significant decrease in decolourization extent (13.8%). The decreased colour removal at higher dye concentrations may be attributed to the inhibition of bacterial growth and cellular metabolic activities due to increased toxic effect of the dye at such high concentrations. Moreover, it has also been observed that azo dyes with sulfonic acid (SO3H) groups on their aromatic rings greatly inhibited the growth of microorganisms at higher dye concentrations (Chen et al., 2003; Kalyani et al., 2009). Blockage of active sites of azoreductase by agglomerated dye molecules has also been adduced as a reason for decreased colour removal (Jadhav et al., 2008; Saratale et al., 2009; Tony et al., 2009). Nevertheless, the ability of Bacillus firmus to efficiently remove varying dye concentrations (5-100 mg L-1) within 24 h at 6% salt concentration suggests its potentials as a viable candidate for biological textile wastewater treatment applications.
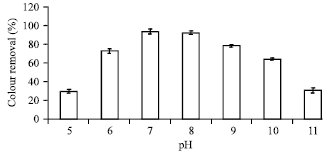
Effect of initial medium pH on dye biodecolourization in SSW: Percent decolorization of Acid orange 10 by B. firmus was checked in media containing varying initial pH. This was important since textile wastewater is slightly alkaline due to the high amounts of sodium hydroxide used in the scouring, washing and mercerizing steps of the textile operations (EPA, 1997; Mattioli et al., 2002; Dos Santos et al., 2006). It is believed that the effects of pH may be related to the transport of dye molecules across the cell membrane which is considered as the rate limiting step for the decolourization (Chang et al., 2001b; Kodam et al., 2005).
Cultures of pH 6-10 all reached decolorization extents of above 60% in 24 h, with pH 7 obtained as the optimal for colour removal (with a decolorization extent of 93.3%). Colour removal was effective within 24 h at all initial pH investigated except at pH 5 and 11 which showed 29.7 and 30.5% decolourization respectively (Fig. 5).
The potential of the bacterial isolate was such that it could decolourize the azo dye with the same efficiency across a broad pH spectrum. This is an indication of the adaptation of the microorganism to the varying pH of the wastewater from where it was isolated and its pH versatile nature. This observation is of significance for biological treatment of dye wastewater since it indicates the ability of the isolate to withstand varying pH conditions and thus, it’s potential for successful application in treatment of textile wastewater.
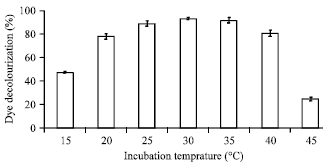
Effect of temperature on dye biodecolourization in saline wastewater: Temperature is an important factor for all processes associated with microbial vitality, including the remediation of wastewater. The effect of incubation temperature on colour removal from SSWM by Bacillus firmus is presented in Fig. 6.
 | |
| Fig. 5: | Influence of initial pH of medium on decolourization of Acid orange 10 by Bacillus firmus within 24 h under high salt condition. NaCl concentration 6%; dye concentration 100 mg L-1, pH: 7, 30°C. Results are means of triplicate experiments (SD is indicated with error bars) |
 | |
| Fig. 6: | Effect of temperature on decolourization of Acid orange 10 by Bacillus firmus within 24 h under high salt condition. NaCl concentration 6%, dye concentration 100 mg L-1, pH: 7. Results are means of triplicate experiments (SD is indicated with error bars) |
 | |
| Fig. 7: | Time bound decolourization of Acid orange 10 by Bacillus firmus in SSW containing different concentrations of SDS. NaCl concentration 6%; dye concentration 100 mg L-1, pH: 7, 30°C. Results are means of triplicate experiments (SD is indicated with error bars) |
Results suggest that the optimal range of temperature for dye decolourization was 20-40°C as no significant difference was obtained for % colour removal obtained at that range. Dye decolourization reached the maximum value of 93% at 30°C with a marginal reduction in the decolourization activity obtained below 20°C and above 40°C. The decolorization could still reach 80.6% at 40°C in 24 h, though the decolorization extent was lower than that at 30°C. The strain was inactive at 45°C, because no considerable cell growth was observed and just slight decolorization was obtained. The decline in colour removal at higher temperatures has previously been attributed to loss of cell viability or the denaturation of the azo reductase enzyme (Chang et al., 2001a; Saratale et al., 2009). In a previous study, Olukanni et al. (2009) showed that the optimum temperature for the degradation of Methyl red by a Micrococcus strain was 37°C.
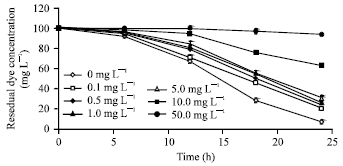
Dye biodecolourization at different SDS concentrations in SSW: The SDS concentrations (0.1, 0.5, 1.0 and 5.0 mg L-1) evaluated slightly restricted the colour removal capability of the microbe but did not adversely affect it as reduction in decolourization of only ca. 13-18% was obtained at those concentrations (Fig. 7). The optimum concentration of SDS for colour removal was 0.1 mg L-1. Surfactant concentrations above 5 mg L-1 proved to significantly suppress the decolourization of Acid orange 10 by the bacterium. Nevertheless, the considerable colour removal obtained in medium with SDS concentrations lower than 5 mg L-1 is encouraging because depending on the dyeing process, many chemicals which include Salts and Surfactants (SDS) may be added to improve dye adsorption onto the fibres (Dos Santos et al., 2007). Because of their amphipathic nature, surfactants are known to alter the structure and function of the cellular membrane, induce cellular lysis (Glover et al., 1999) and alter the structure and functions of important bacterial enzymes (Dong et al., 1997; Goncalves et al., 2003). Some studies have shown that surfactant addition reduces the biodegradation of pollutants because the toxicity of the surfactants decreases the viability and activity of bacterial cells (Sun et al., 2008). Therefore, a pre-requisite for effective biodegradation in surfactant-contaminated environments is that the degrading microbes are not adversely affected by the surfactant Plante et al. (2008). Since surfactants are present in textile wastewater, it is imperative to use isolates with enhanced resistance to surfactants for biotreatment in order to improve their potentials for industrial application. The resistance of the test isolate to the investigated surfactant concentrations in medium could possibly be explained by their adaptation to the surfactants occurring in textile wastewater from where it was isolated.
 | |
| Fig. 8: | Influence of additional carbon sources on mean residence time required for complete colour removal during degradation of Acid orange 10 by Bacillus firmus under high salt condition. NaCl concentration 6%; dye concentration 100 mg L-1, pH: 7, 30°C. Results are means of triplicate experiments (SD is indicated with error bars) |
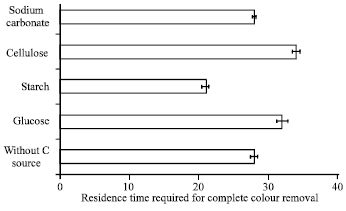
Effect of additional carbon source on dye biodecolourization in SSW: The effect of various carbon sources on the efficiency of colour removal by Bacillus firmus was determined using glucose, starch, cellulose and sodium carbonate as co-substrates in medium. The medium containing starch showed the fastest decolourization extent as complete colour removal in SSWM was obtained by 21 h whereas, complete colour removal in medium with no additional carbon source was obtained at 28 h (Fig. 8). Complete decolourization in medium with other added carbon sources was obtained at the following times: glucose (32 h); cellulose (34 h) and sodium carbonate (28 h). These alternative carbon sources did not enhance colour removal instead; their presence in medium caused a delay in complete colour removal in medium. The reason for the delay caused by inclusion of cellulose and sodium carbonate in medium is not known but in the case of glucose, its presence in medium may have resulted in its initial preference to the dye as a carbon source/substrate by the bacterium and/or may have reduced bacterial activity due to the slight drop in pH as a result of the production of organic acids during fermentation. In some studies, glucose had been added to enhance the decolourization performance of biological systems (Haug et al., 1991; Kapdan et al., 2000), while other authors have reported that glucose inhibited the decolourizing activity (Evangelista-Barreto et al., 2009; Knapp and Newby, 1995). The decrease in retention time for complete colour removal by starch is exciting since most textile wastewater contain starch arising from the sizing and desizing processes (Dos Santos et al., 2006). The sizing process utilizes sizing agents such as starch which are added to provide strength to the fibres and minimize breakage. Desizing is employed next to remove sizing materials prior to weaving.
CONCLUSION
In the present study, the ability of Bacillus firmus to decolourize Acid orange 10 under saline conditions and the effects of various process parameters on decolourization of the dye were investigated. Results obtained show the microorganism was halotolerant and was capable of effectively decolourization of the dye at different salt (1-6%) and dye (5-100 mg L-1) concentrations. The pH range for effective decolourization was 6-10 with pH 7 obtained as optimum for colour removal. Acid orange removal from solution was also effective at a temperature range of 20-40°C with 30°C obtained as optimum. Colour removal capacity of this bacterium was considerably stable at SDS concentrations between 0.1-5.0 mg L-1 and was enhanced in the presence starch; both compounds being components of industrial textile wastewater. The ability of this isolate to decolourize Acid orange 10 under saline conditions suggests its potential for use in the effective treatment of real dye wastewater.
REFERENCES
- Carliell, C.M., S.J. Barclay, N. Naidoo, C.A. Buckley, D.A. Mulholland and E. Senior, 1994. Anaerobic decolorisation of reactive dyes in conventional sewage treatment processes. Water SA, 20: 341-344.
Direct Link - Carliell, C.M., S.J. Barclay, C. Shaw, A.D. Wheatley and C.A. Buckley, 1998. The effect of salts used in textile dyeing on microbial decolourisation of a reactive azo dye. Environ. Technol., 19: 1133-1137.
Direct Link - Chang, J.S., Y.P. Chou and S.Y. Chen, 2001. Decolorization of azo dyes with immobilized Pseudomonas luteola. Process Biochem., 36: 757-763.
CrossRefDirect Link - Chang, J.S., C. Chou, Y.C. Lin, P.J. Lin, J.Y. Ho and T.L. Hu, 2001. Kinetic characteristics of bacterial azo-dye decolorization by Pseudomonas luteola. Water Res., 35: 2841-2850.
CrossRefDirect Link - Cinar, O. and K. Demiroz, 2010. Biodegradation of Azo Dyes in Anaerobic-Aerobic Sequencing Batch Reactors. In: Biodegradation of Azo Dyes (The Handbook of Environmental Chemistry), Erkurt, H.A. (Ed.). Vol. 9, Springer Verlag, Berlin Heidelber, ISBN-13: 978-3642118463, pp: 59-72.
CrossRef - Claus, H., G. Faber and H. Konig, 2002. Redox-mediated decolorization of synthetic dyes by fungal laccases. Appl. Microbiol. Biotechnol., 59: 672-678.
CrossRefDirect Link - De Baere, L.A., M. Devocht, P.V. Assche and W. Verstraete, 1984. Influence of high NaCl and NH4Cl salt levels on methanogenic associations. Water Res., 18: 543-648.
CrossRef - Delille, D., A. Basseres and A. Dessommess, 1998. Effectiveness of bioremediation for oil-polluted Antarctic seawater. Polar Biol., 19: 237-241.
CrossRefDirect Link - Dincer, A. and F. Kargi, 2000. Effects of operating parameters on performances of nitrification and denitrification processes. Bioprocess Biosyst. Eng., 23: 75-80.
CrossRefDirect Link - Diniz, V. and B. Volesky, 2005. Effect of counter ions on lanthanum biosorption by Sargassum polycystum. Water Res., 39: 2229-2236.
CrossRef - Dong, G., C. Vieille and J.G. Zeikus, 1997. Cloning, sequencing and expression of the gene encoding amylopullulanase from Pyrococcus furiosus and biochemical characterization of the recombinant enzyme. Applied Environ. Microbiol., 63: 3577-3584.
PubMedDirect Link - Dos Santos, A.B., I.A.E. Bisschops and F.J. Cervantes, 2006. Closing Process Water Cycles and Product Recovery in Textile Industry: Perspective for Biological Treatment. In: Advanced Biological Treatment Processes for Industrial Wastewaters, Cervantes, F.J., A.C. Van Haandel, S.G. Pavlostathis (Eds.). Vol. 1. International Water Association, London, pp: 298-320.
- Dos Santos, A.B., F.J. Cervantes and J.B. van Lier, 2007. Review paper on current technologies for decolourisation of textile wastewaters: Perspectives for anaerobic biotechnology. Bioresour. Technol., 98: 2369-2385.
CrossRefDirect Link - Dubey, S.K., A. Pandey, A.K. Bajaj and K. Misra, 2007. Some commercial azo dyes as inhibitors of mushroom tyrosinase DOPA oxidase activity. J. Pharmacol. Toxicol., 2: 718-724.
CrossRefDirect Link - El-Kady, M. and F. El-Shibini, 2001. Desalination in Egypt and the future application in supplementary irrigation. Desalination, 136: 63-72.
CrossRef - Evangelista-Barreto, N.S., C.D. Albuquerque, R.H.S.F. Vieira and G.M. Campos-Takaki, 2009. Cometabolic decolorization of the reactive azo dye orange II by Geobacillus stearothermophilus UCP 986. Text. Res. J., 79: 1266-1273.
CrossRefDirect Link - Field, J.A., A.J.M. Stams, M. Kato and G. Schraa, 1995. Enhanced biodegradation of aromatic pollutants in cocultures of anaerobic and aerobic bacterial consortia. Antonie van Leeuwenhoek, 67: 47-77.
PubMed - Forgacs, E., T. Cserhati and G. Oros, 2004. Removal of synthetic dyes from wastewaters: A review. Environ. Int., 30: 953-971.
CrossRefDirect Link - Glover, R. E., R.R. Smith, M.V. Jones, S.K. Jackson and C.C. Rowlands, 1999. An EPR investigation of surfactant action on bacterial membranes. FEMS Microbiol. Lett., 177: 57-62.
CrossRefPubMedDirect Link - Goncalves, A. M. D., M.R. Aires-Barros and J.M.S. Cabral, 2003. Interaction of an anionic surfactant with a recombinant cutinase from Fusarium solani pisi: A spectroscopic study. Enzyme Microb. Technol., 32: 868-879.
CrossRef - Guo, J., J. Zhou, D. Wang, C. Tian, P. Wang, M. Salah Uddin, 2008. A novel moderately halophilic bacterium for decolorizing azo dye under high salt condition. Biodegradation, 19: 15-19.
CrossRefPubMedDirect Link - Haug, W., A. Schmidt, B. Nortemann, D.C. Hempel, A. Stolz and H.J. Knackmuss, 1991. Mineralization of the sulfonated azo dye mordant yellow 3 by a 6-amononaphthelene-2-sulfonate-degrading bacterial consortium. Applied Environ. Microbiol., 57: 3144-3149.
PubMedDirect Link - Igwe, J.C., O.F. Mbonu and A.A. Abia, 2007. Sorption kinetics, intraparticle diffusion and equilibrium partitioning of azo dyes on great millet (Andropogon sorghum) waste biomass. J. Applied Sci., 7: 2840-2847.
CrossRefDirect Link - Jadhav, S.U., M.U. Jadhav, A.N. Kagalkar and S.P. Govindwar, 2008. Decolorization of brilliant blue G dye mediated by degradation of the microbial consortium of Galactomyces geotrichum and Bacillus sp. J. Chin. Inst. Chem. Eng., 39: 563-570.
CrossRefDirect Link - Kalyani, D.C., A.A. Telke, R.S. Dhanve and J.P. Jadhav, 2009. Ecofriendly biodegradation and detoxification of reactive red 2 textile dye by newly isolated Pseudomonas sp. SUK1. J. Hazard. Mater., 163: 735-742.
CrossRefPubMedDirect Link - Kapdan, I.K., F. Kargi, G. McMullan and R. Marchant, 2000. Decolorization of textile dyestuffs by a mixed bacterial consortium. Biotechnol. Lett., 22: 1179-1181.
CrossRefDirect Link - Kargi, F., 2002. Enhanced biological treatment of saline wastewater by using halophilic bacteria. Biotechnol. Lett., 24: 1569-1572.
CrossRefDirect Link - Kargi, F. and A. Dincer, 1996. Enhancement of biological treatment performance of saline wastewater by halophilic bacteria. Bioprocess Biosyst. Eng., 15: 51-58.
CrossRefDirect Link - Kinkle, B.K., M.J. Sadowsky, K. Johnstone and W.C. Koskinen, 1994. Tellurium and selenium resistance in Rhizobia and its potential use for direct isolation of Rhizobium meliloti from soil. Appl. Environ. Microbiol., 60: 1674-1677.
PubMed - Knapp, J.S. and P.S. Newby, 1995. The microbiological decolorization of an industrial effluent containing a diazo-linked chromophore. Water Res., 29: 1807-1809.
CrossRefDirect Link - Kodam, K.M., I. Soojhawon, P.D. Lokhande and K.R. Gawai, 2005. Microbial decolorization of reactive azo dyes under aerobic conditions. Word J. Microbiol. Biotechnol., 21: 367-370.
CrossRefDirect Link - Lodato, A., F. Alfieri, G. Olivieri, A. Di Donato, A. Marzocchella and P. Salatino, 2007. Azo-dye conversion by means of Pseudomonas sp. OX1. Enzyme Microb. Technol., 41: 646-652.
CrossRef - Manu, B. and S. Chauhari, 2003. Decolorization of indigo and azo dyes in semi continuous reactors with long hydraulic retention time. Process Biochem., 38: 1213-1221.
CrossRef - Margesin, R. and F. Schinner, 2001. Potential of halotolerant and halophilic microorganisms for biotechnology. Extremophiles, 5: 73-83.
CrossRef - Mattioli, D., F. Malpei, G. Bortone and A. Rozzi, 2002. Water Minimization and Reuse in Textile Industry. In: Water Recycling and Resource Recovery in Industry: Analysis, Technologies and Implementation, Lens, P.N.L., L. Hulshoff Pol, P. Wilderer and T. Asano (Eds.). IWA Publishing, Cornwall, UK., ISBN-13: 9781843390053, Pages: 677.
- Maurya, N.S., A.K. Mittal, P. Cornel and E. Rother, 2006. Biosorption of dyes using dead macro fungi: Effect of dye structure, ionic strength and pH. Bioresour. Technol., 97: 512-521.
CrossRef - Nriagu, J.O. and J.M. Pacyna, 1988. Quantitative assessment of worldwide contamination of air, water and soils by trace metals. Nature, 333: 134-139.
CrossRefDirect Link - Ogugbue, C.J. and T. Sawidis, 2011. Optimisation of process parameters for bioreduction of azo dyes using Bacillus firmus under batch anaerobic condition. Int. J. Environ. Stud., 68: 651-665.
CrossRef - Olukanni, O.D., A.A. Osuntoki and G.O. Gbenle, 2009. Decolourization of azo dyes by a strain of Micrococcus isolated from a refuse dump soil. Biotechnology, 8: 442-448.
CrossRefDirect Link - Omar, H.H., 2008. Algal decolorization and degradation of monoazo and diazo dyes. Pak. J. Biol. Sci., 11: 1310-1316.
CrossRefPubMedDirect Link - Ozalp, G.G., Y.C. Gomec, S. Gonuldinc, I. Ozturk and M. Altinbas, 2003. Effect of high salinity on anaerobic treatment of low strength effluents. Water Sci. Technol., 48: 207-212.
PubMedDirect Link - Padamavathy, S., S. Sandhya, K. Swaminathan, Y.V. Subrahmanyam and S.N. Kaul, 2003. Comparison of decolorization of reactive microorganisms isolated from various sources. J. Environ. Sci., 15: 628-632.
PubMedDirect Link - Rai, H.S., M.S. Bhattacharyya, J. Singh, T.K. Bansal, P. Vats and U.C. Banerjee, 2005. Removal of dyes from the effluent of textile and dyestuff manufacturing industry: A review of emerging techniques with reference to biological treatment. Crit. Rev. Environ. Sci. Technol., 35: 219-238.
CrossRefDirect Link - Rajendran, R., S.K. Sundaram and K.U. Maheswari, 2011. Aerobic biodecolorization of mixture of azo dye containing textile effeluent using adapted microbial strains. J. Environ. Sci. Technol., 4: 568-578.
CrossRef - Rajeswari, K., R. Subashkumar and K. Vijayaraman, 2011. Biodegradation of mixed textile dyes by bacterial strains isolated from dyewaste effluent. Res. J. Environ. Toxicol., 5: 97-107.
CrossRefDirect Link - Saratale, R.G., G.D. Saratale, J.S. Chang and S.P. Govindwar, 2009. Ecofriendly decolorization and degradation of Reactive Green 19A using Micrococcus glutamicus NCIM-2168. Bioresour. Technol., 110: 3897-3905.
PubMed - Stolz, A., 2001. Basic and applied aspects in the microbial degradation of azo dyes. Applied Microbiol. Biotechnol., 56: 69-80.
CrossRefDirect Link - Sun, N., H. Wang, Y. Chen, S. Lu and Y. Xiong, 2008. Effect of surfactant SDS, Tween 80, Triton X-100 and rhamnolipid on biodegradation of hydrophobic organic pollutants. Proceedings of the 2nd International Conference on Bioinformatics and Biomedical Engineering, May 16-18, 2008, Shanghai, China, pp: 4730-4734.
CrossRef - Tchatchueng, J.B., B.B. Loura, J. Atchana and R. Kamga, 2009. TiO2-MoO3 as photocatalyst for azo and triphenylmethane dyes decolouration. J. Environ. Sci. Technol., 2: 31-39.
Direct Link - Tony, B.D., D. Goyal and S. Khanna, 2009. Decolorization of direct red 28 by mixed bacterial culture in an up-flow immobilized bioreactor. J. Ind. Microbiol. Biotechnol., 36: 955-960.
CrossRefPubMedDirect Link - Yasar, A., N. Ahmad, A.A.A. Khan, H. Khan and M. Khalid, 2007. Kinetics of UV, fenton and photo fenton processes for decolorization of various red CL-5B Dye concentrations. J. Applied Sci., 7: 2339-2344.
CrossRefDirect Link - Yoo, E.S., J. Libra and U. Wiesmann, 2000. Reduction of azo dyes by Desulfovibrio desulfuricans. Water Sci. Technol., 41: 15-22.
Direct Link - Zhang, F., A. Yediler, X. Liang and A. Kettrup, 2004. Effects of dye additives on the ozonation process and oxidation by products: A comparative study using hydrolysed CI Reactive red 120. Dyes Pigments, 60: 1-7.
CrossRef - Chen, K.C., J.Y. Wu, D.J. Liou and S.C.J. Hwang, 2003. Decolorization of the textile dyes by newly isolated bacterial strains. J. Biotechnol., 101: 57-68.
CrossRefDirect Link