
Abdelghafar M. Abu-Elsaoud
Department of Botany, Faculty of Science, Suez Canal University, Ismailia-41522, Egypt
Sultan T. Tuleukhanov
Department of Biophysics and Physiology, Al-Faraby Kazakh National University, Almaty, Kazakhstan
Science International
Year: 2013 | Volume: 1 | Issue: 3 | Page No.: 39-50







ABSTRACT
Recently the biostimulatory role of physical methods have been observed in major agricultural crop plants, He-Ne laser is one of the most significant biophysical method for enhancing not only seed germination but also plant growth and development. Seeds of wheat cultivars from Kazakhstan (cultivars; aksay, kazakhstanskaya-10 and eretrospermum-350) and from Egypt (cultivar; sakha-168) were exposed to He-Ne laser irradiations with 5 mW.mm-2 power intensity for 1 to 1800 sec exposition time. Various germination and growth parameters and indices were recorded and calculated. Changes on the rate of superoxide dismutase activity, lipid peroxidation level and hydrogen peroxide accumulation were monitored. Wheat seed pre-treatment caused significant increase in various germination parameters including germination percentage, germination rate (seed h-1), relative germination rate coefficient (Wk) and enhancing various growth parameters. Several doses of He-Ne laser promoted the germination to a maximum of 100% germination, increasing also the germination rate. Moreover, seed pretreatment with different doses of He-Ne laser caused significant changes in levels of hydrogen peroxide, lipid peroxidation and Cu-Zn superoxide dismutase activity. Hydrogen peroxide accumulation and lipid peroxidation levels increased dramatically with the increasing doses of helium neon laser compared to control. However Cu-Zn superoxide dismutase activities dropped following seed pretreatment with He-Ne laser compared to control.
PDF Abstract XML References Citation
How to cite this article
Abdelghafar M. Abu-Elsaoud and Sultan T. Tuleukhanov, 2013. Can He-Ne Laser Induce Changes in Oxidative Stress and Antioxidant Activities of Wheat Cultivars from Kazakhstan and Egypt?. Science International, 1: 39-50.
URL: https://scialert.net/abstract/?doi=sciintl.2013.39.50
URL: https://scialert.net/abstract/?doi=sciintl.2013.39.50
INTRODUCTION
The increasing needs of agricultural products from different ecological zones for food production as well as different food industry requires probing innovative and safer methods to boost the agricultural productivity to meet the demand of increasing human population4. The use of various chemicals methods for improving plant productivity have been changing the properties of soil, water and atmosphere which in turn led to unfavorable conditions for growth and development of plants. The bio-physico-stimulatory methods have been observed in different agronomical characters of major crop plants and are being used in single or combination of physical factors5. The effect of low-intensity laser irradiation on the chemical composition and structure of lipids in wheat tissue culture were extensively studied by Salyaev36.
A comprehensive review on the clinical role of low intensity laser therapy (laser photo-stimulation) in biology and medicine were prepared focusing mainly on wound healing and pain relief which are mainly highlighted to show the clinical efficacy of laser therapy34. Controversies about the use of low intensity laser as a therapeutic modality for wound healing and pain relief are presented and a brief explanation is provided to overcome these controversies34. The importance of standard parameters is emphasized for the applications of low intensity lasers in biology and medicine. A justification has been made to warrant further research on the use of low intensity laser as a therapeutic modality. Although the therapeutic applications of low intensity laser are imminent, the heterogeneity in treatment protocols and study design calls for a vigilant interpretation of the findings34.
The effect of pulsed nitrogen laser radiation (337.1 nm) on morphological characteristics and biochemical contents in seedlings from treated green-gram (Vigna radiata L.) seeds were studied, seeds were germinated and grown in petri dishes for a week17. The shoot and root lengths and fresh and dry weights of the seedlings were maximum with the 30 min exposure, while protein was maximum with 20 min, RNA and DNA contents with 5 min exposure time. Chlorophyll content was not affected by the irradiation17.
The effect of low-intensity laser radiation on accumulation of lipid peroxidation secondary products was studied in wheat (Triticum aestivum L.) tissue culture35. A five-min-long callus irradiation by the helium-neon laser light with the wavelength ? = 632.8 nm and the intensity of 10 mW cm–2 resulted in an increase in the accumulation of the products of reaction with thiobarbituric acid (TBA-reactive products). The effect was less pronounced within two days after laser treatment, but even in this case the content of TBA-reactive products was greater than in the control. The data obtained confirm that the low-intensity laser radiation can induce lipid peroxidation processes in plant tissues35.
The aim of the current study was to examine the comparative effect of He-Ne laser irradiated seeds of wheat (Triticum aestivum L.) cultivars from both Egypt and Kazakhstan on in vitro germination, growth, physiological and biochemical parameters.
MATERIALS AND METHODS
Plant materials: Triticum aestivum L. seeds were obtained from the botanical institute (Kazakhstan) and the National Agricultural Centre, Egypt. Generally, four wheat cultivars were selected: three from Kazakhstan namely; Triticum aestivum L. cv. Aksay, Triticum aestivum L. cv. Kazakhstanskaya-10, Triticum aestivum L. cv. Eretrospermum-350 and one from Egypt namely; Triticum aestivum L. cv. Sakha-168.
Pre-sowing irradiation experiments: Irradiation at wavelength 632.8 nm was obtained from the He-Ne laser (Model LG-79-1, Russian Federation) with a total maximum output power of 5.23 mW cm–2 and beam diameter of 1.5 mm. Irradiation was performed at room temperature without thermostatic control. Detailed experimental conditions were presented in Table 1.
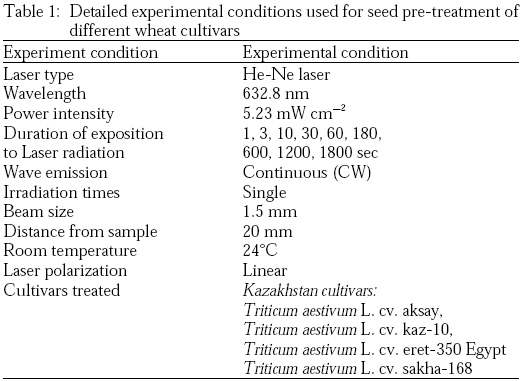 |
Seeds of the four selected cultivars from Kazakhstan and Egypt were rinsed in tap water and then dipped into a sodium hypochlorite solution (1% v/v) for sterilization. The arrangement of the seeds followed the following requirements: seeds were placed in single monolayer. Each seed was irradiated once before sowing. It should be emphasized, that the exposure times and irradiation powers used in our experiments were insufficient to cause any thermal effects in seeds28.
The irradiation process was carried out for each of the four selected wheat cultivars (Aksay, Kazakhstanskaya-10, Eretrospermum-350 and Sakha-168) with helium neon laser as irradiation source. Each seed was irradiated once before sowing in hydrogel with the proposed irradiation doses: 1, 3, 10, 30, 60, 180, 600, 1200 and 1800 sec (s). Wheat seedlings were harvested fifteen days after sowing and measurements of germination, morphological and physiological parameters were performed. Seeds were sown on specialy hydrogels, which are three-dimensional swollen networked structures4. When placed in an aqueous medium, hydrogels swell and retain the volume of the adsorbed aqueous medium in a three-dimensional swollen network of hydrophilic homopolymers or copolymers covalently or ionically cross-linked4,6,10,13. Such aqueous gel networks are also known as aquagels4. The original polymeric hydrogel network was developed by Wichterle and Lim in Czechoslovakia in 195414. In our experiments, the capacity of hydrogels to absorb the aqueous media was exploited to supply wheat cultivars with continuous nourishment of water and salts required by the plant.
Germination parameters: Standard germination data were recorded four times after 3, 5, 7 and 9 days following seed-sowing and various indices were calculated following Ciupak9. Based on the results obtained, the percentage of germinated seeds (Nk) was calculated by the following formula:
Where:
| nk | = | No. of germinated seeds |
| nc | = | Total No. of seeds sown |
Number of germinated seeds (nk) was expressed as the absolute number of germinated seeds and is presented as such in all Figures and Tables.
The germination rate Sk(seed/h) of wheat was calculated by the following equation:-
Where:
| nmax | = | Maximum number of germinated seeds recorded during one count |
| ?t | = | Time interval between two successive counts |
The relative germination rate coefficient Wk of various treated and untreated wheat cultivars was determined by the following equation:
Where
| n(t) | = | No. of treated seeds germinated in time t |
| ncontrol | = | No. of untreated seeds germinated in given time t |
Hydrogen Peroxide (H2O2) level: To determine Hydrogen Peroxide (H2O2) concentration in leaves following irradiation with electromagnetic radiations, the FOX method was employed20,43. Based on the peroxide-mediated oxidation of Fe2+, followed by the reaction of Fe3+ with xylenol orange (o-cresol-sulfonephthalein 3', 3''-bis [methylimino] diacetic acid, sodium salt; Farmitalia Carlo Erba, Milan, Italy), the FOX method accurately measures H2O2.
Lipid peroxidation (LP) level: Estimation of lipid peroxidation (LP) was obtained spectrophotometrically using the thiobarbituric acid-malondialdehyde (TBA-MDA) assay. The most prominent and currently used, assay as an index for lipid peroxidation products is the thiobarbituric acid assay (TBA test). Based on the reactivity of a lipid peroxidation end product, malondialdehyde (MDA) acts as a biological marker when it interacts with TBA to produce a quantifiable red adduct41. Lipid peroxides were extracted with 5 mL of 5% (w/v) metaphosphoric acid and 100 FL of 2% (w/v in ethanol) butyle hydroxytoluene. An aliquot of the supernatant was allowed to react with TBA at 95°C for 45 min and then cooled to room temperature. The resulting TBA-MDA was extracted with 1-butanol and detected spectrophotometrically using a Shimadzu UV-160 scanning spectrophotometer.
Antioxidant enzymes activity: Superoxide dismutase (SOD) activity was measured by the photochemical method described by Winterbourn42. Cu/Zn-SOD was measured by the photochemical method as described by Giannopolitis and Ries15. Assays were carried out under illumination. One unit SOD activity was defined as the amount of enzyme required to cause 50% inhibition of the rate of p-Nitro Blue Tetrazolium Chloride (NBT) reaction at 560 nm.
Statistical analyses: Various statistical analyses were carried out including two-Way-Analysis-of-Variance (ANOVA) to assess variations among tested wheat cultivars and various He-Ne laser exposition doses. Significance was tested at 0.05 level and is give asterisk (*) denoting the significance. Statistical analyses were performed using SPSS statistical software package (SPSS Inc., Version 15.00) and Microsoft Excel professional 2007 (a part of Microsoft Office Professional 2007 Package).
RESULTS
The overall biophysical stimulation effect of helium neon laser on germination was clearly shown by the germination percentage calculated after 3, 5 and 7 days following seed sowing, calculated germination percentage after seven days were presented on Table 2. He-Ne laser pre-treatment clearly induced significant changes in various germination indices of the four wheat cultivars either from Kazakhstan and Egypt, significances were assessed by two-way analysis of variance (ANOVA). A maximum of 100% germination percentage were recorded in various He-Ne laser treatments including 1, 3 and 60 sec (Table 2). Cultivars Kaz-10 showed the highest response to He-Ne laser giving 100% germination percentage after 1s He-Ne laser irradiation. While after 3s He-Ne irradiation, all three Kazakhstan cultivars showed maximum germination percentage. On the other hand, while the Egyptian cultivar Sakha-168 attained minimum germination (23.3%) (Table 2). Generally, seed pre-treatment with low doses of He-Ne laser of power intensity 5.23 mW cm–2 significantly enhanced the germination percentage of wheat cultivars, especially after 1, 3, 60 and 600 sec, while irradiation with 10, 180, 1200 and 1800 sec reduced the germination percentage of all wheat cultivars examined.
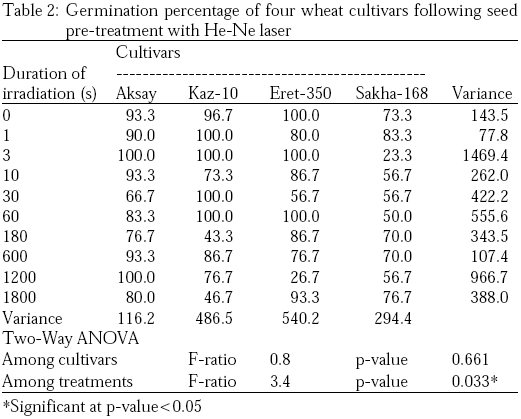 |
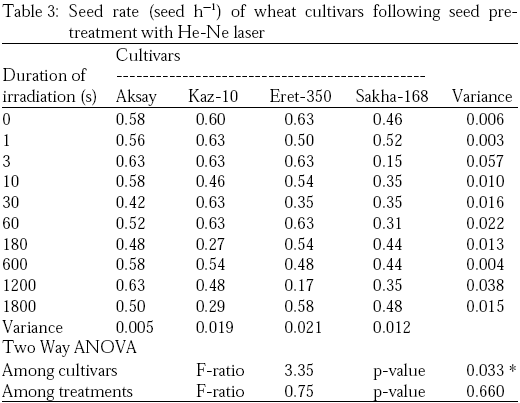 |
Germination rate expressed as seed h–1 for both control and irradiated seeds for the four wheat cultivars showed a general irradiation rate around 0.6 seed h–1 i.e., one seed every 100 min. Minimum germination rate of 0.15 seed h–1 was recorded in the Egyptian cultivar Sakha-168 after 3 sec He-Ne laser irradiation. Meanwhile, a maximum germination rate of 0.63 seed h–1 was recorded in cultivar Kaz-10 after irradiation with 1, 3, 30 and 60 sec He-Ne laser irradiations (Table 3).
Table 2 shows the relative germination rate coefficient (Wk) of both control and irradiated seeds. Two-way ANOVA revealed that there were slight significant differences among the studied cultivars in terms of Wk (F=3.4, p=0.064) and significant differences among time intervals after seed sowing (F=4.4, p=0.035*). Statistical analyses applied to the relative germination coefficients of wheat cultivars irradiated with He-Ne laser for 3 sec showed that: there was a high significant difference among cultivars (F=14.96, p=0.001*) and a non-significant difference among time intervals (F=0.90, p=0.478). Wheat cultivars irradiated for 10s with He-Ne laser showed no variations among the four cultivars, in terms of Wk. In contrast, highly significant changes in Wk with time were recorded (F=8.43, p=0.005*). Irradiation with 10 sec He-Ne laser didn’t actually increase the germination of the four cultivars, i.e., the maximum Wk did not exceed 1, but it did reduce the germination of the four wheat cultivar. Wheat cultivars irradiated with 30 sec of He-Ne laser showed no variations among time intervals in terms of Wk (F=1.27, p=0.340). In contrast, highly significant changes in Wk among studied cultivars were observed (F=8.523, p=0.005*). Maximum relative germination coefficient of 1.11 was recorded in cultivar Kaz-10 after 120 h after sowing. The Kazakhstan cultivar Eret-350 exhibited the highest values of relative germination intervals with time followed by Kaz-10. Maximum relative germination coefficient of 1.62 was recorded in the Eret-350 after 72 h, while a minimum Wk of 0.33 was recorded in the Egyptian cultivar Sakha-168. After 180 and 600 sec He-Ne laser irradiation, Wk showed non-significant variation among neither cultivars or among time intervals. In contrast, two-way ANOVA revealed that Wk of the various monitored wheat cultivars after seed pre-treatment with 1200 and 1800 sec He-Ne laser yielded very highly significant variations among the four wheat cultivars (F=21.17, p=0.000*; and F=42.97, p=0.000*; respectively).
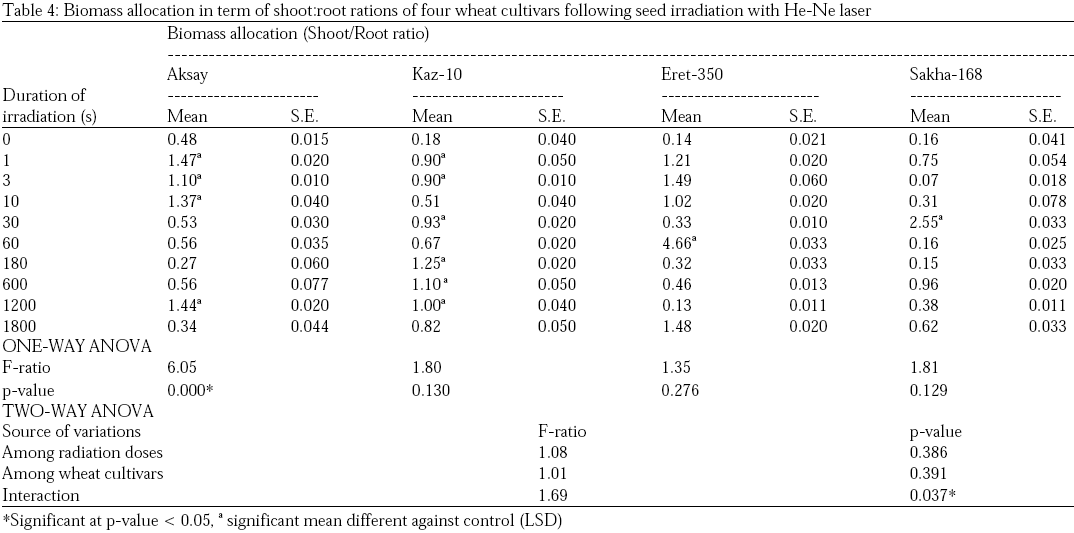
Shoot and root lengths (cm) in both control and treated plants with He-Ne laser radiations were measured and monitored after 3, 5, 7, 9 and 11 days following seed sowing. The relationship between He-Ne irradiation dose and shoot or root length were assessed using simple linear regression. A significant positive trend in shoot lengths was observed on the four tested wheat cultivars after irradiation with He-Ne laser. However, root length in two of the wheat cultivars studied showed a highly significant strong positive response to increased He-Ne laser radiation doses (namely, Kazakhstan cultivar Aksay and Egyptian cultivar Sakha-168), while, results of root lengths of the other two wheat cultivars (Kaz-10 and Eret-350) showed highly significant negative relationships with irradiation doses. Biomass allocation presented here was calculated in terms of shoot:root ratios, gives an accurate estimate of how plants respond to He-Ne laser by directing biomass toward root or shoot systems. Table 4 present the biomass allocation (shoot:root ratios) of the both control and treated wheat cultivars. Wheat cultivar Aksay exhibited biomass allocation toward the shoot system after irradiation with 1, 3 and 10 sec of He-Ne laser. It is important to note that the higher the irradiation time (>10 sec) the higher the allocation bias tended toward the root system. Cultivars Kaz-10, Eret-350 and Sakha-168 showed to some extent no differences in biomass allocation from control samples (Table 4).
Here, it was reported that the application of low intensity He-Ne laser with power intensity 5.23 mW cm–2 slightly enhance the production level of H2O2 (Fig. 1). The hydrogen peroxide level in wheat leaves of plants pre-treated with He-Ne laser increased with the influence of He-Ne laser. However, statistical assessment for the four wheat cultivars by ANOVA concluded this increase to be non-significant (Aksay: F=1.47, p=0.224; Kaz-10: F=1.76, p=0.141; Eret-350: F=1.30, p=0.296 and Sakha-168: F=1.55, p=1.96). Lipid Peroxidation (LP) refers to the oxidative degradation of lipids. It is the process whereby free radicals "steal" electrons from lipids in cell membranes, resulting in cell damage. This process proceeds by a free radical chain reaction mechanism, whereby increases in levels of LP become an indirect indicator of membrane and cellular damage.
 |
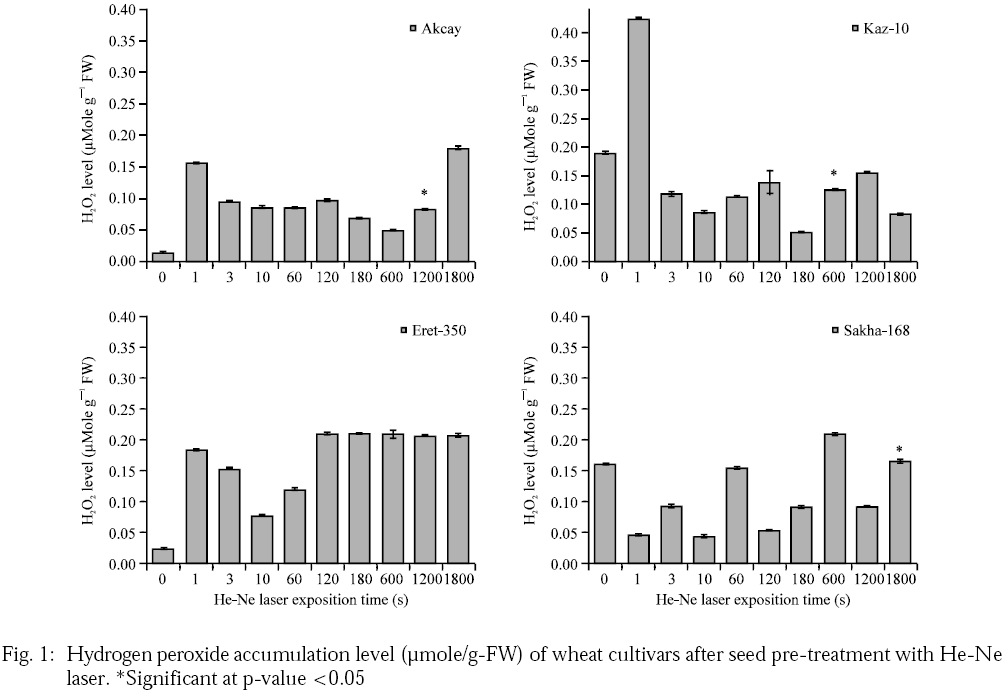 |
He-Ne seed pre-sowing irradiation caused statistically significant increased levels of LP to be detected in all four wheat cultivars (Fig. 2). These relationships and variations were subjected to two-way ANOVA analysis and least significant post-hoc test statistics (Aksay: F=92.8, p=0.000*; Kaz-10: F=332.9, p=0.000*; Eret-350 F=231.5, p=0.000*** and Sakha-168: F=2749.4, p=0.000*).
 |
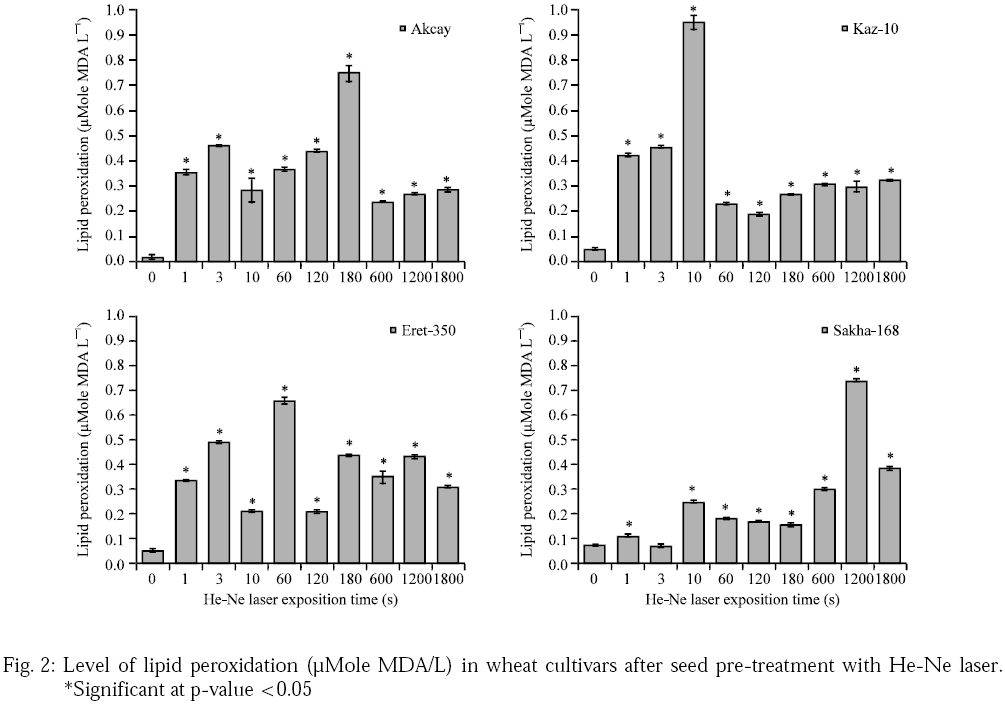 |
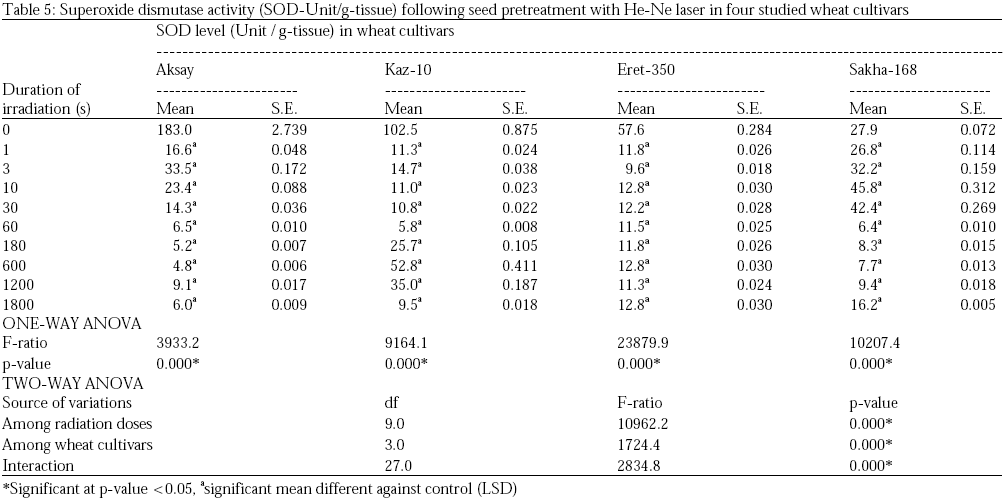
Superoxide dismutase (SOD) enzyme catalyses the dismutation of superoxide into oxygen and hydrogen peroxide. As such, it is an important antioxidant defence in nearly all cells exposed to oxygen. The role of superoxide dismutase is to act as a free radical scavenger preventing oxidative damage23,48. In our study, the SOD activities detected were clearly decreased after pre-sowing seed irradiation with He-Ne laser when compared to SOD enzyme activities in tissues of controlled plants in all studied cultivars (Aksay, Kaz-10, Eret-350 and Sakha-168). Furthermore, SOD activities recorded appeared in inverse proportion to increased doses of He-Ne laser (Table 5).
Two-way ANOVA of superoxide dismutase activities revealed that there are highly significant variations among cultivars (F=1724, p=0.000*) and among radiation doses based on SOD activity (F=10962, p=0.000*). Post-hoc least significant difference test revealed that there are highly significant difference in the SOD activity detected following He-Ne laser radiation doses as opposed to those detected in untreated controls in wheat Triticum aestivum cvs. Aksay, Kaz-10, Eret-350 and Sakha-168 (Table 5). Generally superoxide dismutase activities dropped following seed pre-treatment with He-Ne laser.
DISCUSSION
Results of the present study indicated that He-Ne laser triggers increases in various germination, morphological and growth parameters of the four wheat cultivars under examination in addition to various biochemical and physiological parameters. Results were consistent with those published by Govil18, Kutomkina24, Dinoev38, Muthusamy5 and many others. Low-power laser effects are considered to be photobiological phenomenon24. Reaction cascades begin with absorption of light in the mitochondria causing the signal to be transduced to the cellular membrane and from the membrane to the nucleus24. Whether this is the pathway followed resulting in germination is not yet clear. The components of the respiratory chain (cytochromes and flavoproteins) are characterized by their spectroscopic absorbance of light at ?=632.8 nm and behave as primary photoacceptors24. Here, again, it should be noted that the molecular mechanisms of germination are not fully understood24. In favour of light-stimulated action upon respiratory chain components remains that, as a rule, germination is accompanied by an abrupt increase in ATP content24, second some chemical compounds known to activate germination have been reported as capable of stimulating the endogenous activities of the electron transport system. Under large doses of laser radiation, its positive action changes, as a rule, resulting in inhibition of vital activity processes and becomes the main hindrance to a successful application of laser therapy and remains a cause of disappointment.
The effects of laser irradiation on an organism are principally attributed to light, electromagnetism, temperature and pressure. However, low power lasers (especially, the visible wavelength laser) are supposed to emit very little heat and pressure effect. Therefore, some researchers believe that the influential mechanism of laser irradiation is most likely its light effect and electromagnetism effect45. Still others believe the influence is due to the laser’s inherent biostimulation effect. Although biostimulation is frequently attributed to photochemical interactions, this has yet to be scientifically ascertained or validated. In most cases, wavelengths in the visible range (e.g., lasers at 632.8 nm) are used because of their efficiency and great optical penetration depths; the latter being critcally important when deeper tissue structures are to be reached27. Podlesny and Podlesna31 reported that laser irradiation of sowing pea and maize seeds positively influenced a-amylase activity and the concentration of free radicals, in addition to increasing the activities of some phyto-hormones, mainly indole-3-acetic acid (IAA)32. These reported influences, of laser on seeds, often manifested in the later periods of ontogenesis and resulted in plant maturation quickening.
Various processes of plants growth and development are modulated by internal cues, such as hormonal balance. However, plants are also particularly sensitive to their light environment and respond to external factors. Plant phytochrome are sensitive to red (Pr) and far-red (Pfr) light. Their red-light-absorbing form (Pr) is synthesized in the dark and gets converted into the far-red absorbing form (Pfr) upon absorption of red light. Inversely, far-red light will convert Pfr back into Pr. Given that many physiological responses correlate with the amount of Pfr, phytochromes can be regarded as light-regulated switches or botanical rheostats37. The laser, as a specific light, can be absorbed effectively by macromolecules to cause a variety of photochemical effects44 laser irradiation can boost the activities of SOD, APX and CAT32,33,25,3.
The results from our experiments demonstrated clearly that not only the parameters of germination seeds were increased greatly, but also the physiological metabolism and the growth and development of seedlings were significantly accelerated in response to He–Ne laser irradiation. Therefore, the role of laser pre-treatment is classified as a long-term effect. The current knowledge and published literature throughout the field of plant laser irradiation allows us to postulate on potential mechanisms underlying this effect. (1) The He–Ne laser radiation wavelength of 632.8 nm is close to the absorbing wavelength of Pr, making induction of the phytochrome very likely. The activities of related enzymes modulated by phytochromes may be subsequently enhanced and result in phytochrome-mediated responses accelerating (e.g., the decomposition rate of lower entropy macromolecules). Accordingly, the entropy and internal energy of seeds were enhanced during germination. The seed’s inherent high order kinetic equilibrium was broken by laser irradiation and a disordered status resulted. (2) Seeds pre-treated with laser have to absorb more energy from their surroundings than then do the untreated controls throughout the course of their individual development due to the laser-driven disruption of kinetic equilibrium in germinating seeds and enhanced internal energy of seeds7. The living organism, as an open-system, must exchange energy with surrounding system(s) in order to keep its high order energy state to compensate for this order being broken46. Consequently, the biochemical and physiological metabolic processes of plants pre-treated with laser were accelerated, resulting in enhanced growth and notably augmented leaf area and biomass7.
Manteifel26 stated that irradiation with 632.8 nm He-Ne-laser resulted in increased numbers of mitochondria per cellular section (20%), but not increased total area of mitochondria per cell section. The latter finding indicated modification of space configuration of the mitochondria in the absence of any accompanying changes in their mass. The increased quantity of mitochondria per cellular section following irradiation could be related to increases in the electrochemical proton gradient and/or phosphorylation activity. He-Ne-laser radiation and mitogen PHA cause mitochondrial de-aggregation to some extent (more pronounced in the case of PHA) and further supports a role for functional activation effects26.
Mitochondrial respiration in the absence of light is partially suppressed by nitric oxide synthesized by mithochondrial NO-synthase. Nitric oxide is known to inhibit respiration by binding to electron carriers, such as cytochromes, cytochrome oxidase and, possibly, iron-sulfur complexes. Intense light irradiation brings about the photolysis of these complexes to restore respiration and ATP synthesis. The beneficial actions produced by low intensity laser light, as well as light from other sources, involve numerous and different photochemical reactions39.
There are a great number of hypotheses concerning possible mechanisms of laser radiations. Vladimirov39 divided these hypotheses into two main categories: 1) Hypotheses based upon the idea of a specific action of coherent (laser) radiation on tissues, biological structures as a whole, water structure, etc. and 2) Hypotheses involving the photochemical action of light, including the radiation of lasers, LEDs and other sources of visible and near infrared light. Karu22 noted the following hypotheses concerning the mechanism of action of laser radiation. First, does the “singlet oxygen” hypothesis (1981), according to which the light-absorbing molecules such as porphyrins and flavoproteins can be changed, for example, in the respiratory chain of mitochondria into derivatives possess the properties of photosensitizers16. Under the action of light, these compounds evolve singlet oxygen that can stimulate, in turn, such processes as the synthesis of RNA and DNA. The authors of the hypothesis found corroboration for this idea in the fact that the spectra of activation of the synthesis of those compounds in HeLa cell cultures contained peaks that could be ascribed to porphyrins and flavin compounds22. The possibility of formation of singlet oxygen under radiation is also considered in paper of Zakharov47.
On the other hand, according to Chen7 and Yan and Zhan46 seeds pre-treated with laser have to absorb more energy from their surroundings than do the controls in the course of the individual development because the laser energy broke the kinetic equilibrium of germination seeds and enhanced the internal energy of seeds. It was reported that as an open system, the living organism must exchange energy with surrounding systems to keep its high order state of the system when this order is broken.
Oxidative stress was monitored in terms of lipid peroxidation (LP) and hydrogen peroxide (H2O2) content. Irradiation with He-Ne laser caused a significant increase in H2O2 content in the four wheat cultivars. Lipid peroxidation levels were decreased in cultivars Aksay and Kaz-10 after irradiation with He-Ne laser.
He-Ne laser irradiation is reported to significantly decrease malondialdehyde (MDA) content and superoxide radical (O2–•) production rate48. While, Mileva29 found that He-Ne laser treatment led to a slightly decreased level of LP products, they concluded that the effect of low-intensity laser-irradiation may depend on the dose applied, individual tissues targeted and other factors.
Considering our data in scope with the published literature, our experimental results strongly suggest that low-intensity laser radiation produces a stimulating influence on various physiological processes, including those that do not show a pronounced interconnection. For example, the lack of an interaction between the degree of plant regeneration and callus formation in individual genotypes confirms a lack of connection between the genetic factors that determine these two characteristics12.
The primary response of plant tissue to radiation is an increase in the content of lipid peroxidation products of peroxide oxidation. Data demonstrated that laser light stimulates morphogenetic processes in plant tissues at later stages, as well and believing that this stimulation may be conditioned metabolic changes caused by the change of content of a number of compounds formed as a result of the primary photoreactions12,37. Such compounds might also include products of peroxide oxidation, with an increase in their amounts as response to the impact of laser radiation. This increase, in turn, affects membrane properties and changes its functional state, a measurable result of which would be an increase in the Ca2+ concentration inside the cells12. The sequence of events following laser radiation exposure may occur as follows: 1) photon adsorption by endogenous photosensitizers and further peroxidation of lipids (photoperoxidation) followed by 2) calcium ion introduction into the cell. Activation of intracellular processes and/or secondary product accumulation may act as a signal not only for the start-up of relevant protection mechanisms, but, probably, for some secondary responses, perhaps even at the transcription level. This probability is indirectly confirmed by the stimulating effect of laser light on morphogenetic processes in wheat and wild crops tissue cultivar12.
Based on data from the stimulation of morphogenetic processes in wheat tissue culture by low-intensity laser radiation37, it was suggested that the changes observed should be accompanied by molecular shifts and structural reconstruction in those tissues subjected to radiation. Chirkova8 indicated that such reconstructions should take place primarily in cell and organellar membranes. These reconstructions produce a profound impact on all forms of functional activity in the membrane, particularly with lipids to a considerable extent. In lipids, fatty acids are the primary subject influenced by both genetic and environmental conditions. Stress may cause shifts in the proportion between various groups of fatty acids and the degree of their non-saturation may change. The length of fatty acids chains, positional situation of double bonds, or the number of polar groups may also change12.
Wasik40 concluded that the influence of laser irradiation on oxidative stress metabolites in living cells is physiologically dynamic and complicated. The authors outline three reactions by which this phenomena may be explained (i) photodynamic action on membranes accompanied by intracellular calcium increase and cell stimulation; (ii) photoreactivation of superoxide dismutase (SOD) protecting tissues from damage and (iii) photolysis of the metal complexes of NO with its release39. All of these effects may be related to fluctuations in intracellular calcium concentration. In mammalian cells, an increase of intracellular calcium is observed following irradiation of lymphocytes and granulocytes with He-Ne laser light39 which results, in turn, in cell activation and proliferation. Accordingly, biological consequences of increased Ca2+ concentration in the cytoplasm will differ depending on the type of cell and/or species exposed to laser radiation; however, it is generally accepted that laser radiation acts on mitochondria or even whole cells40,39,21,1. The results of the some studies suggest that more reactive oxygen species are generated in the electron transport system of the cell. Mitochondria are the major source of intracellular free radicals in all living cell types examined40,39,21,1,2,11.
In the present study, we elicited a stimulatory effect with laser light exposure, this action of laser irradiation is the result of initiation of primary, free radical reactions inducing activation of cells and also led to an increased mitotic index on the 3rd and 4th day after radiation with low doses. He-Ne irradiation acts through an increased content of intracellular calcium in mammalian lymphocytes39. Still other scientists indicate that cellular proliferation could be triggered by the action of the He-Ne laser40. Hu19 suggest the proliferation effect is due to mitochondrial photoacceptor-cytochrome c oxidase activity since He-Ne irradiation immediately induced an increase in mitochondrial membrane potential, ATP and cAMP via enhanced cytochrome-c oxidase activity. Under the action of light, singlet oxygen is released, which, in turn, can stimulate such processes as the synthesis of RNA and DNA40,39,21,1,2,11. Taken together, these observations suggest an overall improvement of respiratory chain function. Additional changes in mitochondria, like the increased levels of ATP in cells following irradiation, indicate the preparation of the cell for division39. It can be supposed that the sequence of events instigated via laser irradiation in this case is as follows: (i) absorption of a photon by an endogenous photosensitizer followed by lipid peroxidation; (ii) influx of calcium ions into the cells and (iii) activation of intracellular processes39.
In our experiments, antioxidant parameters measured in terms of superoxide dismutase SOD enzyme activities in response to seed pre-treatment with various doses of He-Ne radiation sources on four wheat cultivars from Kazakhstan and Egypt (Aksay, Kaz-10, Eret-350 and Sakha-168). Generally, radiation treatments caused significant decreases in SOD activity with the highest significance occurring in cultivars Aksay and Sakha-168.
Our result agreed with that of Zong-Bol48 who stated that He-Ne laser irradiation significantly decreased malondialdehyde (MDA) content and superoxide radical (O2–•) production rate and notably increased peroxidase (POD) enzyme activity and ascorbate (AsA) and glutathione (GSH) concentration in wheat seedlings exposed to moderate drought stress. Conversely, our results disagreed with Mileva30 who stated that the low-Intensity laser irradiation did not affect the oxidative stress in experimental cataract.
Various processes of growth and development in plants are modulated by internal cues such as the hormonal balance and external factors, whereas plants are particularly sensitive to their light environment7. The phytochromes are sensitive to red and far-red light. They are synthesized in their red-light-absorbing form (Pr) in the dark and are converted into their far-red absorbing form (Pfr) upon absorption of red light. Far-red light will convert Pfr back into Pr. Given that many physiological responses correlate with the amount of Pfr. Phytochromes can be regarded as light-regulated switches37. The laser, as a specific light, can be absorbed effectively by macromolecules and result in photochemical effects44. It has been reported that laser irradiation was capable of building up the activities of SOD, APX and CAT32,33.
Results of experiments carried out by Chen7 demonstrated clearly that not only the thermodynamic parameters of germination seeds were increased greatly but also the physiological metabolism and the growth and development of seedlings were significantly accelerated because seed stimulation with He–Ne laser radiation. Therefore, the role of laser pre-treatment appears to be a long-term effect. The underlying mechanism, according to them, might be: (1) the phytochrome was probably induced by the He–Ne laser radiation of 633 nm because its wavelength is close to the absorbing wavelength of Pr. Subsequently, the activities of related enzymes, which were modulated by phytochrome(s), could be enhanced and phytochrome-mediated responses, e.g. the decomposition rate of lower entropy macromolecules, are accelerated. Accordingly, the entropy and internal energy of seeds were enhanced during germination. As a result, kinetic equilibrium was broken by laser irradiation, which led to a status of disorder from high order of the organism’s system. The second mechanism may be seeds pre-treated with laser have to absorb more energy from their surroundings than their control counterparts in the course of the individual development because the laser broke the kinetic equilibrium of germination seeds and enhanced the internal energy of seeds. It was reported that as an open system, the living organism must exchange energy with surrounding systems in order to keep its high order state following this order being broken46,7.
REFERENCES
- Boveris, A. and E. Cadenas, 1975. Mitochondrial production of superoxide anions and its relationship to the antimycin insensitive respiration. FEBS Lett., 54: 311-314.
CrossRefDirect Link - Boveris, A. and B. Chance, 1973. The mitochondrial generation of hydrogen peroxide. General properties and effect of hyperbaric oxygen. Biochem. J., 134: 707-716.
PubMed - Muthusamy, A., P.P. Kudwa, V. Prabhu, K.K. Mahato and V.S. Babu et al., 2012. Influence of Helium-Neon laser irradiation on seed germination in vitro and physic-biochemical character in seedling of Brinjal (Solanum melongena L.) var. Mattu Gulla. Photochem. Photobiol., 8: 1227-1235.
CrossRefPubMedDirect Link - Chen, Y.P., M. Yue and X.L. Wang, 2005. Influence of He-Ne laser irradiation on seeds thermodynamic parameters and seedlings growth of Isatis indogotica. Plant Sci., 168: 601-606.
CrossRefDirect Link - Ciupak, A., I. Szczurowska, B. Gladyszewska and S. Pietruszewski, 2007. Impact of laser light and magnetic field stimulation on the process of buckwheat seed germination. Tech. Sci. 10: 1-10.
Direct Link - De Souza, A., D. Garci, L. Sueiro, F. Gilart, E. Porras and L. Licea, 2006. Pre-sowing magnetic treatments of tomato seeds increase the growth and yield of plants. Bioelectromagnetics, 27: 247-257.
CrossRef - Florez, M., M.V. Carbonell and E. Martinez, 2007. Exposure of maize seeds to stationary magnetic fields: Effects on germination and early growth. Environ. Exp. Bot., 59: 68-75.
CrossRef - Giannopolitis, C.N. and S.K. Ries, 1977. Superoxide dismutases. I. Occurrence in higher plants. Plant Physiol., 59: 309-314.
CrossRefPubMedDirect Link - Sudha, R.G., D.C. Agrawal, K.P. Rai and S.N. Thakur, 1985. Growth responses of Vigna radiata seeds to laser irradiation in the UV-A region. Physiol. Plant., 63: 133-134.
CrossRefDirect Link - Hu, W.P., J.J. Wang, C.L. Yu, C. Lan, G.S. Chen and H.S. Yu, 2007. Helium-neon laser irradiation stimulates cell proliferation through photostimulatory effects in mitochondria. J. Invest. Dermatol., 127: 2048-2057.
CrossRefPubMedDirect Link - Jiang, Z.Y., A.C.S. Woollard and S.P. Wolff, 1990. Hydrogen peroxide production during experimental protein glycation. FEBS Lett., 268: 69-71.
Direct Link - Klebanov, G., A.B. Kapitanov, Y. Tessikin, I.V. Babenkova and B.A. Zhambalova et al., 1998. The antioxidant properties of lycopene. Membr. Cell Biol., 12: 287-300.
PubMedDirect Link - Kutomkina, E.V., T.L. Karu, M.S. Mushegian and V.I. Duda, 1991. He-Ne laser induced germination of Endospores of Anaerobactor polyendosporus. Laser. Life Sci., 4: 147-151.
Direct Link - Manteifel, V.M., T.N. Andreichuk and T.I. Karu, 1991. The effect of irradiation by a He-Ne laser and phytohemagglutinin on lymphocyte mitochondria. Mol. Biol., 25: 273-280.
PubMedDirect Link - McKenzie, A.L., 1990. Physics of thermal processes in laser-tissue interaction. Phys. Med. Biol., 35: 1175-1209.
PubMedDirect Link - Mileva, M., G. Zlateva, S. Karabasheva, V. Hadjimitova and I. Antonov, 2000. Effect of He-Ne laser treatment on the level of lipid peroxidation products in experimental cataract of rabbit eyes. Meth. Findings Exp. Clin. Pharm., 22: 679-681.
PubMed - Mileva, M., R. Bakalova and G. Zlateva, 2004. The low-intensity laser irradiation does not affect the oxidative stress in experimental cataract. Med. Laser Applic., 19: 150-154.
CrossRefDirect Link - Podlesny, J. and A. Podlesna, 2004. Morphological changes and yield of selected species of leguminous plants under the influence of seed treatment with laser light. Int. Agrophysics, 18: 253-260.
Direct Link - Qi, Z., M. Yue and X.L. Wang, 2000. Laser pretreatment protects cells of broad bean from UV-B radiation damage. J. Photochem. Photobio. B Biol., 59: 33-37.
CrossRef - Reddy, G.K., 2004. Photobiological basis and clinical role of low-intensity lasers in biology and medicine. J. Clin. Laser Med Surg., 22: 141-150.
Direct Link - Salyaev, R.K., L.V. Dudarev, S.V. Lankevich, E.G. Ekimova and V.M. Sumtsova, 2003. Effect of low-intensity laser radiation on the lipid peroxidation in wheat callus culture. Russian J. Plant Physiol., 50: 498-500.
CrossRef - Salyaev, R.K., L. Dudareva, S. Lankevich, S. Makarenko, V. Sumtsova and E. Rudikovska, 2007. Effect of low-intensity laser irradiation on the chemical composition and structure of lipids in wheat tissue culture. Doklad. Biol. Sci., 412: 87-88.
CrossRef - Smith, H., 2000. Phytochromes and light signal perception by plants: An emerging synthesis. Nature, 407: 585-591.
CrossRef - Vladimirov, Y.A., A.N. Osipov and G.I. Klebanov, 2004. Photobiological principles of therapeutic applications of laser radiation. Biochemistry (Moscow), 69: 81-90.
PubMedDirect Link - Wasik, M., E. Gorska, M. Modzelewska, K. Nowicki, B. Jakubczak and U. Demkow, 2007. The influence of low-power helium-neon laser irradiation on function of selected peripheral blood cells. J. Physiol. Pharmacol., 58: 729-737.
PubMed - Winterbourn, C.C., R.E. Hawkins, M. Brian and R.W. Carrell, 1975. The estimation of red cell superoxide dismutase activity. J. Lab. Clin. Med., 85: 337-341.
PubMedDirect Link - Wolff, S.P., 1994. Ferrous ion oxidation in presence of ferric ion indicator xylenol orange for measurement of hydroperoxides. Methods Enzymol., 233: 182-189.
CrossRefDirect Link