
Guorong Wu
School of Energy and Power Engineering,
Ganesh K. Jaganathan
School of Energy and Power Engineering,
Danping Song
School of Energy and Power Engineering,
Baolin Liu
Institute of Cryobiology and Food Freezing, University of Shanghai for Science and Technology, 200093 Shanghai, China
Research Journal of Seed Science
Year: 2017 | Volume: 10 | Issue: 1 | Page No.: 38-42







ABSTRACT
Background: Many species with an impermeable seed coat, i.e., physical dormant (PY) have orthodox seeds that can be stored in liquid nitrogen without viablity loss. However, whether the dormancy should be broken before or after storage remains less clear. Objective: The importance of dormancy breaking time during cryopreservation of PY seeds was examined using three species Dodonaea viscosa (Sapindaceae), Senna auriculata (Fabaceae) and Delonix regia (Fabaceae). Materials and Methods: All three species have impermeable seed coats that was broken by dipping in boiling water for 60 sec to make them permeable. Seeds of all three species were divided into two groups with one group receiving dormancy breaking before storage in liquid nitrogen for 2 years and the other was stored dormant. Results: For D. viscosa, storing dormancy broken seeds in liquid nitrogen resulted in significantly less germination percentage after storage compared with the dormant seeds stored directly in liquid nitrogen and dormancy was broken at the end of storage (p<0.05). Similarly, S. ariculata seeds that received dormancy breaking treatment at the end of storage germinated to a higher percentage but the effect was not significant (p>0.05). On the other hand, there was no significant difference in germination percentage between dormant and non-dormant seeds of D. regia stored in liquid nitrogen (p>0.05), although storing dormant seeds and breaking dormancy at the end of storage resulted in higher germination percentage. Conclusion: The results showed that dormancy breaking time could have different effects determining the success of PY species cryopreservation.
PDF Abstract XML References Citation
Received: September 30, 2016;
Accepted: November 17, 2016;
Published: December 15, 2016
How to cite this article
Guorong Wu, Ganesh K. Jaganathan, Danping Song and Baolin Liu, 2017. Cryopreservation of Selected Physical Dormant Species with Special Focus on Dormancy Breaking Time. Research Journal of Seed Science, 10: 38-42.
URL: https://scialert.net/abstract/?doi=rjss.2017.38.42
URL: https://scialert.net/abstract/?doi=rjss.2017.38.42
INTRODUCTION
The understanding that seeds dried to lower moisture content often in the range of 3-7% f.w.b. can be successfully stored at -18°C or in liquid nitrogen for long-term without any viability loss has allowed the storage of seed accessions ex-situ and favored the conservation of plant germplasm1-3. However, this revolutionary method is practically applicable only for species that produce seeds which can be dried to low or safe moisture levels, i.e., orthodox seeds, because the group of so-called 'Recalcitrant seeds' lose their viability during drying and achieving long-term storage of those seeds at low temperature is not feasible due to the transformation of water into ice during low temperature4,5. Even for the orthodox seeds, attaining long-term viability is determined by moisture content of the seeds and the temperature at which the seeds are stored3. The vast majority of studies available agree that the lowest temperature contributes to longer life span of seeds but lowest moisture content is not necessarily a reliable predictor for ensured viability over time6. For example, seeds of Prunus avium L. can be desiccated to 1.6% without viability loss but seeds at such low moisture content are sensitive to liquid nitrogen storage7. This suggests that seeds stored in liquid nitrogen at very low moisture content could induce additional stress making survival problematical. Thus a safe moisture range exists for each species and storing seeds at this level is an important pre-requisite for enhanced longevity.
Beside moisture and temperature, it is becoming apparent that dormancy of orthodox seeds and the time at which dormancy-break is applied might also influence the success of seed storage. For example, Chmielarz7 showed that seeds of Prunus avium stored in liquid nitrogen for 2 years after applying dormancy break resulted in a significantly lower germination percentage compared with seeds given dormancy breaking at the end of storage. In a similar study, Chmielarz8 found that Franxinus excelsior seeds stored at -3°C and liquid nitrogen for 1 and 2 years, germination percentage was lower in seeds that had received dormancy breaking treatment before storage compared with dormancy breaking applied at the end of storage. On the other hand, Jaganathan et al.9 reported that Prunus armeniaca seeds were able to germinate to anequal percentage, regardless of dormancy breaking time, although breaking dormancy at the end of storage led to higher germination percentage. Because most the studies available on this subject matter have been conducted on Physiological Dormant (PD) seeds, one possible reason for the lack of consensus on this subject might be the induction of secondary dormancy in some species8. Interestingly, seeds with impermeable coats, i.e., physical dormancy (PY) are also stored for longtime in liquid nitrogen for conservation purpose10,11. Unlike PD species, seeds with PY cannot enter secondary dormancy during or after storage. However, in some species, the hard seed coats may crack during liquid nitrogen plunging11 but almost nothing about the importance of seed coat in preserving the viability of seeds during long-term storage. This study using three species having PY to further our knowledge of seed storage.
MATERIALS AND METHODS
Seeds of Dodonaea viscosa, Delonix regia and Senna auriculata were collected from 12, 6 and 17 individual plants, respectively growing in Coimbatore district of Tamil Nadu, India (11°10' N, 76°74' E). Seeds collected from the individual plants were grouped, cleaned and stored in jute bags at room temperature (25-30°C, 50-60% RH) until used in the experiments. The moisture content of the seeds was determined by oven drying method at 103°C for 17 h using four replicates of 20 seeds each12. Imbibition test was carried out on each species to test the proportion of impermeable seeds by determining the fresh weight of four replicates of 25 seeds of each species before placing them on wet paper towels taken in petri dishes and measuring the increase in seed weight at an hourly interval in a standard balance after drying the surface water using dry paper towels. Germination test was conducted using four replicates of 50 seeds for each species by sowing them in petri dishes containing 1% agar-water incubated at 20/25°C with 16/8 h-light/dark photoperiod. Germination counts were made twice a week for 6 weeks. A seed was scored as germinated when radical protruded through seed coat by 2 mm.
Freshly collected seeds of all three species were dipped in boiling water for 60 sec to make them permeable. To test the importance of dormancy breaking time in cryopreservation, batches of impermeable seeds dipped in boiling water and impermeable seeds (not given any treatments) were placed in cryoflex tubes in four replicates of 50 seeds each species per group. These tubes were plunged directly into Dewar flask and held for 26 months, except D. viscosa which was stored only for 22 months. Liquid nitrogen was replenished at regular intervals when the level dropped. At the end of the storage period, the seeds received dormancy break before storage was germinated. Whereas, impermeable seeds with no treatment that were not cracked during liquid nitrogen exposure were dipped in boiling water for 60 sec and germinated.
The control i.e., seeds dipped in hot-water and germinated without liquid nitrogen exposure, dormancy broken before liquid nitrogen storage and dormancy broken after liquid nitrogen storage germination percentage were analyzed for statistical significance by a one-way ANOVA. It was used arcsine transformed data to improve the normality but original percentage values are presented. All the statistical analysis were conducted in SPSS (version 20.0).
RESULTS AND DISCUSSION
The moisture content of the seeds tested immediately after collection varied between species (Table 1), however, the range identified here indicate that all three species are orthodox. All the species can be dried further to lower moisture levels above silica gel without viability loss. The number of seeds germinating in each species differed but none of the species germinated above 20% (Table 1). In addition, the imbibition test showed the seeds that remained ungerminated were impermeable to water. When dipped in boiling water for 60 sec, germination percentage significantly improved in all the three species (p>0.05, Table 2). Collectively, these results suggest that all the species have PY.
Successful storage of PY seeds having orthodox behavior in liquid nitrogen has been reported for many species10,13-15. However, long-term storage results are rarely reported and particularly whether the dormancy breaking of these seeds should be employed before or after storage is not clear. To the best of our knowledge, this is the first study on PY species that attempted storage in liquid nitrogen for 2 years. The germination results of the three species stored for 22 or 24 months are given in Table 2. Results indicated that long-term storage of PY seeds in liquid nitrogen is feasible but the percentage of seed viability upon thawing is species-specific. Further, the final germination percentage also varied depending upon the dormancy broken time (Table 2). For Dodonaea viscosa and Senna auriculata seeds, breaking dormancy before storage led to lower germination percentage after storage compared with the groups received dormancy breaking at the end of storage and this effect was significant in Dodonaea viscosa (Table 2). However, the opposite was found to be true in Delonix regia seeds (p>0.05, Table 2).
A hard and impermeable seed coat protect the seed not only from mechanical stress but also from microorganism attack and from temperature and humidity fluctuations16-18.
| Table 1: | Details of species, year collected, initial moisture content and germination percentage at the time of collection |
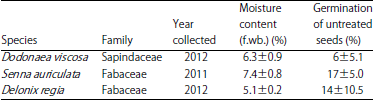 | |
| Table 2: | Germination (%) of three species before storage and after storage with dormancy broken at different time |
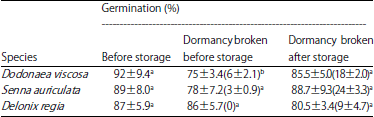 | |
Values given in parenthesis indicate the percentage of seeds with coat cracked during liquid nitrogen plunging | |
The importance of the seed coat in seed longevity has long been noted19,20. One of the important change that occur in hard seeds during liquid nitrogen immersion is the development of cracks in the seed coat. In some species including Trifolium subterraneum21, Medicago rugosa and M. ciliaris22 seeds developed deep cracks in the seed coat, which promoted germination as these cracks serve as openings for water entry but plunging seeds in liquid nitrogen only developed small cracks in M. orbicularis23. In Linum usitatissimum germination percentage was reduced to 34% after storing seeds in liquid nitrogen for 7 days compared with 41% before storage, because of the seed coat and other tissue damage11. However, seed-cracking has not been a common occurrence in other species. In Astragalus hamosus23, for example, no seeds had cracked coat during liquid nitrogen exposure. The size and shape of the seeds have been proposed as the key factors responsible for no seed coat crack in A. hamosus23. Nevertheless, in the present study, it was noted that a proportion of seeds in Dodonaea viscosa and Delonix regia had cracked seed coats when dormant seeds were immersed in liquid nitrogen. This suggested that seed size and shape may not be a reason for preserving seed coat structure during liquid nitrogen plunging.
From the three species investigated in the present study, it suggested that dormancy breaking time could have different effects on species cryopreservation. Overall, dormancy breaking at the end of storage is beneficial. However, it was noted that for some dry seeds the coats may crack and there is an increased possibility of viability loss during long-term storage. It can only speculate on the reasons why different species show different ability to maintain seed coat integrity during liquid nitrogen exposure. The seed coat thickness, the moisture content of the seeds, even the size and shape of the seeds may preserve the whole seed structure when exposed to low temperature, especially at a faster rate. However, in our preliminary experiments, it was observed that dry seeds had completely cracked seed coat during liquid nitrogen immersion, but very few seeds that had received dormancy breaking prior to liquid nitrogen immersion showed visible cracks developed. This behavior appears to be common in species with an impermeable coat24.
It has been suggested that the moisture content of PY seeds can affect the level of dormancy25. Thus, it is possible to surmise that the difference in seed collection year might contribute to the biased result in seed storage. However, the difference in levels of PY and the proportion of seeds with impermeability is closely linked to the moisture content resulting from maturation site. Therefore, we believe that seeds of Senna ariculata collected in 2011 did not affect our results and comparison with other two species collected in 2012. Because moisture content of the seed is an important component, the dryness of seeds, which varied between seed-to-seed, could possibly be the reason for some seeds with cracks. This suggests that optimal moisture not only to avoid freezing injury but also to prevent the seed coat integrity, particularly in PY species, may exist. For successful long-term storage of PY species, this threshold level must be identified.
CONCLUSION
It concluded that all the three species investigated in this study namely Senna auriculata, Dodonaea viscosa and Delonix regia produce orthodox seeds that are amendable to cryopreservation. However, the dormancy broken time critically influences the total No. of seeds germination. Overall, those results and the previous ones suggest that applying dormancy breaking at the end of storage can be beneficial but it call for preliminary experiments when cryopreservation attempts on different species are performed on a large scale.
ACKNOWLEDGMENTS
We kindly thank the help of researchers and students who helped with seed collection and some storage experiments.
REFERENCES
- Engelmann, F., 2004. Plant cryopreservation: Progress and prospects. In Vitro Cell. Dev. Biol. - Plant, 40: 427-433.
CrossRefDirect Link - Hay, F.R. and R.J. Probert, 2013. Advances in seed conservation of wild plant species: A review of recent research. Conservation Physiol., 1: 1-11.
CrossRefDirect Link - Li, D.Z. and H.W. Pritchard, 2009. The science and economics of ex situ plant conservation. Trends Plant Sci., 14: 614-621.
CrossRefDirect Link - Roberts, E.H., 1973. Predicting the storage life of seeds. Seed Sci. Technol., 1: 499-514.
Direct Link - Walters, C., L.M. Wheeler and P.C. Stanwood, 2004. Longevity of cryogenically stored seeds. Cryobiology, 48: 229-244.
CrossRefDirect Link - Ellis, R.H. and T.D. Hong, 2006. Temperature sensitivity of the low-moisture-content limit to negative seed longevity-moisture content relationships in hermetic storage. Ann. Bot., 97: 785-791.
CrossRefDirect Link - Chmielarz, P., 2009. Cryopreservation of dormant orthodox seeds of forest trees: Mazzard cherry (Prunus avium L.). Ann. Forest Sci., 66: 1-9.
CrossRefDirect Link - Chmielarz, P., 2009. Cryopreservation of dormant European ash (Fraxinus excelsior) orthodox seeds. Tree Physiol., 29: 1279-1285.
CrossRefDirect Link - Jaganathan, G.K., G.R. Wu, X.Y. Song and B.L. Liu, 2015. Cryopreservation of dormant Prunus armeniaca L. seeds. Seed Sci. Technol., 43: 456-466.
CrossRefDirect Link - Salomao, A.N., 2002. Tropical seed specie's responses to liquid nitrogen exposure. Braz. J. Plant Physiol., 14: 133-138.
CrossRefDirect Link - Stanwood, P.C., 1980. Tolerance of crop seeds to cooling and storage in liquid nitrogen (-196°C). J. Seed Technol., 5: 26-31.
Direct Link - Kholina, A.B. and N.M. Voronkova, 2012. Seed cryopreservation of some medicinal legumes. J. Bot.
CrossRefDirect Link - Perez-Garcia, F. and E.M. Gonzalez-Benito, 2008. Seed cryopreservation of Halimium and Helianthemum species. Cryoletters, 29: 271-276.
Direct Link - Touchell, D.H. and K.W. Dixon, 1993. Cryopreservation of seed of Western Australian native species. Biodiversity Conservation, 2: 594-602.
CrossRefDirect Link - Dalling, J.W., A.S. Davis, B.J. Schutte and A.E. Arnold, 2011. Seed survival in soil: Interacting effects of predation, dormancy and the soil microbial community. J. Ecol., 99: 89-95.
CrossRefDirect Link - Mohamed-Yasseen, Y., S.A. Barringer, W.E. Splittstoesser and S. Costanza, 1994. The role of seed coats in seed viability. Bot. Rev., 60: 426-439.
CrossRefDirect Link - Baskin, J.M. and C.C. Baskin, 2000. Evolutionary considerations of claims for physical dormancy-break by microbial action and abrasion by soil particles. Seed Sci. Res., 10: 409-413.
CrossRefDirect Link - Crocker, W. and W.E. Davis, 1914. Delayed germination in seed of Alisma plantago. Bot. Gazette, 58: 285-321.
Direct Link - Pritchard, H.W., K.R. Manger and F.G. Prendergast, 1988. Changes in Trifolium arvense seed quality following alternating temperature treatment using liquid nitrogen. Ann. Bot., 62: 1-11.
Direct Link - Patane, C. and K.J. Bradford, 1993. Seed dormancy in two Medicago species typical of natural pastures of Ragusa plateau. J. Agron., 4: 412-418.
Direct Link - Patane, C. and F. Gresta, 2006. Germination of Astragalus hamosus and Medicago orbicularis as affected by seed-coat dormancy breaking techniques. J. Arid Environ., 67: 165-173.
CrossRefDirect Link - Wiesner, L.E., J.E. Lautmann, P.C. Stanwood and L.J. Wheeler, 1994. The effect of liquid nitrogen on alfalfa seed viability, emergence and broken cotyledons. J. Seed Technol., 18: 1-6.
Direct Link - Jaganathan, G.K., 2016. Influence of maternal environment in developing different levels of physical dormancy and its ecological significance. Plant Ecol., 217: 71-79.
CrossRefDirect Link