
Nara Edreira Alencar
Laboratory of Plant Viruses, Department of Phytopathology, University of Lavras, P.O. Box 3037, Lavras-Minas Gerais, Brazil
Antonia dos Reis Figueira
Laboratory of Plant Viruses, Department of Phytopathology, University of Lavras, P.O. Box 3037, Lavras-Minas Gerais, Brazil
Heloisa Oliveira dos Santos
Department of Agriculture, University of Lavras, P.O. Box 3037, Lavras-Minas Gerais, Brazil
Priscilla Sousa Geraldino Duarte
Laboratory of Plant Viruses, Department of Phytopathology, University of Lavras, P.O. Box 3037, Lavras-Minas Gerais, Brazil
Research Journal of Seed Science
Year: 2016 | Volume: 9 | Issue: 1 | Page No.: 14-21







ABSTRACT
Squash mosaic virus (SqMV) is transmitted by seeds and infect various species of Cucurbitaceae causing severe yield losses. Avoiding the early introduction of the SqMV via seeds in field crops is the most important control measure; therefore, finding a reliable method which can identify the physiological qualities of the seeds produced by SqMV infected plants, could help select uninfected seed lots. The aim of this study was to use an X-ray test to evaluate the internal damage of seeds produced by infected Cucurbita pepo cv. caserta, followed by the investigation of germination, the percentage of abnormal plants, emergence, emergence speed, tetrazolium and the emergence speed index. Overall, the percentage of full seeds was much lower in the infected seeds, while the number of damaged seeds was higher. In the germination test, with regard to the percentage of abnormal plants and emergence, the infected seeds showed significant decreases in the performance, when compared with the healthy seeds. The mean speed of emergence and seed vigor were also lower in the infected seeds. Moreover, the radiographic analysis and vigor tests proved, for the first time, to be useful tools in assessing the quality of the seeds produced by the SqMV infected plants and revealed that the quality of the infected seeds was always lower than that of the healthy seeds. Thus, the use of these techniques can assist in the design and optimization of seed analysis, or contribute to additional studies involving virally infected plants, such as in pumpkin seeds.
PDF Abstract XML References Citation
Received: November 24, 2015;
Accepted: December 04, 2015;
Published: December 15, 2015
How to cite this article
Nara Edreira Alencar, Antonia dos Reis Figueira, Heloisa Oliveira dos Santos and Priscilla Sousa Geraldino Duarte, 2016. Image Analysis and Physiological Quality Assessment of Seeds Produced in Pumpkin Plants Infected with the Squash Mosaic Virus (SqMV). Research Journal of Seed Science, 9: 14-21.
URL: https://scialert.net/abstract/?doi=rjss.2016.14.21
URL: https://scialert.net/abstract/?doi=rjss.2016.14.21
INTRODUCTION
The cucurbit, with about 118 genera and over 800 species is one of the most important families of plants cultivated worldwide. Among the species of greatest economic importance are the pumpkin (Cucurbita moschata L.), zucchini (Cucurbita pepo L.), melon (Cucumis melo L.), watermelon (Citrullus vulgaris L.) and cucumber (Cucumis sativus L.) (Bisognin, 2002).
The use of high quality seeds is one critical factor in the commercial production of pumpkin fruits. Overall, the worldwide production of pumpkins in 2014 was 23.4 million tons, grown in an area of 1.83 million hectares and providing an average yield of 13.4 t ha–1 (FAO., 2015). Low yields are generally associated with the use of seeds with poor germination rates, reduced vigor and high contamination by pathogens, such as, viruses. Several surveys on the viral prevalence in Brazilian cucurbita crops have shown high incidences of many viruses, such as the Zucchini Yellow Mosaic Virus (ZYMV), Papaya Ringspot Virus W strain (PRSV-W) and Cucumber Mosaic Virus (CMV), but low or no incidence of the Squash mosaic virus (SqMV) (Moura et al., 2001; Halfeld-Vieira et al., 2004; Ramos et al., 2003; Da Silveira et al., 2009). However, Alencar et al. (2012) in a survey of viruses that infect cucurbits in the Tocantins state of Brazil, found that 56% of the samples were infected with SqMV.
The first report on mosaic symptoms in pumpkins caused by the SqMV was published in 1934, in California (USA). Since then, this virus has also been reported in North and South America, Europe and Australia (Sherf and MacNab, 1986). The SqMV is a member of the Comovirus genus, belonging to the subfamily Comovirinae, family Secoviridae and order Picornavirales (Sanfacon et al., 2011). Its genome is composed of two ssRNAs+, encapsulated separately in isometric particles of 20-30 nm in diameter. The RNA1 encodes one polyprotein, which is later processed into 5 proteins: protease co-factor, helicase, VPg (viral protein genome-linked), protease and RNA-dependent RNA polymerase. The RNA2 encodes 3 proteins: MP (Movement Protein), LCP (Large Capsid Protein) and SCP (Small Capsid Protein) (Haudenshield and Palukaitis, 1998; Han et al., 2002; Li et al., 2015).
The natural host plants of the SqMV are limited to the Cucurbitaceae family (Haudenshield and Palukaitis, 1998; Nelson and Knuhtsen, 1973a) and the symptoms depend on the virus isolate, plant species and environment; however, most of the susceptible hosts show a severe systemic mosaic, with leaf and fruit deformation. The SqMV can be transmitted by chrysomelid beetles (Diabrotica spp. and Acalyma spp.) (Freitag, 1956; Lastra, 1968; Sitterly, 1960; Stoner, 1963), coccinellid beetles (Cohen and Nitzany, 1963), grasshoppers (Stoner, 1963) and by seeds (Blancard et al., 1996).
About 18% of the known viruses can be transmitted by one or more natural hosts, while 33% can be transmitted by at least one host (Stace-Smith and Hamilton, 1988; Johansen et al., 1994). The rate of virus transmission, from infected plants to seeds, depends on several factors, like the virus and plant species (Albrechtsen, 2006) and can vary from values below 1% up to 100% (Alvarez and Campbell, 1978; Nelson and Knuhtsen, 1973a, b). Viral transmission via the seeds allows the early introduction of the pathogen inoculum in the field, leading to high spreading in the presence of a beetle vector. Therefore, using virus-free seeds is one of the most important measures to control and prevent the spread of this virus. The SqMV can be transmitted by approximately 10% of the seeds produced by infected plants, which means that the seeds should be carefully analyzed to detect the virus, aiming to avoid early virus spreading and consequently, undesirable yield losses (Blancard et al., 1996).
The occurrence of empty and morphologically damaged seeds is quite common; however, the detection of morphologically damaged seeds is difficult, due to the presence of a hard seed coat, as well as the methodology used for seed processing, which does not allow viewing the embryo without damaging the seeds. A good solution for this problem is an analysis of the seeds via X-rays. The X-ray test is not a test of individual viability, but it generates information that helps to assess the viability of the seed lot, showing any morphological deficiencies that indicate their potential. This test has been widely used to assess the internal structures of forest and vegetable seeds, since it is fast, accurate and non-destructive, allowing the determination of the maturity stage and the occurrence of empty or undeveloped seeds, as well as any mechanical damage or injuries caused by insects (Pinto et al., 2009; Gagliardi and Marcos-Filho, 2011; Gomes-Junior et al., 2012; Kotwaliwale et al., 2014; Panchal et al., 2014). Moreover, the seeds submitted for X-ray analysis can be subsequently germinated, establishing a correlation between the mechanical damage or alterations observed in the inner parts of the seeds and any effects observed in the seed germination process (Dos Santos et al., 2009; Gomes-Junior et al., 2012; Gagliardi and Marcos-Filho, 2011; Panchal et al., 2014).
Although, X-ray analyses are widely used to assess the viability of seeds (Bino et al.,1993; Liu et al., 1993; Gagliardi and Marcos-Filho, 2011; Costa et al., 2014), they have not been employed to analyze seeds produced by plants infected with viruses, with regard to detecting physical damage, the capability for germination and the production of viable plants. Therefore, in this study, the seeds produced by pumpkin plants (cv. caserta) infected with the SqMV were analyzed via X-ray in order to evaluate the potential of this technique for indicating internal damage in infected seeds. The analyzed seeds were subsequently sown and the germination rate of the obtained plants was evaluated and compared with the results of the X-ray analysis, in order to establish a correlation between these results and the seed viability in producing healthy plants.
MATERIALS AND METHODS
This study was conducted at the Laboratory of Seed Analysis, Department of Agriculture and at the Laboratory of Molecular Virology of the Department of Plant Pathology (DFP) of the Federal University of Lavras, using pumpkin seeds produced by SqMV infected and healthy plants.
The SqMV isolate used in the experiments belongs to the DFP/UFLA collection and is named SqMV-LC2. It was originally collected from a naturally infected watermelon plant growing in the "Lagoa da Confusão" municipality of the state of Tocantins in Brazil. The infected leaves of the pumpkin plant were kept dehydrated in a -80°C freezer and the virus was mechanically inoculated in pumpkins (cv. caserta) for the virus multiplication.
The seed production by the infected and healthy pumpkin plants was conducted in a greenhouse, under controlled conditions. After flowering, artificial pollination was done by detaching the male flowers from the plants and transferring the pollen to the stigma of the female flowers, daily, between 8 and 10 AM. When the pollen was viable in the open flowers. The fruits produced were allowed to ripen and the seeds were collected, processed and kept in a cold room for use during the experiments.
The healthy and infected seeds were submitted to X-ray analysis, using a Faxitron X-ray unit (MX20 model DC-12 option) with an intensity of 18 kV for 20 sec During the experiment, 3000 seeds each of the healthy and infected plants were analyzed, they were distributed in parcels containing one hundred seeds, arranged in transparent blades and five repetitions were done. The experimental design was completely randomized in a factorial arrangement (3×2) with three seed classes and two lots of seeds (healthy and infected with SqMV).
The obtained data, previously submitted for normality tests of waste and variance homoscedasticity were tested using an analysis of variance and the significant differences between the means were determined by using the Tukey test at 5% probability. The statistical analyses were performed using the SISVAR® program (Ferreira, 2011).
The moisture content was determined in a 105°C oven, for 24 h (Ministerio da Agricultura and Pecuaria e Abastecimento, 2009) using two replicates of 50 seeds from each batch/class. After being taken to the desiccators for cooling, the samples were weighed and the results were expressed as a percentage.
For each germination test, calculations of the T50 and emergence speed index were conducted with 20 seeds per class and 10 repetitions. Sowing was done between Germitest type papers moistened with distilled water (2.5 mL g–1 of paper), with the seeds arranged according to the images obtained in the X-ray analysis. The scrolls were kept in a growth chamber at 25°C and the evaluations were performed in two counts: On the fourth and the eighth days after sowing, according to the rules for seed analysis (ISTA., 2009). The results were expressed as the mean percentage of the normal and abnormal plantlets. The non-germinated seeds from each treatment were cut lengthwise, using a scalpel, placed in a dark plastic container and immersed in a tetrazolium salt solution of 0.075% for 3.5 h at 37°C. After staining, an evaluation was performed using a common stereoscopic microscope to determine the viability (Gagliardi and Marcos-Filho, 2011).
In the emergence test, the seeds were arranged in a sequence of X-ray images and were sown in plastic trays containing the proper substrate. Fifty seeds were tested, with 4 random replicates. Visual inspections were performed daily, from the first day after emergence and the percentage of normal seedlings was evaluated at 21 days. The daily counting of the seed emergence was carried out at the same time each day, until emergence stabilization, to calculate the emergence percentage and emergence speed index, according to Edmond and Drapala (1958):
Where:
| VE | = | Emergency speed (in days) |
| E1, E2, En | = | Normal number of emerged plants in the first, second and last counts |
| N1, N2, Nn | = | Number of days from the first, second and last counts on the days after sowing |
After germination, the seedlings were tested using the double-antibody sandwich enzyme linked immunosorbent assay (DAS-ELISA) (Clark and Adams, 1977) using polyclonal antisera against the SqMV (from Bioreba) following the manufacturer’s protocol and via the immunocapture reverse transcription polymerase chain reaction (IC-RT-PCR), according to Shankar et al. (2009), using the same antisera and the following primers: SqMV-1648 F: 5’ TTTGACGGCATGGTC 3 and SqMV-2465 R: 5’ GGAAAGAAGCCACAAC 3’.
RESULTS AND DISCUSSION
Based on the results of the X-ray test, the seeds were divided into three classes: filled to apparently normal (Fig. 1a), physically damaged (for seeds that showed any alteration, such as those seen in Fig. 1b and c and empty, with no apparent seed content (Fig. 1d), according to the ISTA (2009).
The average water content of the seeds was 8.3%, with a maximum variation of 0.5%. This low variation in the water content at this point is important, because when the variation is high, the process of deterioration is accelerated and the formation of products that cause immediate harm is likely to occur. Since pumpkin seeds are orthodox, they not only tolerate desiccation, but likely rely on it to redirect the metabolic processes of development towards germination (Golovina et al., 2000; Hoekstra et al., 2001; Pritchard et al., 2004; Walters, 2015). In addition, is known that in some seeds, the lower the water content, the higher the optical density, enabling greater differentiation of the seed structures in radiographic analyses (Simak et al., 1989). For smaller seeds, such as in bell peppers, the water content necessary to generate a good image can be a little higher.
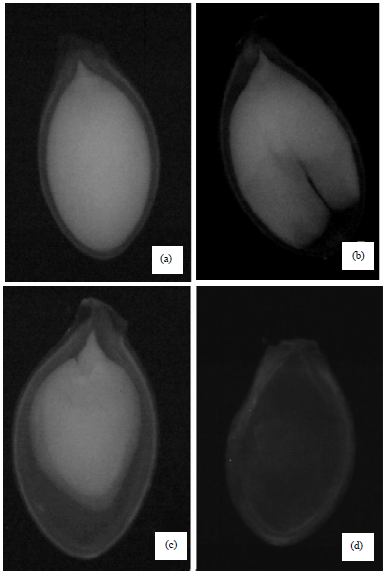 | |
Fig. 1(a-d): | Radiographic images of pumpkins seeds. (a) Filled seed, (b) Physically damaged seed 1, (c) Physically damaged seed 2 and (d) Empty seed |
Gagliardi and Marcos-Filho (2011) tested pepper seeds with 10, 12, 15, 18 and 20% water content and found that the least amount of water necessary for good radiography was 12%.
The average percentages of the seeds in each class are described in Table 1. The analysis of variance indicated significant differences between the percentages of seeds in the same class, produced by the healthy and infected plants. The infected plants produced only 15% filled seeds, which means that most of them were damaged or empty. When compared with the healthy plants, which produced more than 90% filled seeds, it was possible to observe how much the SqMV can negatively affect the seed production. De Carvalho et al. (2009) analyzed l0 lots of healthy Cucurbita pepo seeds and found from 54-89% filled seeds, showing that the percentage of filled seeds produced by the infected plants was really lower than that expected for the pumpkin seeds of any species.
The results of the germination rate, percentage of abnormal seedlings, emergence, average speed of emergence, viability (tetrazolium test), T50 and emergency speed index of each seed class are described in Table 2.
The performance of the seeds produced by the infected plants was expressly lower when compared with the ones produced by the healthy plants. In addition, those seeds classified as filled, produced by the SqMV infected plants, showed less than 50% germination, while almost 100% of the filled seeds produced by the healthy plants were able to germinate. The percentage of germination of the damaged seeds was also lower, when compared with the seeds from the healthy plants. The potential for X-ray tests to analyze seed quality has been reported by several authors and usually, the percentage of germination of the filled seeds is much higher than that of the damaged seeds (De Carvalho et al., 2009; Dos Santos et al., 2009; Gagliardi and Marcos-Filho, 2011; Sturiao et al., 2012; Silva et al., 2014). In this study, besides producing a lower number of filled seeds, the infected plants also produced seeds with a lesser germination potential.
The same behavior was observed when the other parameters were analyzed. For instance, the seeds produced by the healthy plants generated only 12% abnormal seedlings in the damaged class; while 34% of the filled seeds and 49% of the damaged seeds from the infected plants generated abnormal seedlings. It is known that abnormal seedlings may result from genetic factors, as well as from the environment and management practices, leading to the absence, atroph, or deformity of their vital organs, such as the radicle, hypocotyl and plumule, which are difficult or impossible to overcome (Rajjou et al., 2012).
| Table 1: | Mean percentage of seeds in each of the three classes detected in X-ray analysis |
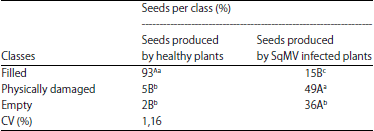 | |
*Means followed by the same letters, uppercase in rows and lowercase columns, are not significantly different from each other (Tukey’s test, p<0.05) | |
| Table 2: | Results of the evaluation of several parameters presented by seeds produced by healthy and SqMV infected plants |
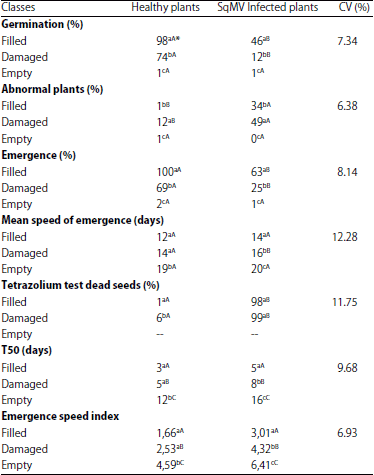 | |
*Means followed by the same letters, uppercase in rows and lowercase in columns, are not significantly different from each other (Tukey’s test, p<0.05) | |
The largest generation of abnormal seedlings by the seeds from the infected plants seems to be due to the viral effect on the seed tissues, leading to several alterations in their normal physiological conditions.
A small number of empty seeds was able to germinate and generate seedlings. This production of seedlings by empty seeds can be explained by the fact that even though they seemed empty, the difference in the density captured by the X-ray test was not large enough to show the presence of the cotyledon and embryo. Moreover, in the emergence tests, the results were similar to those observed in the germination tests, in which the seed performance was superior in the filled seeds and inferior in the damaged and empty seeds. In the seeds produced by the infected plants, the percentage of emergence was lower in the filled and damaged classes and similar in the empty class. In addition, the same statistical similarity was seen in the speed of emergence of the filled and empty classes; however, it was still lower in the damaged class. These results could be related to the normal physiological variations among the different seed lots tested in the experiments.
Variations between the percentage of germination and speed of emergence in distinct genotypes have frequently been detected. For example, Dos Santos et al. (2015) analyzed Passiflora edulis seeds and noted differences in the germination and emergence speed between the genotypes and the progeny. They concluded that the seed dormancy was associated with the genotype of the embryo and that it was most likely caused by a dominant genetic effect. However, in this study, which used pumpkin seeds from a single genotype, it was possible to determine the variation in the speed of emergence of the two damaged lots, demonstrating once again the effect of the viral infection on the seed performance.
The results obtained with the germination tests were not always similar to those of the emergence, which means that germination tests are not sufficient for indicating the potential performance of a seed lot. During the production process, information on the physiological potential of the seeds is of fundamental importance in making decisions. Along with the germination test, the most often used parameter for determining the seed quality should be a vigor test, which describes the percentage and speed of emergence. These combined data better reflect the behavior and degree of deterioration of the seeds in stress conditions; however, the vigor test usually has a better correlation with the seedling emergence (Rajjou et al., 2012).
The tetrazolium test showed that practically all of the non-germinating seeds from the infected plants were dead, but it did not correlate with the seeds produced by the healthy plants. This shows that the SqMV not only affected the performance of the seeds, but it also led to seed death. The tetrazolium test is considered to be promising, with the main advantage of enabling the evaluation of the viability of the seeds in a relatively short period of time. It can be used to establish bases for marketing, determining the point of harvest and evaluating seed viability during the processing and storage periods (De Carvalho et al., 2014).
Finally, the T50 and emergence speed index were not relevantly affected by the SqMV and presented similar values for both the healthy and infected plants. However, they had high variations among the seed classes, ranging from 5 days for the filled seeds to 16 days for the empty seeds.
In this study, the radiographic analyses showed very good potential for classifying and indicating the viability of the seeds produced by the SqMV infected pumpkin seeds, based on internal morphology. Similar results have been obtained by several authors, including De Carvalho et al. (2009) using Cucurbita moschata cv. Piramoita seeds, Gagliardi and Marcos-Filho (2011) using bell pepper seeds, Sturiao et al. (2012) using palma seed and De Carvalho et al. (2010) using castor bean seeds.
Considering that the SqMV has a negative effect on infected pumpkin plants, decreasing the physiological quality of the seeds produced, this test may help evaluate the quality of the seed lots destined for planting and multiplication. In addition, it could contribute to decreasing the introduction and spreading of the SqMV through seeds in pumpkin production fields.
The DAS-ELISA and the IC-RT-PCR, when applied to the sprouted seeds (including the tegument), detected the SqMV in nearly all of the samples. Both of these tests were very efficient in this detection, however, the IC-RT-PCR was slightly more sensitive, detecting 100% of the virus when compared to 96% of the samples detected by DAS-ELISA. It is known that not all of the plants derived from these seeds become infected; for example, De Almeida (2013) observed that, despite the fact that all of the seeds coming from the infected plants tested positive for SqMV, only 15% produced infected seedlings.
ACKNOWLEDGMENT
We thank the CNPq, CAPES and FAPEMIG funding agencies for providing research grants to the research team.
REFERENCES
- Alencar, N.E., A.D.R. Figueira, J.E.M. de Almeida, L.B. dos Santos and I.R. do Nascimento, 2012. Molecular biological identification of detected viruses in cucurbit species from the State of Tocantins. J. Biotechnol. Biodivers., 3: 32-37.
Direct Link - Alvarez, M. and R.N. Campbell, 1978. Transmission and distribution of squash mosaic virus in seeds of cantaloupe. Phytopathology, 68: 257-263.
CrossRefDirect Link - Bisognin, D.A., 2002. Origin and evolution of cultivated cucurbits. Ciencia Rural, 32: 715-723.
CrossRefDirect Link - Bino, R.J., J.W. Aartse and W.J. van der Burg, 1993. Non-destructive X-ray analysis of Arabidopsis embryo mutants. Seed Sci. Res., 3: 167-170.
CrossRefDirect Link - De Carvalho, M.L.M., R.A. Alves and L.M. de Oliveira, 2010. Radiographic analysis in castor bean seeds (Ricinus communis L.). Revista Brasileira de Sementes, 32: 170-175.
CrossRefDirect Link - De Carvalho, M.L.M., C.D. da Silva, L.M. de Oliveira, D.G. Silva and C.M. Caldeira, 2009. X-Ray test to evaluate pumpkin seed quality. Revista Brasileira de Sementes, 31: 221-227.
CrossRefDirect Link - De Carvalho, T.C., C.R.S. Grzybowski, O.C. Ohlson and M. Panobianco, 2014. Adaptation of the tetrazolium test method for estimating the viability of sorghum seeds. J. Seed Sci., 36: 246-252.
CrossRefDirect Link - Clark, M.F. and A.N. Adams, 1977. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol., 34: 475-483.
CrossRefPubMedDirect Link - Costa, D.S., J. Kodde and S.P.C. Groot, 2014. Chlorophyll fluorescence and X-ray analyses to characterise and improve paddy rice seed quality. Seed Sci. Technol., 42: 449-453.
CrossRefDirect Link - Cohen, S. and F.E. Nitzany, 1963. Identity of viruses affecting cucurbits in Israel. Phytopathology, 53: 193-196.
Direct Link - Ferreira, D.F., 2011. [Sisvar: A computer statistical analysis system]. Ciencia Agrotecnologia, 35: 1039-1042, (In Portuguese).
Direct Link - Freitag, J.H., 1956. Beetle transmission, host range and properties of squash mosaic virus. Phytopathology, 46: 73-81.
Direct Link - Gagliardi, B. and J. Marcos-Filho, 2011. Relationship between germination and bell pepper seed structure assessed by the X-Ray test. Scientia Agricola, 68: 411-416.
CrossRefDirect Link - Golovina, E.A., F.A. Hoekstra and A.C. van Aelst, 2000. Programmed cell death or desiccation tolerance: Two possible routes for wheat endosperm cells. Seed Sci. Res., 10: 365-380.
CrossRefDirect Link - Gomes-Junior, F.G., J.T. Yagushi, U.L. Belini, S.M. Cicero and M. Tomazello-Filho, 2012. X-ray densitometry to assess internal seed morphology and quality. Seed Sci. Technol., 40: 102-107.
CrossRefDirect Link - Halfeld-Vieira, B.A., N.F. Ramos, F.A.R. Filho, M.F.B. Goncalves, K.L. Nechet, P.R. Pereira and J. Lima, 2004. Serological identification of virus species in watermelon in the State of Roraima. Fitopatologia Brasileira, 29: 687-689.
CrossRefDirect Link - Han, S.S., K. Yoshida, A.V. Karasev and T. Iwanami, 2002. Nucleotide sequence of a Japanese isolate of Squash mosaic virus. Arch. Virol., 147: 437-443.
CrossRefDirect Link - Haudenshield, J.S. and P. Palukaitis, 1998. Diversity among isolates of Squash mosaic virus. J. Gen Virol., 79: 2331-2341.
CrossRefDirect Link - Hoekstra, F.A., E.A. Golovina and J. Buitink, 2001. Mechanisms of plant desiccation tolerance. Trends Plant Sci., 6: 431-438.
CrossRefPubMedDirect Link - Johansen, E., M.C. Edwards and R.O. Hampton, 1994. Seed transmission of viruses: Current perspectives. Annu. Rev. Phytopathol., 32: 363-386.
CrossRefDirect Link - Kotwaliwale, N., K. Singh, A. Kalne, S.N. Jha, N. Seth and A. Kar, 2014. X-ray imaging methods for internal quality evaluation of agricultural produce. J. Food Sci., 51: 1-15.
CrossRefDirect Link - Li, R., S. Gao, S. Berendsen, Z. Fei and K.S. Ling, 2015. Complete genome sequence of a novel genotype of squash mosaic virus infecting squash in Spain. Genome Announcements, Vol. 3.
CrossRefDirect Link - Liu, Y., W.J. van der Burg, J.W. Aartse, R.A. van Zwol, H. Jalink and R.J. Bino, 1993. X-ray studies on changes in embryo and endosperm morphology during priming and imbibition of tomato seeds. Seed Sci. Res., 3: 171-178.
CrossRefDirect Link - Moura, M.C.C.L., J.A.A. Lima, V.B. Oliveira and M.F.B. Goncalves, 2001. [Serological identification of virus species infecting cucurbits in producing areas of the State of Maranhao, Brazil]. Fitopatologia Brasileira, 26: 90-92.
Direct Link - Ramos, N.F., J.A.A. Lima and M.F.B. Goncalves, 2003. Symptoms of potyvirus interactions on melon hybrids, watermelon varieties and zucchini. Fitopatologia Brasileira, 28: 199-204.
CrossRefDirect Link - Nelson, M.R. and H.K. Knuhtsen, 1973. Squash mosaic virus variability: Review and serological comparisons of six biotypes. Phytopathology, 63: 920-926.
Direct Link - Nelson, M.R. and H.K. Knuhtsen, 1973. Squash mosaic virus variability: Epidemiological consequences of differences in seed transmission frequency between strains. Phytopathology, 63: 918-920.
Direct Link - Panchal, K.P., N.R. Pandya, S. Albert and J.J. Gandhi, 2014. A X-ray image analysis for assessment of forage seed quality. Int. J. Plant Anim. Environ. Sci., 4: 103-109.
Direct Link - Pinto, T.L.F., S.M. Cicero, J.B. Francaneto and V.A. Forti, 2009. An assessment of mechanical and stink bug damage in soybean seed using X-ray analysis test. Seed Sci. Technol., 37: 110-120.
CrossRefDirect Link - Pritchard, H.W., M.I. Daws, B.J. Fletcher, C.S. Gamene, H.P. Msanga and W. Omondi, 2004. Ecological correlates of seed desiccation tolerance in tropical African dryland trees. Am. J. Bot., 91: 863-870.
CrossRefDirect Link - Rajjou, L., M. Duval, K. Gallardo, J. Catusse, J. Bally, C. Job and D. Job, 2012. Seed germination and vigor. Annu. Rev. Plant Biol., 63: 507-533.
CrossRefDirect Link - Dos Santos, C.E.M., M.A.D.O. Morgado, R.G.P. Matias, A. Wagner Jr. and C.H. Bruckner, 2015. Germination and emergence of passion fruit (Passiflora edulis) seeds obtained by self-and open-pollination. Acta Scientiarum. Agron., 37: 489-493.
CrossRefDirect Link - Dos Santos, S.A., R.F. da Silva, M.G. Pereira, J.C. Machado and C.F. Machado et al., 2009. X-ray technique application in evaluating the quality of papaya seeds. Seed Sci. Technol., 37: 776-780.
CrossRefDirect Link - Shankar, A.C.U., C.S. Nayaka, B.H. Kumar, S.H. Shetty and H.S. Prakash, 2009. Detection and identification of the blackeye cowpea mosaic strain of Bean common mosaic virus in seeds of cowpea from southern India. Phytoparasitica, 37: 283-293.
CrossRefDirect Link - Silva, P.P., R.A. Freitas, S.M. Cicero, J. Marcos-Filho and W.M. Nascimento, 2014. Image analysis to associate morphological and physiological characteristics in pumpkin seeds. Horticultura Brasileira, 32: 210-214.
CrossRefDirect Link - Da Silveira, L.M., M.A. Queiroz, J.A.A. Lima, A.K.Q. Nascimento and I.S.L. Neto, 2009. Serological survey of virus in cucurbit species in the lower middle Sao Francisco river basin, Brazil. Trop. Plant Pathol., 34: 123-126.
CrossRefDirect Link - Simak, M., U. Bergsten and G. Henriksson, 1989. Evaluation of ungerminated seeds at the end of germination test by radiography. Seed Sci. Technol., 17: 361-369.
Direct Link - Stace-Smith, R. and R.I. Hamilton, 1988. Inoculum thresholds of seed borne pathogens: Viruses. Phytopathology, 78: 875-880.
Direct Link - Stoner, W.N., 1963. A mosaic virus transmitted by beetles and a grasshopper. Phytopathology, 53: 890-890.
Direct Link - Sturiao, W.P., P.R.C. Landgraf and T.P. Rosa, 2012. X-ray test for the evaluation of the internal morphology of the seeds of seafortia palm trees. Semina: Ciencias Agrarias, 33: 3003-3008.
Direct Link - Walters, C., 2015. Orthodoxy, recalcitrance and in-between: Describing variation in seed storage characteristics using threshold responses to water loss. Planta, 242: 397-406.
CrossRefDirect Link - Sanfacon, H., T. Iwanami, A.V. Karasev, R. van der Vlugt and J. Wellink et al., 2011. Family Secoviridae. In: Virus Taxonomy: Classification and Nomenclature. Ninth Report of the International Committee on Taxonomy of Viruses, King, A.M.Q., M.J. Adams, E.B. Carstens and E.J. Lefkowitz (Eds.)., Academic Press, San Diego, CA., pp: 881-899.
- Ministerio da Agricultura, Pecuaria e Abastecimento, 2009. Regras Para Analise de Sementes. 1st Edn., Ministerio da Agricultura, Pecuaria e Abastecimento, Secretaria de Defesa Agropecuaria, Brasilia, ISBN: 978-85-99851-70-8, Pages: 399.
Direct Link