
P.Y. Cavasin
Federal University of Lavras, Lavras, Aquenta Sol, 37200-900, Lavras, Minas Gerais, Brazil
H.O. Santos
Federal University of Lavras, Lavras, Aquenta Sol, 37200-900, Lavras, Minas Gerais, Brazil
L.A.A. Gomes
Federal University of Lavras, Lavras, Aquenta Sol, 37200-900, Lavras, Minas Gerais, Brazil
T.B. Fantazzini
Federal University of Lavras, Lavras, Aquenta Sol, 37200-900, Lavras, Minas Gerais, Brazil
E.V.R. Von-Pinho
Federal University of Lavras, Lavras, Aquenta Sol, 37200-900, Lavras, Minas Gerais, Brazil
R.M.O. Pires
Federal University of Lavras, Lavras, Aquenta Sol, 37200-900, Lavras, Minas Gerais, Brazil
E.R. Carvalho
Federal University of Lavras, Lavras, Aquenta Sol, 37200-900, Lavras, Minas Gerais, Brazil
Research Journal of Seed Science
Year: 2021 | Volume: 14 | Issue: 1 | Page No.: 11-18







ABSTRACT
Background and Objective: Lettuce seed germination at high temperatures is a problem faced by farmers and seed producers in Brazil. At high temperatures, germination is reduced or null and changes in physiology and the patterns of germination-related enzymes, such as endo-β-mannanase, can occur. The objective of this study was to determine the relationship between endo-β-mannanase activity and tolerance to thermoinhibition in lettuce seeds, as well as to determine the relationship between seeds and antioxidant enzymes. Materials and Methods: Seeds from two lettuce cultivars, Everglades (thermotolerant) and Veronica (thermosensitive), were used. The seeds were assessed for first count, germination and germination speed index at seven temperatures (20, 23, 25, 27, 30, 33 and 35°C). Additionally, the activity of the enzymes endo-β-mannanase, catalase, ascorbate peroxidase and superoxide dismutase and the levels of hydrogen peroxide were quantified in each cultivar at the different temperatures. The experimental design was completely randomized in a 2×7 factorial arrangement, with two cultivars and seven temperatures. Results: At temperatures above 30°C, the Everglades cultivar showed higher values than the Veronica cultivar, including for endo-β-mannanase activity, in all treatments. Additionally, the oxidation levels, as confirmed by the analysis of oxidative pathway enzymes, were higher in the Veronica cultivar, thus confirming that the cultivar is thermosensitive to temperatures above 25°C. Conclusion: It is concluded that high temperature directly affects the germination parameters and consequently the activity of endo-β-mannanase in lettuce seeds, especially in the thermosensitive cultivar Veronica.
PDF Abstract XML References Citation
How to cite this article
P.Y. Cavasin, H.O. Santos, L.A.A. Gomes, T.B. Fantazzini, E.V.R. Von-Pinho, R.M.O. Pires and E.R. Carvalho, 2021. Endo-β-Mannanase Enzyme Activity is Related to Thermo Inhibition in Lactuca sativa Seeds. Research Journal of Seed Science, 14: 11-18.
URL: https://scialert.net/abstract/?doi=rjss.2021.11.18
URL: https://scialert.net/abstract/?doi=rjss.2021.11.18
INTRODUCTION
Lettuce (Lactuca sativa L.) is one of the main leafy vegetables produced and consumed worldwide and it is grown in regions with different characteristics1. Despite the advances obtained by genetic improvement programmes over the years, lettuce cultivars are limited by the fact that their seeds are sensitive to variations in the moisture and temperature of the germination medium. When lettuce is sown in regions with high temperatures, there is a reduction in germination and vigour, causing delays in the production of seedlings and, consequently, a reduction in yield and direct losses for the producer2.
Temperature is the primary factor regulating seed germination, acting directly or indirectly on germination through dormancy and viability. In general, lettuce seeds show maximum germination at approximately 20°C and most cultivars do not germinate when exposed to temperatures above 30°C3.
High temperatures in the seed imbibition phase may lead to the occurrence of two distinct phenomena, known as thermoinhibition and thermodormancy, which lead to reduced seed germination and these mechanisms differ in their natural irreversibility4. Thermoinhibition is reversible when all factors promoting inhibition, such as temperature, light and humidity, become favourable. In turn, thermodormancy requires treatment for germination to occur, even under favourable conditions for germination5.
Although most lettuce cultivars produced are sensitive to thermoinhibition, there are reports in the literature of genotypes whose seeds can germinate at high temperatures. These genotypes include the butterhead lettuce cultivar Everglades, whose seed germination rate is higher than 70% at 35°C6,7.
The mechanisms of action affecting the germination of lettuce seeds at high temperatures seem to be related to the weakening of the endosperm, which allows the embryo to grow under these conditions2. For such weakening to occur, the activity of enzymes such as Alcohol Dehydrogenase (ADH), Malate Dehydrogenase (MDH), Catalase (CAT), Esterase (EST), Pyruvate Decarboxylase (PDC), Glutamate Oxaloacetate Transferase (GOT) and in particular, endo-β-mannanase, is essential. Endo-β-mannanase is one of the main enzymes responsible for the hydrolysis of mannans in the endosperm during lettuce seed germination6.
Regarding temperature sensitivity, thermosensitive lettuce cultivars have more mannose in the cell wall than do thermotolerant genotypes; higher amounts of mannose mean that a longer time is required for endo-β-mannanase to complete hydrolysis and therefore, the germination time is longer8,9.
In equatorial and tropical regions where temperatures of 30°C or more occur, lettuce cultivars with a higher limit temperature for germination must be developed7. Given the above, the objective of this study was to determine the relationship between tolerance to thermoinhibition in lettuce seeds and physiological changes in the Everglades and Veronica cultivars.
MATERIALS AND METHODS
Study area: The study was conducted at the seed department of the Federal University of Lavras (UFLA), Seed Analysis Laboratory, Brazil, from October, 2020-January, 2021.
Seed collection: Seeds from two lettuce cultivars, Everglades (thermo inhibition tolerant) and Veronica (thermo inhibition sensitive), were used. The cultivars used in the study were selected according to the results obtained by Catão et al.4 based on germination and seedling growth data under high-temperature conditions during the germination process.
Methodology: During plant growth, until the time of seed harvest, spraying, fertilization and irrigation were performed according to the needs of the crop uniformly. Seeds from both cultivars produced under the same conditions were harvested, sanitized and stored in a cold chamber at 8°C until testing.
Four replicates of 50 seeds were used to evaluate the first count (FC), percent germination (G) and Germination Speed Index (GSI). The seeds were distributed under two sheets of blotting paper in germination boxes. The papers were previously moistened with distilled water at a ratio of 2.5 times the weight of the paper. The germination boxes were placed in a BOD germination chamber at temperatures of 20, 23, 25, 27, 30, 33 and 35°C with a 12 hrs photoperiod. Germinated seeds were counted daily until the seventh day to determine the G and GSI values. Data from the FC test were obtained on the fourth day. The results were expressed as the percentage of normal seedlings, according to the ISTA (https://www. seedtest.org/en/ista-rules-2019-_content---1--3410.html).
Determination of endo-β-mannanase activity: For the determination of endo-β-mannanase activity, seeds of the two cultivars were soaked at different temperatures for 48 hrs under the same conditions mentioned. Above 48 hrs, the seeds were macerated in a crucible with liquid nitrogen and polyvinylpyrrolidone (PVP). To each 100 mg of powder from each sample, 300 μL of extraction buffer (0.1 M HEPES/0.5 M NaCl and ascorbic acid (5 mg ascorbic acid per mL buffer), pH 8.0) was added.
Then, the samples were centrifuged (30 min/14,000 rpm) and 20 μL was pipetted onto agarose gel containing 6 mL of locust bean gum (LBG; 0.24 g of agarose and 24 mL of 1 M citric acid/0.4 M NaHPO4·2H2O, pH 5.0). The gel was incubated for 21 hrs and developed according to the method proposed by Silva et al.10. The endo-β-mannanase activity was calculated according to Downie et al.11.
Determination of lipid peroxidation: Lipid peroxidation was determined by quantifying Malondialdehyde (MDA), as described by Hodges et al.12. For this analysis, 100 mg of seed fresh weight was macerated in liquid nitrogen and homogenized with 1 mL of 80% ethanol three times, totalling 3 mL. The reaction was performed with 1 mL of 20% Thiobarbituric Acid (TBA), 0.65% TBA and 0.01% Beta-hydroxytoluene (BHT). The estimated lipid peroxidation was obtained by absorbance reading in a spectrophotometer at 440 and 532 nm.
The enzymatic extracts were obtained following the protocol of Biemelt et al.13, where 200 mg of seedlings were macerated in liquid nitrogen supplemented with insoluble polyvinylpolypyrrolidone (PVPP), to which 1.5 mL of extraction buffer (400 mM potassium phosphate (pH 7.8), 10 mM EDTA and 200 mM ascorbic acid) was added. The homogenate was centrifuged at 13,000 g for 10 mins at 4°C and the collected supernatant was used to quantify the enzymatic activity.
Determination of superoxide dismutase: Superoxide Dismutase (SOD) activity was estimated by the ability of the enzyme to inhibit the photoreduction of Nitrotetrazolium Blue (NBT)14. The readings were performed at 560 nm. One unit of SOD corresponds to the amount of enzyme capable of inhibiting the photoreduction of NBT by 50% under the test conditions.
Determination of catalase: Catalase (CAT) activity was determined according to Havir and McHale15. The activity of this enzyme was determined by the decrease in absorbance at 240 nm every 15 sec for 3 min, monitored by the consumption of hydrogen peroxide. The molar extinction coefficient used was 36 mM cm–1, as described by Azevedo et al.16.
Determination of ascorbate peroxidase: Ascorbate Peroxidase (APX) activity was determined according to Nakano and Asada17 by monitoring the oxidation rate of ascorbate at 290 nm and the molar extinction coefficient used was 2.8 mM cm–1.
Physiological analysis: The experimental design used in the physiological seed quality and enzymatic activity tests was completely randomized in a 2×7 factorial arrangement, with the factors being the two lettuce cultivars (Everglades and Veronica) and the seven temperatures (20, 23, 25, 27, 30, 33 and 35°C).
Statistical analysis: All data were computed and the analysis of variance and Tukey’s means test (5% error probability) were performed using R for Windows (https://www.R-project.org/).
RESULTS
Both the thermotolerant cultivar (Everglades) and the thermosensitive cultivar (Veronica) showed higher values for FC (Table 1) and GSI (Table 2) at temperatures from 23-30°C than at other temperatures. The highest FC values for the Everglades cultivar were found at 27°C and the highest values for the Veronica cultivar were found at 25°C.
Both the Everglades and Veronica cultivars showed higher FC values at temperatures below 30°C (Table 1); at temperatures above 30°C, there was a decrease in FC, indicating thermoinhibition. At 33°C, the Everglades cultivar showed an FC of 53%, while the Veronica cultivar showed an FC of 4%. At 35°C, there was no germination in either cultivar.
Regarding GSI, there was a reduction in the values for both cultivars at temperatures above 30°C (Table 2). At 35°C, the GSI of the Veronica cultivar was 0.00 and that of the Everglades cultivar was 1.04.
At 30°C, the Everglades cultivar presented a G of 96%, significantly different from that (88%) of the Veronica cultivar (Table 3). With increasing temperature, there was a reduction in the germination rates of the cultivars. The Veronica cultivar did not germinate at 35°C.
The endo-β-mannanase activity of the two cultivars at different temperatures is shown in Fig. 1.The cultivars showed no significant differences at 20-27°C; however, there was a significant difference between the cultivars at 30°C. The Everglades cultivar showed higher endo-β-mannanase activity than the Veronica cultivar.
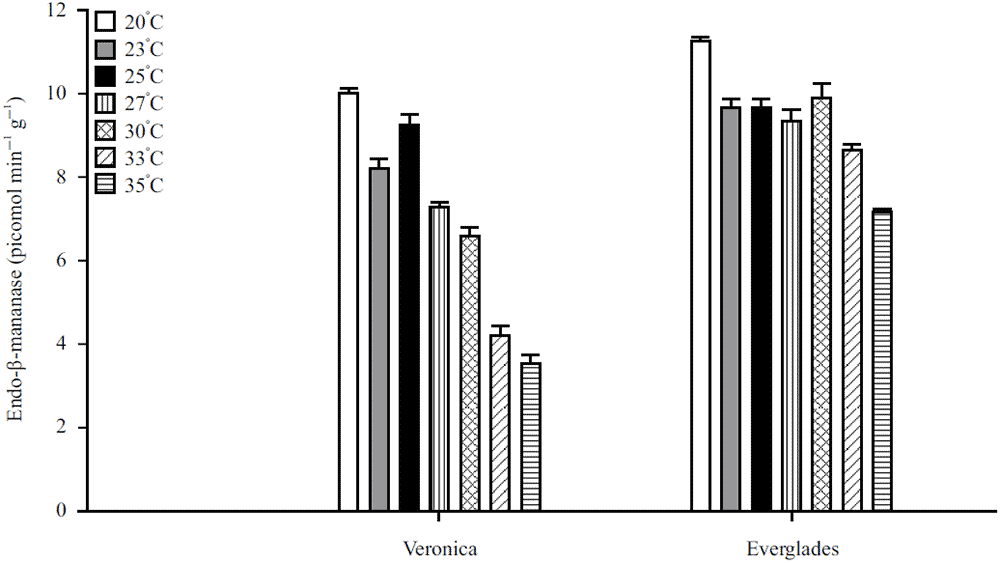 | |
| Fig. 1: | Endo-β-mannanase activity in seeds of the lettuce cultivars Veronica and Everglades subjected to different temperatures during germination |
| Table 1: | First germination count in seeds of the lettuce cultivars Everglades and Veronica at different temperatures |
 | |
| Means followed by the same uppercase letter in a column and the same lowercase letter in a row do not differ significantly by Tukey’s test at 5% probability | |
| Table 2: | Germination speed index in seeds of the lettuce cultivars Everglades and Veronica at different temperatures |
 | |
| Means followed by the same uppercase letter in a column and the same lowercase letter in a row do not differ significantly by Tukey’s test at 5% probability | |
| Table 3: | Germination of seeds of the lettuce cultivars Everglades and Veronica at different temperatures |
 | |
| Means followed by the same uppercase letter in a column and the same lowercase letter in a row do not differ significantly by Tukey’s test at 5% probability | |
Regarding lipid peroxidation and antioxidant system enzymes (MDA, SOD, CAT and APX), both cultivars had lower levels below 25°C and at higher temperatures, there was greater membrane damage and enzyme activation (Fig. 2a-d). The Everglades cultivar did not differ significantly from the Veronica cultivar at temperatures of 20 and 23°C; however, at other temperatures, the Everglades cultivar was more tolerant because the peroxidation and antioxidant activities were lower than those observed in the Veronica cultivar.
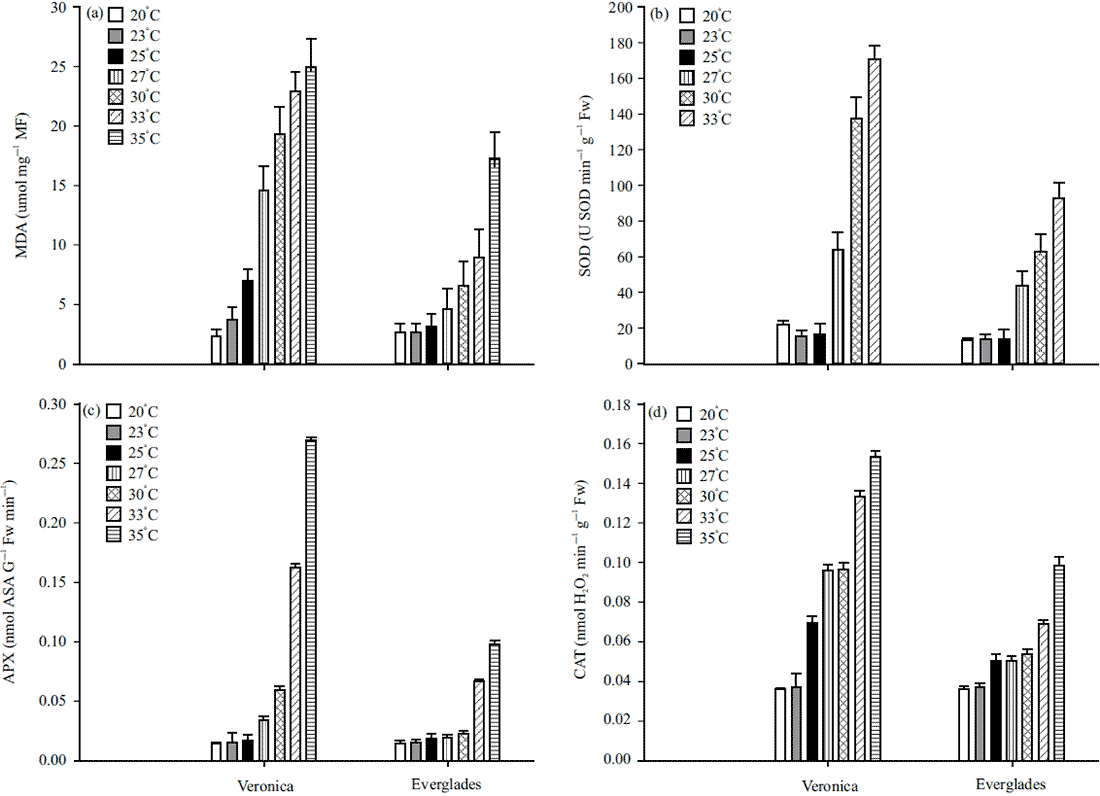 | |
| Fig. 2(a-d): | Enzymatic activity of Veronica and Everglades cultivars at different temperatures during germination |
| (a) Lipid peroxidation, (b) Superoxide dismutase enzyme activity, (c) Ascorbate peroxidase enzyme activity and (d) Catalase enzyme activity | |
DISCUSSION
Temperatures above 30°C interfere negatively in the activity of the endo-β-mannanase enzyme and consequently in the germination of lettuce seeds. Thus, thermotolerant cultivars have less influence on temperature than thermosensitive cultivars.
Temperature is inarguably the most important environmental factor governing the maximum germination percentage and germination rate, as it directly affects the biosynthesis of Abscisic Acid (ABA) and Gibberellin (GA) and the response to ethylene and other metabolites18. In addition, delayed or reduced germination due to given stress can be explained by the effects on the mobilization of reserves, which occurs in the early stages of seed germination, by the action of secondary metabolites19.
The lettuce seed embryo is surrounded by the endosperm, which consists of a layer of two to four cells. The endosperm can delay or prevent seed germination by acting as a physical barrier to radicle emission, especially under unfavourable conditions. Thus, the weakening (softening) of the endosperm is a prerequisite for radicle emission under high-temperature conditions20. Because the cell walls of the endosperm are composed mainly of polymers of galactomannans, endo-β-mannanase may potentially be a regulatory enzyme in the weakening of the endosperm. The ethylene hormone could also be involved in this process, directly acting on the weakening of the seed endosperm through an unknown mechanism; be involved in the regulation of enzymes responsible for digesting the endosperm cell wall (for example, endo-β-mannanase); or be involved in the combination of both. All these processes would favour radicle emission and consequently the germination of lettuce seeds under high temperatures.
Under high temperatures, ABA-related genes are more highly expressed and germination is inhibited in thermosensitive lettuce seeds; however, in thermotolerant seeds, genes related to ethylene and GA are more highly expressed and germination is allowed21.
To further detail the behaviour of endo-β-mannanase in lettuce, Wang et al.22 performed molecular characterization via cDNA isolation of this enzyme from the Grand Rapids cultivar. Transcription was observed only in germinated seeds (15 hrs after imbibition). Thus, it can be concluded that the synthesis of endo-β-mannanase does not occur before contact of the radicle with the endosperm. Furthermore, iso forms of this enzyme with differentiated activities were identified. This can affect the rate of action of endo-β-mannanase on the endosperm, influencing germination. The higher endo-β-mannanase activity was observed one hrs before root protrusion in the Dark Green Boston lettuce cultivar (thermosensitive) when the seeds were produced under controlled conditions at 30/20°C than when they were produced at 20/10°C23. In addition, there was attenuation of thermoinhibition in seed production at 30/20°C, regardless of whether the cultivar was thermosensitive or thermotolerant.
Germination reduction or inhibition is due to a series of biochemical reactions highly dependent on temperature. Thermoinhibition occurs by increasing the ABA biosynthesis pathway (NCED1 and NCED9), while the regulators of GA biosynthesis (GA3ox1 and GA20ox1) are altered at high temperatures24.
It is known that GA causes a relatively small increase in endo-β-mannanase activity, while ABA causes a large decrease. In tests with rice seeds in the presence of GA, there was an increase in the enzymatic activity of endo-β-mannanase. Furthermore, in the same study, GA was able to reverse the inhibitory effect of ABA on enzyme activity, indicating that thermotolerant seeds may have a greater content of GA, which would consequently increase the synthesis of endo-β-mannanase. The endo-β-mannanase enzyme acts on the hemicellulose fraction of the endosperm cell walls, especially mannans and galactomannans, weakening the cell wall and allowing root protrusion duringgermination25.
In palm seeds (Acrocomia aculeata), endo-β-mannanase and α-galactosidase immediately become active in germination, indicating the important role of enzymes in mobilizing storage reserves and weakening the endosperm, which in turn allows the expansion of the haustorium during seedling development26. The same process occurs in seeds of plants from the Brazilian Cerrado of the genus Comanthera (C. bisulcata and C. elegans), which showed higher
expression of endo-β-mannanase, consequently favouring radicle protrusion27. In Acrocomia aculeata (Arecaceae) seeds, the endosperm also plays an active role in germination and serves as a site of intense enzymatic activity28.
The activity of endo-β-mannanase can be influenced by different external factors, as reported by Albuquerque et al.8, who found that the addition of micronutrients, amino acids and growth regulators to lettuce seeds affects germination. As a confirmation of the influence of the environment, it was found that high temperatures associated with longer storage periods cause a reduction in germination and the enzymatic patterns of lettuce seeds7.
Endo-β-mannanase activity in Coffea arabica L. seeds is also modulated by different types of processing (drying, depulping and demucilage). Coffee seeds subjected to natural processing show greater endo-β-mannanase activity, whereas when the mucilage is removed during depulping and demucilage, endo-β-mannanase activity is reduced9.
The biochemical response of seeds is within what is known as stress tolerance. Tolerance involves osmotic adjustment and the synthesis or increased activity of enzymes and antioxidant molecules. Several studies report that under stress conditions, seeds trigger the production and accumulation of reactive O2 species (ROS), which can damage cell membranes and this damage results in an increased lipid peroxidation content, thus impairing cell metabolism29. Each ROS has specific characteristics and causes specific damage; for example, singlet oxygen is associated with cell membrane damage, superoxide is associated with protein damage and the hydroxyl radical is associated with membrane and DNA damage30.
To avoid ROS accumulation, plants have enzymatic defence systems that enable the elimination of these compounds and thus protect against oxidative stress. SOD activity is related to the production of superoxide and CAT and APX activity indicate high levels of hydrogen peroxide31. Even though there was an increase in the enzymatic activity of the antioxidant system in the lettuce seeds, it was not enough to eliminate the excess ROS, which was associated with increased lipid peroxidation.
SOD is considered the first line of defence in plants. This enzyme breaks down the superoxide ion into hydrogen peroxide (H2O2) and water. Once H2O2 is formed, other enzymes, such as CAT and APX, act to transform it into water. It is important to note that if H2O2 is not transformed into water, it can react with transition metals and give rise to the hydroxyl radical, which damages the plasma membrane, DNA and proteins, resulting in increased membrane fluidity, mutations and loss of protein functionality30.
The Everglades cultivar showed higher values for the different germination parameters than did the thermosensitive Veronica cultivar, demonstrating its thermotolerant character. Additionally, it is concluded that high temperatures have a direct influence on lettuce physiological parameters and consequently modulate endo-β-mannanase activity when the plants are exposed to temperatures above 30°C and this effect occurred to a greater degree in the thermosensitive cultivar.
CONCLUSION
This study investigated the activity of the endo-β-mannanase enzyme to temperature in the germination of Lactuca sativa L. It was found that thermotolerant lettuce seeds (Everglades) have higher enzyme activity compared to thermosensitive lettuce seeds (Veronica) and consequently better physiological parameters, which directly impact germination. Temperatures above 30°C interfere directly with the enzyme activity, however, cultivar Everglades maintains better % germination compared to cultivar Verônica.
SIGNIFICANCE STATEMENT
This study found that the enzyme endo-β-mannanase has greater activity in thermotolerant cultivars, even at high temperatures, confirming the potential of the cultivar Everglades. This study contributes to lettuce improvement programs aimed at high-temperature tolerance in germination.
ACKNOWLEDGMENTS
The authors thank the Research Foundation of the State of Minas Gerais (Fapemig), Brazilian Federal Agency for Support and Evaluation of Graduate Education (Capes) and the National Council for Scientific and Technological Development (CNPq) for the Doctorate Pedro 165293/2019-1 scholarship.
REFERENCES
- Rossi, E.M., R.R. Guzela, M. Kochhann, L.K. Menezes and J.F.B. Honorato, 2020. Consumer knowledge and efficiency of laundry and disinfection methods of lettuce (Lactuca sativa) commercialized in supermarkets in a southern city of Brazil. Braz. J. Food Technol., Vol. 23.
CrossRefDirect Link - Wang, L., J. Hao, Z. Qi, W. Liu, C. Liu, Y. Han and S. Fan, 2019. Cloning and expression of mitogen-activated protein kinase 4 (MAPK4) in response to high temperature in lettuce (Lactuca sativa). Int. J. Agric. Biol., 21: 54-60.
Direct Link - Wei, S., X. Yang, G. Huo, G. Ge and H. Liu et al., 2020. Distinct metabolome changes during seed germination of lettuce (Lactuca sativa L.) in response to thermal stress as revealed by untargeted metabolomics analysis. Int. J. Mol. Sci., Vol. 21.
Direct Link - Catão, H.C.R., L.A.A. Gomes, R.M. Guimarães, P.H.F. Fonseca, F. Caixeta and A.G. Galvão, 2018. Physiological and biochemical changes in lettuce seeds during storage at different temperatures. Hortic. Bras., 36: 118-125.
CrossRefDirect Link - Yoong, F.Y., L.K. O’Brien, M.J. Truco, H. Huo and R. Sideman et al., 2016. Genetic variation for thermotolerance in lettuce seed germination is associated with temperature-sensitive regulation of ethylene response factor1 (ERF1). Plant Physiol., 170: 472-488.
CrossRefDirect Link - de Almeida, F.A., R. Silva-Mann, H.O. dos Santos, R.W. Pereira and A.F. Blank, 2019. Germination temperatures affect the physiological quality of seeds of lettuce cultivars. Biosci. J., 35: 1143-1152.
CrossRefDirect Link - Catão, H.C.R.M., L.A.A. Gomes, R.M. Guimarães, P.H.F. Fonseca, F. Caixeta and J.C. Marodin, 2016. Physiological and isozyme alterations in lettuce seeds under different conditions and storage periods. J. Seed Sci., 38: 305-313.
CrossRefDirect Link - Albuquerque, K.A.D., P.D.O. Alvim, P.D.A. Silva and A.D. Veiga, 2010. Physiological and biochemical quality of lettuce seeds coated with micronutrients, aminoacids and growth regulators. Biosci. J., 26: 843-848.
Direct Link - Ferreira, V.F., M.A. Ricaldoni, S.D.V.F. da Rosa, M.A. de Figueiredo, S.V.B. Coelho and T.B. Fantazzini, 2018. Endo-β-mannanase enzyme activity in the structures of Coffea arabica L. seeds under different types of processing and drying. Cienc. Rural, Vol. 48.
CrossRefDirect Link - da Silva, E.A.A., P.E. Tootop, A.C. van-Aelst and H.W.M. Hilhorst, 2004. Abscisic acid controls embryo growth potential and endosperm cap weakening during coffee (Coffea arabica cv. Rubi) seed germination. Planta, 220: 251-261.
CrossRef - Downie, B., H.W.M. Hilhorst and J.D. Bewley, 1994. A new assay for quantifying endo-β-d-mannanase activity using congo red dye. Phytochemistry, 36: 829-835.
CrossRefDirect Link - Hodges, D.M., J.M. DeLong, C.F. Forney and R.K. Prange, 1999. Improving the thiobarbituric acid-reactive-substances assay for estimating lipid peroxidation in plant tissues containing anthocyanin and other interfering compounds. Planta, 207: 604-611.
CrossRefDirect Link - Biemelt, S., U. Keetman and G. Albrecht, 1998. Re-aeration following hypoxia or anoxia leads to activation of the antioxidative defense system in roots of wheat seedlings. Plant Physiol., 116: 651-658.
CrossRefDirect Link - Giannopolitis, C.N. and S.K. Ries, 1977. Superoxide dismutases. I. Occurrence in higher plants. Plant Physiol., 59: 309-314.
CrossRefPubMedDirect Link - Havir, E.A. and N.A. McHale, 1987. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol., 84: 450-455.
CrossRefDirect Link - Azevedo, R.A., R.M. Alas, R.J. Smith and P.J. Lea, 1998. Response of antioxidant enzymes to transfer from elevated carbon dioxide to air and ozone fumigation, in the leaves and roots of wild-type and a catalase-deficient mutant of barley. Physiologia Plant., 104: 280-292.
CrossRefDirect Link - Nakano, Y. and K. Asada, 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in Spinach chloroplasts. Plant Cell Physiol., 22: 867-880.
CrossRefDirect Link - Mattana, E., M. Sacande, K.A. Sanogo, R. Lira, P. Gomez-Barreiro, M. Rogledi and T. Ulian, 2017. Thermal requirements for seed germination of underutilized Lippia species. South Afr. J. Bot., 109: 223-230.
CrossRefDirect Link - Amâncio, B.C.S., K.P. Govêa, L. de Oliveira Ribeiro Trindade, A.R. da Cunha Neto, T.C. de Souza and S. Barbosa, 2020. Sandwich method applied to the screening of allelopathic action in Byrsonima spp. (Malpighiaceae). Biologia, 75: 175-182.
CrossRefDirect Link - Nascimento, W.M., D.J. Cantliffe and D.J. Huber, 2001. Endo-β-mannanase activity and seed germination of thermosensitive and thermotolerant lettuce genotypes in response to seed priming. Seed Sci. Res., 11: 255-264.
Direct Link - Nascimento, W.M., D.J. Huber and D.J. Cantliffe, 2013. Carrot seed germination and ethylene production at high temperature in response to seed osmopriming. Hortic. Bras., 31: 554-558.
CrossRefDirect Link - Wang, A., J. Li and J.D. Bewley, 2004. Molecular cloning and characterization of an endo-β-mannanase gene expressed in the lettuce endosperm following radicle emergence. Seed Sci. Res., 14: 267-276.
CrossRefDirect Link - Nascimento, W.M., 2003. Preventing thermoinhibition in a thermosensitive lettuce genotype by seed imbibition at low temperature. Sci. Agricola, 60: 477-480.
CrossRefDirect Link - Geshnizjani, N., F. Ghaderi-Far, L.A.J. Willems, H.W.M. Hilhorst and W. Ligterink, 2018. Characterization of and genetic variation for tomato seed thermo-inhibition and thermo-dormancy. BMC Plant Biol., Vol. 18.
CrossRefDirect Link - de Freitas, M.N., S.D.V.F. da Rosa, A.D.C.S. Clemente and C.C. Pereira, 2017. Relevance of endo-β-mannanase enzyme in coffee seed deterioration process. Afr. J. Agric. Res., 12: 1253-1258.
CrossRefDirect Link - Bicalho, E.M., S.Y. Motoike, E.E. de Lima e Borges, G. da Mata Ataíde and V.M. Guimarães, 2016. Enzyme activity and reserve mobilization during macaw palm (Acrocomia aculeata) seed germination. Acta Bot. Brasilica, 30: 438-444.
CrossRefDirect Link - Assis, J.G.R., E.R. Marques, M.L.M. Carvalho, R.M.O. Pires, C.A. Lopes and R.W. Pereira, 2019. Expression of enzymes during the germination of seeds in endangered cerrado species. J. Agric. Sci., Vol. 11.
CrossRefDirect Link - Mazzottini-dos-Santos, H.C., L.M. Ribeiro and D.M.T. Oliveira, 2017. Roles of the haustorium and endosperm during the development of seedlings of Acrocomia aculeata (Arecaceae): Dynamics of reserve mobilization and accumulation. Protoplasma, 254: 1563-1578.
CrossRefDirect Link - Rocha, G.A., P.V. Romanatti, F.M. Oliveira, A.R.D. Neto, F.J. Pereira and M. Polo, 2018. Ecophysiology of the tree species Cedrela fissilis Vell. (Meliaceae) submitted to flooding. Cerne, 24: 323-333.
CrossRefDirect Link - Ventura, R.B., V.M. Soto, A.J. Otiniano and P.L. Gratão, 2019. Effect of water stress and temperature increase on productive, physiological and biochemical variables of Arnaldoa, 26: 287-296.
CrossRefDirect Link - Larré, C.F., C.L. Moraes, J. Borella, L. do Amarante, S. Deuner and J.A. Peters, 2016. Antioxidant activity and fermentative metabolism in the plant Erythrina crista-galli L. under flood conditions. Semina: Ciências Agrárias, 37: 567-580.
CrossRefDirect Link