
F.J. Al- Saffar
Department of Anatomy, College of Veterinary Medicine, Baghdad University, Baghdad, Iraq
LiveDNA: 964.4860
A.G. Al- Haaik
Department of Anatomy, College of Veterinary Medicine, University of Mosul, Mosul, Iraq
LiveDNA: 964.14952
Singapore Journal of Chemical Biology
Year: 2016 | Volume: 5 | Issue: 1 | Page No.: 11-19







ABSTRACT
Objective: The project aimed to study the relationship between paneth and stem cells in the developed small intestine of rabbits. Methodology: To conduct this investigation, 48 offspring rabbits at different postnatal ages (1, 10, 15 and 40 days) were collected and set equally into four groups. Results: Microscopic findings revealed absence of paneth cells in the crypts of small intestine of one day aged kits. In 10 days aged suckling puppies and subsequent ages, paneth cells were detected with variable size, structure and density of their cellular granules in relation to different ages and their maturity. Micromorphometric measurements showed lowest mean diameter of paneth cell at the age of 10 days in jejunum and the highest was at the age of 15 days in duodenum. The number of paneth cells was increased with age progress in three segments of small intestine but the lowest mean number of them was at the age of 10 days in duodenum and ileum but the highest was at the age of 40 days in jejunum. The Lgr5 positive intestinal stem cells found at the bases of the villi and at inter-villus spaces even in kits. In 10 days aged suckling puppies, few numbers of mitotic figures was detected in some newly formed crypts. At subsequent ages, a dramatic increase in the number of mitotic figures was recorded in the crypts of Lieberkühn. Conclusion: It can be concluded that paneth cells may enhance the process of intestinal stem cell mitosis and subsequently the growth of the intestine.
PDF Abstract XML References Citation
Received: July 23, 2016;
Accepted: August 04, 2016;
Published: September 15, 2016
How to cite this article
F.J. Al- Saffar and A.G. Al- Haaik, 2016. Histomorphological Relationship of Paneth Cells with Stem Cells in the Small Intestine of Indigenous Rabbit at Different Postnatal Ages. Singapore Journal of Chemical Biology, 5: 11-19.
URL: https://scialert.net/abstract/?doi=sjchbio.2016.11.19
URL: https://scialert.net/abstract/?doi=sjchbio.2016.11.19
INTRODUCTION
It is well known that the mammalian intestine is covered by bulged villus epithelium functioning in digestion and absorption and precipitate in host defense and removal of pathogens1. This intestinal epithelium contains absorptive enterocytes and secretory cells that are renewed from the stem cells exist at the lower parts of the intestinal crypts. Infact, goblet and paneth cells are continue to share similar characters during their development, whereas enteroendocrine cells are developed separately through different mechanism. During their process of differentiation, paneth cells move back to the bases of the crypts and become interspersed between the stem cells whereas, the other types of cells are migrated toward the tips of villi2,3.
Paneth cells were described in the past century as granulated cells situated at the bases of the small intestinal glands which are called crypts of Lieberkühn. Few hypotheses were considered them important regulator of innate intestinal defense. They regulates microbial density persist in the small intestinal lumen and so play a role in the protection of nearby stem cells. They migrated downward to the stem cells zone directed toward the crypts bases and they are considered relatively long lived cells (20 days compared with 3-5 days for enterocytes). Mature paneth cells are capable to secrete both antimicrobial peptides (cryptdin) and the growth factors4. The latter, postulated that migration, high density and long residency time of these cells at the crypt base may influence the structure and/or function of the niche of the adjacent stem cells.
In the past decade, Zanuzzi et al.5 documented that the intestinal epithelium has a critical role in host defense out of which paneth cell is one of the specialized existed cellular epithelial type involved in such function. It secretes many substances of antimicrobial properties in response to different stimuli. Under pathological conditions, changes were reported in their number, morphology and location as well as in their cytoplasmic granules number. Histologically, paneth cell are pyramidal in shape containing a basal spherical nucleus with a visible nucleolus and slightly eosinophilic stained cytoplasm. They are usually set in tight groups exhibiting ill-defined cellular borders. Their heterogeneous populations of secretory granules with heterogeneous size were recognized in some cells when stained with hematoxylin and eosin, Masson’s trichrome and PAS staining techniques.
Multilabel Immunocytochemical studies conducted by Bry et al.6 on the developing mouse intestine revealed the differentiation of paneth cells by the 7th postnatal day. They found out morphologic appearance of paneth cells with apical secretory granules in most of intestinal cryptdin-positive cells.
Recent considerations postulated that the gastrointestinal tract is an important site of interactions between the body and the exterior surroundings or environment. Accordingly, the intact mucosal epithelial lining constructs the first line of defense against different exterior attacks which may be chemical, mechanical or microbial in nature and subsequently renews itself rapidly as a mechanism to maintain the homeostasis7,8. Infact, the action of cellular regeneration and replacement is conducted via tissue-restricted adult stem cells which are existed at the bases of the intestinal crypts. These cells go through essential symmetric divisions. They were randomly generating a larger group of more rapidly dividing progenitor cells referred to as transit-amplifying cells. The latter cells give rise to the four terminally differentiated types of cells in intestinal mucosa that are absorptive enterocytes, goblet cells, secretory enteroendocrine cells and paneth cells9. Out of them, the differentiated enterocytes constructs the major type of cells of the intestinal mucosa, which undergoes a process of apoptosis and is usually extruded into the lumen of the bowel.
Zhou et al.10 proposed that the intestinal mucosa undergoes usually continuous processes such as proliferation, differentiation and apoptosis which are well coordinated by many signaling pathways. The epithelial lining of the intestine is usually undergoes such constantly rapid renewal processes. The intestinal glands (crypts of Lieberkühn) are highly dynamic niche with multipotent stem cells positioned in their lower third parts. These cells act to generate new cells that are eventually differentiated into the four above mentioned specialized cell types of the small intestine.
Cancer was proposed to be the disease of the stem cells because they are the only long-lived cells in a promptly proliferating tissue one of them is the intestinal tract; a trait which makes them excellent targets for the accumulation of successive mutations required for the evolution of the tumors. Actually, stem cells were expected to be responsible for the initiation, maintenance and recurrence of the tumors11.
Researchers focused on the properties of stem cells provide a foundation for therapeutic advancement in oncology, clinical genetics and a diverse array of degenerative disorders. For instance, the elucidation of cellular pathways governing proliferative regulation and differentiation will propose medical strategies regarding the cause of cancer. Intestinal stem cells reside at the basal part of the crypt and give rise to all cell types found within the crypt. These are specialized epithelial cells characterized by their ability for self-maintenance and provide the basis for ongoing cell replacement. They are capable to divide asymmetrically, producing one stem cell, which remains undifferentiated multipotent and one daughter cell, which is committed to differentiation. Committed daughter cells undergo several additional rounds of cell division during their migration from the proliferative to the differentiated compartment. Upon differentiation, cells lose their ability to divide and eventually died. The mature gastrointestinal system therefore comprises, at all times, both undifferentiated, differentiated pluripotent stem cells and functional epithelial cells12.
In the colon, stem cells are located at the base of the crypts with proliferation occurring in the lower third of their epithelial folds. Epithelial cells are migrated in ordered cohorts towards the luminal surface then differentiated to form colonocytes, goblet cells and enteroendocrine cells. Cells are entered a death program at the luminal surface characterized by senescence or apoptosis and are sloughed into the lumen13. More than 80% of all epithelial tissue in the colon comprises colonocytes14. Colonocytes have both absorptive and secretory functions. It absorbed water, sodium and short-chain fatty acids while secreting bicarbonate and potassium. Goblet cells meanwhile, elaborate mucins, trefoil proteins and other factors that help to protect the intestinal mucosa against injuries and facilitate tissue repair15.
Koo and Clevers16 referred about the innovation of LGR5 marker of the intestinal stem cells which led to the development explosively of much new line of researches. In fact the inner mechanisms of the stem cell niche in the intestinal crypt are now well understood. Now-a-days, LGR5(+) stem cells can be cultured over long periods in vitro as epithelial organoid and such technology opens new possibilities of using these cultured adult stem cells for gene therapy, drug advances, disease modeling and regenerative medicine.
MATERIALS AND METHODS
Rabbit’s collection and study design: Twenty pregnant indigenous does were kept and maintained under laboratory conditions of temperature 25°C and 12 h day, 12 h night cycle and allowed free access of food (standard basal diet and greenish food) and tap water ad libitum. Animals were purchased from Animal House of the College of Veterinary Medicine, Baghdad University. From these does, 48 offspring rabbits at different postnatal ages (regardless to sex) were collected and set equally into four groups, each of 12 animals. The newly born kits of 1 day age were set as (P1) group. The second group of 10 days aged suckling puppies which was set as (P10). The third group of 15 days of age of suckling and feeding on food was set as pre-weaned group (P15). Whereas, the fourth group of 40 days of age was set as post-weaned animals fed on solid basal diet and greenish (P40) and weaning was set on day 30 of age.
Animals were euthanized by intra-cardiac injection of over dose of sodium pentobarbital17 (100 mg kg–1). The representative specimens of 1 cm were cut from each segment of small intestine (mid-duodenum, mid-jejunum and mid-ileum). All specimens were evacuate from the intestinal content by washing with cold normal saline18 and then directly immersed in 10% neutral buffered formalin for 48 h. Specimens were processed by routine histological processing method. About 5 μm paraffin sections were obtained by using rotary microtome19. For the aim of staining of the histological sections and applying some measurements to paneth cell, the following stains and procedures were conducted:
| • | For the demonstration of paneth cells in the bases of the crypts of the small intestine and study its morphology and distribution H and E, Masson’s trichrome and Mallory trichrome stains were used19 |
| • | For counting of paneth cell, 8-24 crypts were evaluated only for those that were aligned along the longitudinal axis and that the lumen of the crypt can be visualized along its length, counting was performed at a constant magnification (X1000) counting the number of paneth cells per crypt20 |
| • | Diameter of paneth cell (long axis+short axis/2) were measured using the color USB 2.0 digital image system (Scope Image 9.0) which was provided with image processing software |
All the above parameters and obtained data were set in tables to compare through different postnatal ages as well as among different segments of small intestine within each age.
Immunohistochemical procedure for the stem cells: Anti-Lgr5 antibody (US biological/USA) and ImmunoCruz™ rabbit LSAB staining system: sc-2051 (santacruz/USA) will be used as intestinal stem cell marker for identification of the intestinal stem cells and their dividing in different parts of the small intestine (duodenum, jejunum and ileum) during the different selected developing periods (1, 10, 15 and 40 days of age). Sections of 5 μm thick were used to conduct this technique which was processed according to the procedure mentioned by provider of the staining kit.
RESULTS
Microscopic examination of the intestinal wall revealed that its structure was composed of four main tunicae that were mucosa, submucosa, muscularis and serosa. Tunica mucosa showed villi which were finger shaped projections and crypts of Lieberkühn which were invaginations located within lamina propria. Villi and crypts were lined with simple columnar cells intermingled with numerous goblet cells.
At one day of age, microscopic examination of the crypts of three segments of small intestine of kits (P1) revealed absence of paneth cells in their lining epithelium. In suckling puppies (P10) the lining cells of the crypts revealed the presence of paneth cells which were relatively large and spherical cells with basely located triangular nuclei. They were filled with acidophilic granules and easily identified. Most of paneth cells were located at the bases of the crypts of Lieberkühn and few cells were detected at the level of their lower third part. The cytoplasmic granules of paneth cells appeared rounded and orange to red in color when stained with either hematoxylin and eosin stain or Masson’s trichrome stains and blue with Mallory trichrome stain (Fig. 1). The size, structure and density of their cellular granules were variable in relation with the different ages and the maturity of the cells themselves. Current findings showed that the lowest mean diameter of paneth cell was at the age of 10 days in jejunum and the highest was in ileum (Fig. 1, Table 1).
The granules of paneth cells were denser and ill-defined in the age of 10 days and some cells contain semi-homogenous acidophilic material instead of granules distributed in the cytoplasm, whereas appeared well defined rounded granules at both ages of 15 and 40 (Fig. 1). Actually, current findings revealed that some crypts were possessed one or more paneth cells and others were fully free of paneth cell so that the recorded mean number was about 0.5 paneth cell per each crypt (Table 2). In the age of 15 days, evident increase in the size of paneth cells was noticed and at which the highest diameter was recorded in the crypts of the duodenal crypts.
| Table 1: | Diameter of paneth cells (μm) |
 | |
| The similar letters in columns indicates non-significant differences (p<0.05) among age’s and different letters in columns indicates significant difference (p<0.05) among age’s | |
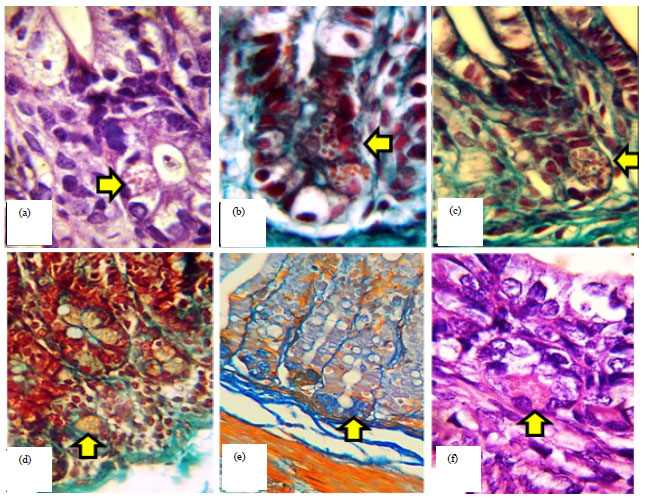 | |
| Fig. 1(a-f): | Photomicrograph of paneth cells (yellow arrows) at different postnatal ages, (a) Upper panel: Duodenum of P1, P10 and P15 stained with H and E, (b, c) Masson trichrome at X100, (d) Lower panel: Jejunum of P40 stained by Masson trichrome, (e) Mallory trichrome at X40 and (f) Ileum of P40 stained by H and E at X100 |
Most of the paneth cells were located at the bases of crypts, the density of their granules were decreased but seems to be well-defined spherical acidophilic granules. The number of paneth cells was increased upto 1.7, 2.1 and 0.9 paneth cells/crypt in duodenum, jejunum and ileum, respectively. However, few crypts were found free of paneth cells at the bases of the crypts.
In the post-weaned rabbits of 40 days of age, current findings showed that paneth cells were reduced in their sizes of which most of them were located at the bases of the crypts. Their granules were well-defined and densely packed. The number of paneth cells was increased in this age (2.1, 2.9 and 1.3 crypt in duodenum, jejunum and ileum, respectively). Most of the intestinal crypts were possessed paneth cells and the data of paneth cell number and their diameters were listed in Table 1 and 2.
To study the dividing intestinal stem cell at different postnatal ages of the local studied rabbits, Lgr5 stem cell marker was conducted for such purpose. Light microscopy of the small intestinal mucosa of the three intestinal segments revealed that newly born kits at one day of age, possessed intestinal stem cells located at the bases of the villi and at inter-villus spaces. They appeared dark brown cells with darkly stained nuclei, accompanied with large number of immature cells of large pale nuclei. Few mitotic figures were observed at this age in the intestinal mucosa and most of them were in the interphase stage. At 10 days of age, intestinal mucosa of suckling puppies showed relative raise in number of mitotic figures and they were detected in some newly formed crypts (1-2 mitotic figure/10 fields under microscopic field of X40). Most of these figures were observed in metaphase and anaphase stages. Such microscopic findings i.e., mitotic figures were detected in few crypts at this age similarly to those of one day kits (Fig. 2).
At subsequent age, pre-weaned rabbits (15 days of age) as well as the next age of 40 days, a dramatic increase in the number of mitotic figures was recorded in the crypts of Lieberkühn.
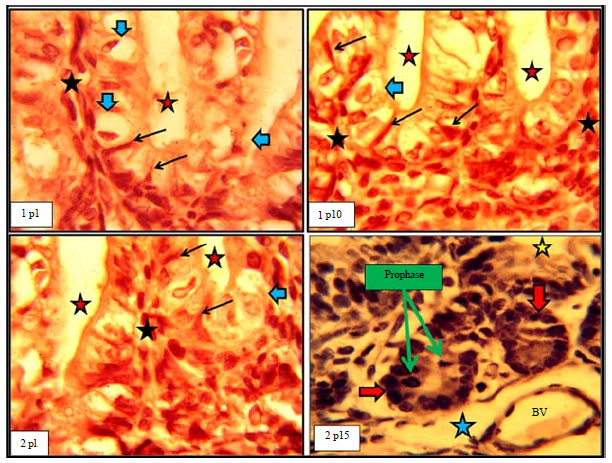 | |
| Fig. 2: | Photomicrograph of stem cells (black arrows) at intestinal crypts (red arrows), Upper panel: Stem cells present in the duodenum of one day aged kits (1 P1), duodenum of 10 days suckling puppies (1 P10), Lower panel: Jejunum of kits (2 P1), Jejunum of 15 days, pre-weaned rabbits showed two cells in prophase stage of mitosis (green arrows), intervillus lumina (red stars), immature cells (blue arrows), core of villi (black stars), blood vessel (BV) in lamina propria, X100 and Lgr5 marker |
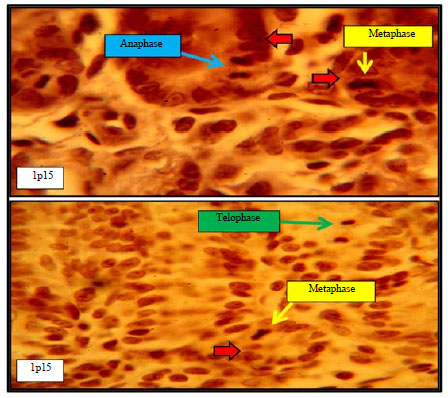 | |
| Fig. 3: | Photomicrograph of duodenal crypts (red arrows) of 15 days aged pre-weaned rabbits, different stages of mitosis (anaphase, metaphase and telophase) X100 and Lgr5 marker |
| Table 2: | Number of paneth cells/crypt |
 | |
| The different letters in each columns indicates significant difference (p<0.05) among age’s | |
There was about 1-3 mitotic figure/crypt (under microscopic field of X40). Obviously, all stages of mitosis were detected in the intestinal mucosa of these rabbits that were prophase, metaphase, anaphase and telophase. In this age most of the intestinal crypts viewed mitotic figures at different stages of mitosis (Fig. 3).
DISCUSSION
The current findings showed decline in the number of paneth cells in the direction extended from duodenum to the ileum. Such decline was in agreement with those observed by Zanuzzi et al.5 through his study on rabbits. The latter Immunocytochemical investigation referred to that the number of crypts containing cryptdin-positive paneth cells and the number of positive cells per crypt was decreased progressively along the duodenal-ileal axis. In addition, they found dramatic increase in paneth cell number per crypt during the period extended between 14th and 28th which was around weaning time in the current study.
Currently, the lining cells of the crypts revealed the presence of paneth cells which identified clearly in 10 days aged suckling puppies and subsequent ages by using hematoxylin and eosin, Masson’s trichrome and Mallory stains, it was absent in kits aged one day. Most of paneth cells were located at the bases of the crypts of Lieberkühn and few cells were detected at the level of lower third of the crypts. Differently, in the intestine of the human, paneth cells were not restricted to the bases of the crypts but also to the entire length of the crypts and even in the villi21. Similarly in sheep, Eurgun et al.22 reported absence of paneth cells in the villi of their small intestinal epithelium and that the paneth cells were differentiated as such towards the bases of the crypts. Actually, the demonstration of paneth cell and its acidophilic granules require adequate differential stains, however similar to the current study, Zanuzzi1 et al.5 and Vasquez Cachay et al.23 and others were applied hematoxylin and eosin and Masson’s trichrome stains for the demonstration of paneth cells.
The highest number of paneth cells was recorded in the jejunum of 40 days post-weaned rabbits but the largest size of paneth cell was recorded in the age of 15 days. Similar to these findings, Bry et al.6 found that paneth cell can detected only at 7th postnatal day in developing mice because of shorter periods of suckling and weaning in these animal species. Whereas, Pfoze and Singh24 referred to the possibility of paneth cell detection in human only at 24 weeks and at this age they were comparable to the histological appearance of adult Paneth cells. On the other hand, Vasquez Cachay et al.23 found morphologically the paneth cells in the guinea pig at day 2 using Masson’s trichrome staining technique and at day 4 by applying hematoxylin and eosin. They recorded differences between the studied ages and measured highest number of paneth cells in the duodenum of 13 days old piglets. Current findings confirmed the presence of paneth cells in the small intestine of indigenous rabbits from 1st week of postnatal life but not at birth.
The paneth cell reaches its well-developed stage in the age of 15 days (critical age) where at this period the growing rabbits start to feed, in addition to its mother milk, on greenish fibers which may be contaminated hence, the rabbit needs special defense mechanism which can be provided by paneth cells. Additionally, the highest number of paneth cells in this age was recorded in the jejunum and this result was not parallel with those recorded by Zanuzzi et al.5 who mention that by the 14th postnatal day, the number of cryptdin-positive crypts in both proximal and distal halves of the small intestine was equal.
Currently, the immunohistochemical procedure showed that the intestinal stem cells (ISCs) were characterized by high expression of Lgr5 marker. They were intervening between paneth cells at the bases of intestinal crypts. The expression of lgr5 stem cell as well as the mitotic figures of dividing stem cell was well studied in the intestine of local rabbits at different selected postnatal ages according to type of feeding. In one day aged kits and 10 days aged suckling puppies, the use of lgr5 stem cell marker revealed presence of intestinal stem cells (ISCs) at the bases of the villi and in the newly formed crypts intermingled with immature enterocytes. Actually, no mitotic figures were observed in those of one day but few figures were showed interphase and metaphase in the suckling puppies of 10 days. The current identification of ISCs by the application of Lgr5 marker was well assessed and confirmed its previous application in recent study of Barker et al.25 and Van der Flier et al.26 in human and Snippert et al.9 in mice. Infact, these cells were named previously as crypt base columnar cells27,28.
Current findings indicated proportional relationship between paneth and stem cells existence at the bases of the intestinal crypts. Actually, the outcome showed higher number of paneth cells found in the age of 15 days simultaneously the mitotic figures of the dividing stem cells in the crypts were increased dramatically and such feature may explain the effect of paneth cells on the adjacent stem cells by enhancing their ability for multiplication by mitosis. The relationship between these two types of cells was mentioned unclearly before6,22,29. However, current observations reflected obvious relationship between them predominantly during pre-weaned period of the local pre-weaned rabbits. This positive relationship came in inconsistency with recent hypotheses of Snippert et al.9. The latter studied intestinal crypt homeostasis and they found that the growth of cultured stem cells if paired with paneth cell was higher than those cultured alone without paneth cell. In other words, more than half of Lgr5 stem cells grew out into an intestinal organoid when sorted together with a adjacent paneth cells. Other researchers such as Potten et al.13, Sato et al.30 and Barker et al.31 were referred to the relationship between paneth and stem cells and they considered paneth cell as niche or supporting cell for stem cell because they were intermingled together at the crypts bases.
Two types of stem cells population are believed to be present in the crypts of Lieberkühn, the first is the crypt base columnar cells located in the deepest part of the crypts which are usually divided a symmetrically to give rise two types of cells and the second is called the transit amplifying cells which have stemness properties and are usually divided symmetrically to give rise one type of cells29,31. Current high mitotic figures that were detected in 15 days aged pre-weaned rabbits may represent the second type of such population. Whereas, the first type was showed lesser dividing activity because it divides to generate new stem cell and other cell of the second type of population in each mitotic process. Currently, examination of Lgr5 stem cells in the small intestine showed orientation of mitotic spindle in apical-basal axis of the most divided cells at the bases of intestinal crypts predominately at 15 days aged pre-weaned rabbits. The arrangement of the mitotic spindle in Lgr5 stem cells was also described recently by Quyn et al.32 in the small intestine and colon of both human and mouse. The existence and prominent mitotic character of these stem cells may be occurred due to their capacity to divide approximately once per day25. Current results showed that there was a relationship between stem cell proliferation and apoptosis due to the apparent manifestation of mitotic figures of stem cell and apoptotic cells in the same age of 15 days. Similarly to the findings of Ramachandran et al.33, the cell proliferation was confined to the crypts while differentiation occurs during rapid migration of cell toward the villi the differentiated cells underwent a process of programmed cell death called apoptosis. The combination of apoptosis and cell proliferation was known responsible for shaping tissues and organs during the processes of development34. Infact, apoptosis found plays an important role in controlling lineage tracing from different intestinal stem cells populations in the mouse intestine35.
CONCLUSION
This study aimed to find out the relationship between paneth cells and intestinal stem cells during the development of the digestive tract of the rabbit which is one of the important animal models in study. Precisely to find the effect control of paneth on the mitosis of stem cell which are propagated continuously to regenerate the apoptotic cells or dead cells of the other types of cells exist and structured the intestinal epithelium. Current findings showed such relationship indicated by the presence of numerous mitotic divisions in crypts comprised fair number of paneth cells and vise versa. Actually such effect was obvious at 15 days of age which was the active developmental stage in the studied rabbits. Paneth cell can be concluded very important structural intestinal cells in the normal development of the intestine. This data expected to provide good base to other study such as cancer investigations of intestine in human and even in animals because such disease was related to the abnormal changes in the genetic materials of stem cells in subsequently influence the normal growth of the intestinal tract. To rabbits, many countries considered this animal a good source of food for human, so that intestinal trouble may demolish such industry. Care should be focused on the developed rabbits particularly during period around weaning to obtain subsequently fully healthy animals so that obtaining good source of food for human.
In conclusion, there was a proportional relationship between maturation of paneth cells and intestinal stem cells through increase number of divided stem cell simultaneously with development of paneth cells.
ACKNOWLEDGMENT
The authors strongly acknowledge the council of the Veterinary Medicine College, Baghdad University to support this study.
REFERENCES
- Miller, H., J. Zhang, R.K. Lee, G. B. Patel and W. Chen, 2007. Intestinal M cells: The fallible sentinels? World J. Gastroenterol., 13: 1477-1486.
PubMedDirect Link - Schuijers, J. and H. Clevers, 2012. Adult mammalian stem cells: The role of Wnt, Lgr5 and R-spondins. EMBO J., 31: 2685-2696.
CrossRefDirect Link - Garabedian, E.M., L.J.J. Roberts, M.S. McNevin and J.I. Gordon, 1997. Examining the role of paneth cells in the small intestine by lineage ablation in transgenic mice. J. Biol. Chem., 272: 23729-23740.
CrossRefPubMedDirect Link - Zanuzzi, C.N., P.A. Fontana, C.G. Barbeito, E.L. Portiansky and E.J. Gimeno, 2008. Paneth cells: Histochemical and morphometric study in control and Solanum glaucophyllum intoxicated rabbits. Eur. J. Histochem., 52: 93-100.
PubMedDirect Link - Bry, L., P. Falk, K. Huttner, A. Ouellette, T. Midtvedt and J.I. Gordon, 1994. Paneth cell differentiation in the developing intestine of normal and transgenic mice. Proc. Natl. Acad. Sci. USA., 91: 10335-10339.
Direct Link - Quante, M. and T.C. Wang, 2009. Stem cells in gastroenterology and hepatology. Nat. Rev. Gastroenterol. Hepatol., 6: 724-737.
CrossRefDirect Link - Barker, N., J.H. van Es, J. Kuipers, P. Kujala and M. van den Born et al., 2007. Identification of stem cells in small intestine and colon by marker gene Lgr5. Nature, 449: 1003-1007.
CrossRefDirect Link - Snippert, H.J., L.G. van der Flier, T. Sato, J.H. van Es and M. van den Born et al., 2010. Intestinal crypt homeostasis results from neutral competition between symmetrically dividing Lgr5 stem cells. Cell, 143: 134-144.
CrossRefDirect Link - Zhou, Y., P. Rychahou, Q. Wang, H.L. Weiss and B.M. Evers, 2015. TSC2/mTORC1 signaling controls Paneth and goblet cell differentiation in the intestinal epithelium. Cell Death Dis., Vol. 6.
CrossRefDirect Link - Zeki, S.S., T.A. Graham and N.A. Wright, 2011. Stem cells and their implications for colorectal cancer. Nat. Rev. Gastroenterol. Hepatol., 8: 90-100.
CrossRefDirect Link - Bach, S.P., A.G. Renehan and C.S. Potten, 2000. Stem cells: The intestinal stem cell as a paradigm. Carcinogenesis, 21: 469-476.
CrossRefDirect Link - Potten, C.S., C. Booth and D. Pritchard, 1997. The intestinal epithelial stem cell: The mucosal governor. Int. J. Exp. Pathol., 78: 219-243.
CrossRefDirect Link - Chang, W.W.L. and C.P. Leblond, 1971. Renewal of the epithelium in the descending colon of the mouse. I. Presence of three cell populations: Vacuolated-columnar, mucous and argentaffin. Am. J. Anatomy, 131: 73-99.
CrossRefDirect Link - Deplancke, B. and H.R. Gaskins, 2001. Microbial modulation of innate defense: Goblet cells and the intestinal mucus layer. Am. J. Clin. Nutr., 73: 1131S-1141S.
PubMedDirect Link - Koo, B.K. and H. Clevers, 2014. Stem cells marked by the R-spondin receptor LGR5. Gastroenterology, 147: 289-302.
CrossRefDirect Link - Suvarna, S.K., C. Layton and J.D. Bancroft, 2013. Bancroft's Theory and Practice of Histological Techniques. 7th Edn., Churchill Livingstone, Edinburgh, Scotland, ISBN-13: 9780702050329, Pages: 637.
Direct Link - Di Sabatino, A., E. Miceli, W. Dhaliwal, P. Biancheri and R. Salerno et al., 2008. Distribution, proliferation and function of Paneth cells in uncomplicated and complicated adult celiac disease. Am. J. Clin. Pathol., 130: 34-42.
CrossRefDirect Link - Deschner, E.E., 1967. Observations on the Paneth cell in human ileum. Exp. Cell Res., 47: 624-628.
CrossRefDirect Link - Eurgun, E., L. Eurgun, R.N. Asti and A. Kurum, 2003. Light and electron microscopic morphology of Paneth cells in the sheep small intestine. Revue Medecine Veterinaire, 154: 351-355.
Direct Link - Vasquez Cachay, M.E., E.P. Gomez, J.L. Rodriguez Gutierrez, B.L. Mejia, N.F. Perez, C.N. Zanuzzi and C. Barbeito, 2014. Paneth cell identification in the small intestine of guinea pig offsprings (Cavia porcellus). Anatom. Rec., 297: 856-863.
CrossRefDirect Link - Pfoze, K. and T.N. Singh, 2013. Histogenesis of Paneth cells in the human small intestine. J. Evol. Med. Dental Sci., 2: 3051-3054.
Direct Link - Barker, N., S. Bartfeld and H. Clevers, 2010. Tissue-resident adult stem cell populations of rapidly self-renewing organs. Cell Stem Cell, 7: 656-670.
CrossRefDirect Link - Van der Flier, L.G., A. Haegebarth, D.E. Stange, M. van de Wetering and H. Clevers, 2009. OLFM4 is a robust marker for stem cells in human intestine and marks a subset of colorectal cancer cells. Gastroenterology, 137: 15-17.
CrossRefPubMedDirect Link - Cheng, H. and C.P. Leblond, 1974. Origin, differentiation and renewal of the four main epithelial cell types in the mouse small intestine I. Columnar cell. Am. J. Anatomy, 141: 461-479.
CrossRefDirect Link - Cheng, H. and C. P. Leblond, 1974. Origin, differentiation and renewal of the four main epithelial cell types in the mouse small intestine V. Unitarian theory of the origin of the four epithelial cell types. Am. J. Anatomy, 141: 537-561.
CrossRefPubMedDirect Link - Potten, C.S., C. Booth, G.L. Tudor, D. Booth and G. Brady et al., 2003. Identification of a putative intestinal stem cell and early lineage marker; musashi-1. Differentiation, 71: 28-41.
CrossRefDirect Link - Sato, T., R.G. Vries, H.J. Snippert, M. van de Wetering and N. Barker et al., 2009. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature, 459: 262-265.
CrossRefDirect Link - Barker, N., A. van Oudenaarden and H. Clevers, 2012. Identifying the stem cell of the intestinal crypt: Strategies and pitfalls. Cell Stem Cell, 11: 452-460.
CrossRefDirect Link - Quyn, A.J., P.L. Appleton, F.A. Carey, R.J.C. Steele and N. Barker et al., 2010. Spindle orientation bias in gut epithelial stem cell compartments is lost in precancerous tissue. Cell Stem Cell, 6: 175-181.
CrossRefDirect Link - Ramachandran, A., M. Madesh and K.A. Balasubramanian, 2000. Apoptosis in the intestinal epithelium: Its relevance in normal and pathophysiological conditions. J. Gastroenterol. Hepatol., 15: 109-120.
CrossRefDirect Link - Dash, P.R., J.E. Cartwright and G.S.J. Whitley, 2003. Nitric oxide inhibits polyamine-induced apoptosis in the human extravillous trophoblast cell line SGHPL-4. Hum. Reprod., 18: 959-968.
CrossRefDirect Link - Zhu, Y., Y.F. Huang, C. Kek and D.V. Bulavin, 2013. Apoptosis differently affects lineage tracing of Lgr5 and Bmi1 intestinal stem cell populations. Cell Stem Cell, 12: 298-303.
CrossRefDirect Link