
M.A. El-Metwally
Department of Mycological Research, Plant Pathology Research Institute, ARC, Egypt
Aml E.A. El-Saidy
Department of Seed Technology Research, Field Crop Research Institute, ARC, Egypt
K.M. Abd El-Hai
Department of Leguminous and Forage Crop Diseases, Plant Pathology Research Institute, ARC, Egypt
Research Journal of Seed Science
Year: 2014 | Volume: 7 | Issue: 4 | Page No.: 97-115







ABSTRACT
Laboratory and greenhouse experiments were conducted to study the role of some vitamins in decrease of seed deterioration of soybean (Glycine max L. Merrill) during storage and enhance seedling vigor. Seeds stored in cotton bags for 0, 2, 4 and 6 month periods after sprayed with Vitamin C (50, 75 and 100 ppm), Vitamin B9 (10, 15 and 20 ppm) and Vitamin A (50, 100 and 150 ppm). Results indicated that nine known genera of fungi were isolated in seed health test, Fusarium followed by Penicillium recorded the superiority in frequency while Rizoctonia came late after Cephalosporium. Vitamins application decreased growth of different fungal genera in all storage periods, moderate level of Vitamin A was more effective. Germination characters (Germination percentage and its energy and index), seedling parameters (shoot length, root length and seedling dry weight) and seed chemical analysis (oil and protein percentages) were decreased with increasing storage period. In contrast, dead, rotted seeds and rotted seedlings percentage increased significantly with increasing storage periods. Generally, vitamin treatments counteracted the harmful effect of storage on soybean seeds. The application of vitamins as seed treatments increased significantly germination characters, seedling characters and seed chemical analysis as well as survived seed health and minimized dead seeds, rotted seeds and rotted seedlings. Vitamin A was more effective in germination characters while Vitamin B9 was more effective in seedling characters and seeds chemical analysis. The maximum values of shoot length and dry weights of shoot and root were recorded from seed treated with Vitamin A. Moreover, Vitamin C gave the maximum values of chlorophyll a, carotenoids and total phenols. The present investigation recommended using Vitamin A at 100 ppm or Vitamin C at 75 ppm as seed treatment to decrease soybean seed deterioration during storage.
PDF Abstract XML References Citation
Received: June 22, 2014;
Accepted: September 02, 2014;
Published: November 12, 2014
How to cite this article
M.A. El-Metwally, Aml E.A. El-Saidy and K.M. Abd El-Hai, 2014. Role of Some Vitamins in Decreasing of Soybean Seed Deterioration During Storage. Research Journal of Seed Science, 7: 97-115.
URL: https://scialert.net/abstract/?doi=rjss.2014.97.115
URL: https://scialert.net/abstract/?doi=rjss.2014.97.115
INTRODUCTION
Soybean (Glycine max L. Merrill) from one of the most important fabaceous crops all over the world. The embryo of soybean seeds is the storehouse of oil and protein. The oil content of the seed is variable ranging from 19-22% which contains unsaturated fatty acids with no cholesterol. The seed protein is ranging between 30-50% which contain most important amino acids. So, soybean seeds proved more useful than other grain legumes by virtue of its dual propose (oil and protein) which may help in solving the storage of protein and oil in Far East. The seeds are rich in vitamins, calcium and iron as well as about 30% carbohydrate content with total soluble sugars equal to about 10% (Snyder and Kwon, 1987; Abd El-Hai, 2001; Abd El-Hai et al., 2010).
Soybean seed quality is declining faster than seeds of other agronomic crops. Seed deterioration is a major problem due to several pathogens and environmental conditions (temperature and relative humidity) which contributing to the loss of seed viability and vigor during storage (Malvick, 1988; Maheshbabu and Hunje, 2008). About more than one third of pathagens reported to infect soybeans are considered economically important (Sinclair, 1982). Alternaria spp., Fusarium spp., Helminthosporium spp., Curvularia spp., Cladosporium spp., Chaetomium spp., Rhizopus nigricans, Aspergillus spp. and Penicillium spp. are common fungi associated with healthy-looking soybean seeds; they play less role than physiologic factors in causing deterioration of soybean seeds (Oyckan, 1978; Ndimande et al., 1981). Seed vigor reduction is due to decrease in seed quality and percentage and rate of germination is greatly influenced by relative humidity and storage temperature. These factors affect on the presence of microorganisms that can accelerate the process of deterioration (Khaliliaqdam et al., 2012). Protection of seed viability depend on storage conditions and duration (Kandil et al., 2013).
Enhancement of seed vigor is a useful approach to improve overall growth and performance of agricultural crops in order to increase productivity. The use of chemicals in seed treatments is their residual toxicity and adversely affects on human and animals besides affecting on seed quality. Therefore, alternative control methods are needed to nullify the hazards of fungicide on public health and environmental balance. Many authors stated that resistance can be systematically induced by chemical substrates dependent on the host plant physical or chemical barriers (Khalifa et al., 2007; Walters et al., 2007). Today, the most widely used application of seed treatment in the traditional one of protecting the germinating seedling against seed- and soil-borne fungi in the period immediately after planting. There is a positive correlation between the sythesis of secondary metabolites such as phenolics and traditional seed vigor parameters (Randhir and Shetty, 2003). Ascorbic acid (Vitamin C) and folic acid (Vitamin B9 or folecin) are water soluble with antioxidant potential and have the ability to donate protons to cytosol, similar to salicylic acid. They also appear to be suitable growth regulators for increasing seed vigor and phenolic elicitation (McCue et al., 2000). Moreover, both vitamins could potentially serve as indirect stimulators of proline biosynthesis that has been linked to Pentose Phosphate Pathway (PPP) activity during stress (Shetty and Wahlqvist, 2003). Folic acid (folate) is necessary for the production of new cells, for DNA synthesis and RNA synthesis and for preventing changes to DNA, thus for preventing cancer (Kamen, 1997). Vitamin A (Retinol or beta-carotene) has a function in very different role as an irreversible oxidized from of retinol which is an important hormone-like growth factor for cells and plays a role in antioxidant activity (Solomons and Orozce, 2003).
Therefore, this investigation aimed to study the role of certain vitamins (Vitamin C, Vitamin B9 and Vitamin A) in decrease of soybean seed deterioration under different storage periods.
MATERIALS AND METHODS
A storage experiment was conducted at Seed Technology Research Unit Laboratory in Mansoura, Dakahlia Governorate, Field Crop Research Institute, Agricultural Research Center, Egypt. The experiment lasted from November 2012 to May 2013. The aim was to reduce soybean seed deterioration under different storage periods (0, 2, 4 and 6 months) using different concentrations of three vitamins (C, B9 and A). In addition, a pot experiment was carried out for 30 days during summer season of 2013 at greenhouse of the Plant Pathology Department, Faculty of Agriculture, Mansoura University to evaluate the response of soybean to the previously tested treatments after six months of storage.
Source of seeds and tested chemical: Soybean (Giza 22 cultivar) seeds obtained from the field experiment after harvest were used for storage studies, Legumes Crops Research Department, Agricultural Research Center, Egypt. Vitamin C (Ascorbic acid), Vitamin B9 (folic acid) and Vitamin A (Retinol) were obtained from Al-Gomhoria Company, Egypt.
Seed treatments: Samples of seeds were individually sprayed with distilled water (wet control), Ascorbic acid (Vitamin C) at 50, 75 and 100 ppm, Folic acid (Vitamin B9) at 10, 15 and 20 ppm and Retinol (Vitamin A) at 50, 100 and 150 ppm. The untreated seeds (dry seeds) served as control. All treatments were kept at laboratory temperature. After spraying, the seeds were surface-dried in an incubator with forced air circulation for 48 h on filter paper at a temperature of 25°C to return to original moisture 12-14% (on dry weight basis). The seeds were stored in cloth bags (500 g for each one) and kept in laboratory conditions for period of 0, 2, 4 and 6 months.
Laboratory experiment
Seed Health Testing (SHT): Detection of seed-borne mycoflora was carried out following the procedures published by the International Seed Testing Association (ISTA, 1996). Two hundred seeds from each sample and the two controls were tested using the standard blotter. Ten seeds were plated in 11 cm diameter petri-dish containing three layers of water-soaked blotters using sterilized tap water. The plates were incubated at 20±2°C for 7 days under 12 h alternating cycles of cool white fluorescent light and darkness. Plates were examined under a stereoscopic binocular microscope (6-50X) for the presence of seed-borne fungi and to study their habit characters. When necessary, the compound microscope was used for confirming the identification after having examination of morphology of conidia and conidiophores. Fungi occured on seeds were identified by means of comparison with the description sheets of Commonwealth Mycological Institute, Kew, Surrey, England (CMI), Danish Government Institute of Seed Pathology (DGISP) publications as well as publications of (Raper and Fennel, 1965; Ellis, 1971; Chidambaram et al., 1973; Moubasher et al., 1977; Booth, 1985; Burrges et al., 1988; Singh et al., 1991).
Germination characters: Treated and untreated seeds were subjected to standard germination test as the rules of International Seed Testing Association (ISTA,1985). Counts of germinating seeds were taken daily up to eight days after the start of germination. Germination Energy (GE) was recorded as the percentage of germinating seeds at 4th days after plantation. It is relative to the total number of seeds tested (Ruan et al., 2002). The Germination Index (GI) was calculated as described in the Association of Official Seed Analysts (AOSA, 1983) using the following equation:
Disease assessment: The dead seeds and rotted seeds percentage (ungerminated seed) as well as rotted seedlings percentage and survival seedlings were recorded for each storage period.
Seedling characters: Treated and untreated seeds were sown in sterilized sand with the same previous method to determine the seedling characters after eight days. Germination percentage was defined as the total number of normal seedlings at the end of the test. Shoot and root lengths (cm) were determined from 10 normal seedlings and then dried in a forced air oven at 105°C for 24 h to obtain seedlings dry weight (g) under laboratory conditions.
Seed chemical analysis: Seed samples were collected periodically from each treatment were oven dried, finely ground for chemical analysis. Seed oil percentage (%) was determined after extraction with Soxhelt’s apparatus using petrolium hexane as an organic solvent according to AOAC (1998). Seed nitrogen percentage was estimated by using micro Kjeldahel apparatus and multiplied by the converting factor (6.25) to get seed protein percentage (Jackson, 1962).
Greenhouse experiment
Morphological characters: Samples of each treatment were cultured in plastic pots (25 cm diameter) which were filled with 2 kg (2 sand: 1 clay). Ten peanut seeds were sown in 1st May 2013 at 21.3°C and relative humidity 58% in each pot. After germination (eight days), the plants were thinned to leave only three seedlings. The experiment was watered using equal amount of water per pot each time. At the end of the experiment, shoot and root length (cm), fresh weight of shoot and root (g) and dry weight of shoot and root (g) were measured.
Physiological characters: At 30 days from planting, the blade of third leaf from tip was taken to determine photosynthetic pigments (chlorophyll a and b and carotenoids) which were extracted with methanol after adding traces of sodium carbonate (Robinson and Britz, 2000) and determined according to Mackinney (1941). Total phenols were determined in fresh shoot using the folin-ciocalteau reagent according to Singleton and Rossi (1965).
Statistical analysis: Statistical analysis was performed using analysis of variance technique by means of “MSTAT-C” computer software package for the factorial completely randomized design (Laboratory experiment) and randomized complete block design for greenhouse experiment as published by Gomez and Gomez (1984). Using the Least Significant Difference test (LSD), differences were compared among treatment means at levels of 5 and 1% of probability.
RESULTS
Laboratory experiments
Seed health testing: Table 1 shows that nine genera of fungi were isolated from soybean seed samples. They were Alternaria, Aspirgillus, Botrytis, Cephalosporium, Fusarium, Penicillium, Rhizoctonia, Rhizopus and Verticillium. Generally, five genera i.e., Alternaria, Aspirgillus, Botrytis, Penicillium and Rhizopus increased while, other genera decreased in both controls with increasing storage periods from 0-6 months. Taking in consideration that fungal population enhanced in check moist seeds than in check dry seeds. Aspergillus followed by Fusarium then Penicillium involved a large number of isolates obtained while, Rhizoctonia came late after Cephalosporium.
| Table 1: | Frequency (%) of seed-borne fungal population of soyabean seed as affected by the tested some vitamins at different storage periods and their interactions |
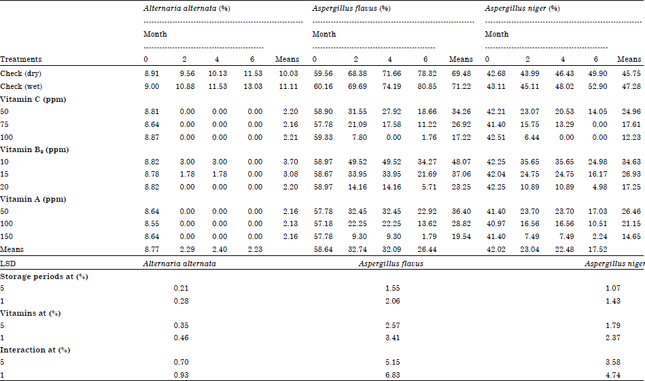 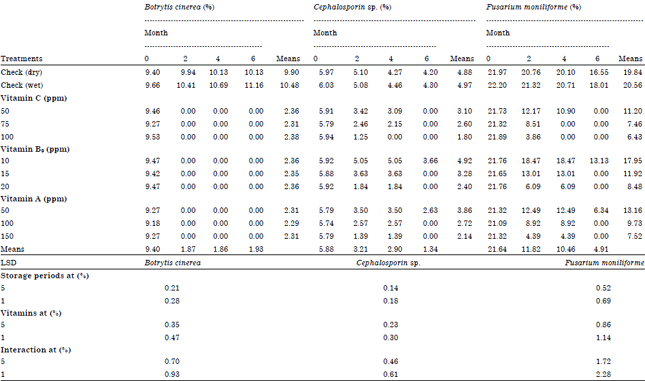 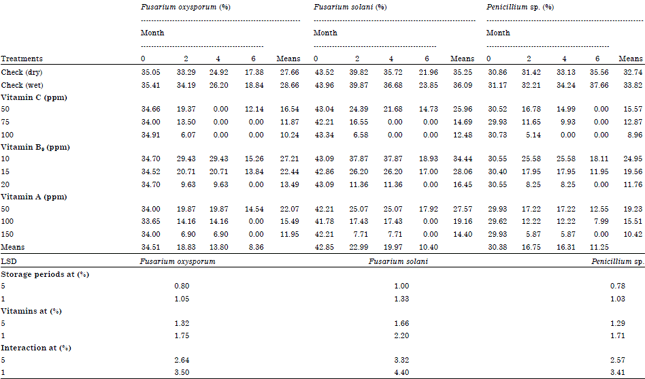 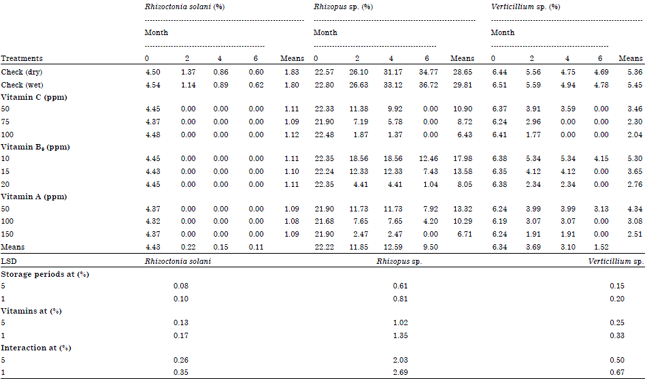 | |
Statistical analysis showed highly significant differences among storage periods, vitamin treatments and its interactions. All treatments gave inhibitory effects on the presence of different fungal genera in all storage periods. In this respect, the moderate concentration of all tested vitamins was more effective than low or high concentration. The maximum inhibitions in fungal populations were recorded with Vitamin A followed by Vitamin C while Vitamin B9 came late. All treatments led to complete inhibition of Rhizoctonia, Botrytis and Alternaria at 2, 4 and 6 month storage periods except low and moderate concentrations of Vitamin B9 which slightly showed the persence of Alternaria at 2 and 4 storage months.
Germination characters: Data in Table 2 shows that the germination percentage as well as its energy and index of soybean seeds decreased significantly with increasing storage period from 0-6 months. On the other hand, seed treated with any concentration of the tested vitamins increased significantly the above germination characters and there are positive relationship between the concentration and their effects. The high level of Vitamin A (150 ppm) gives the highest values of germination percentage followed by the high level of Vitamin C (100 ppm). Moreover, the highest values of germination energy and its index occurred under the application of Vitamin B9 at high level (20 ppm). While, seeds of dry control gave the lowest values of germination characters. The combination between storage periods and any of treatments had no significant effect on germination characters.
Disease assessment: The effects of storage periods, vitamins and their interactions on dead seeds percentage, rotted seeds percentage, rotted seedlings percentage and survival healthy percentage are presented in Table 3 and 4.
Data in Table 3 showed that dead and rotted seeds percentage increased significantly with increasing storage periods. The sharp increase occurred under six month storage period. The highest values of these traits produced from wet control. On contrast, all treatments at any concentration significantly decreased dead and rotted seeds and counteracted the harmful effects of storage periods. In this respect Vitamin C at 75 ppm was more effective followed by moderate concentration of Vitamin A (100 ppm).
The results in Table 4 showed that rotted seedlings percentage was increased significantly by increased of storage periods. The maximum rotted seedlings percentage was recorded at 6 storage month. There is a negative correlation between storage periods and total survival seedlings percentage. All concentrations of any vitamins significantly deceased rotted seedlings consequently increased healthy seedlings. Moderate level of Vitamin C followed by moderate level of Vitamin A then high level of Vitamin C gave the lowest rotted seedlings. While, highest values of survival healthy seedlings percentage occurred under the same treatments. Generally, the application of vitamins as soybean seed treatments decreased the depressing effects of storage periods on seeds and seedlings of soybean.
Seedling characters: Table 5 indicated that the shoot length, root length and dry weight of soybean seedlings were decreased as storage periods were increased. The maximum reduction in these parameters occurred after six months from storage. All tested vitamins clearly increased seedling characters compared with both controls (dry and wet). The highest values of shoot length was recorded by the application of Vitamin B9 at high level followed by high level of Vitamin A then high level of Vitamin C.
| Table 2: | Germination characters of soybean seed as affected by some vitamins treated at different storage periods and their interactions |
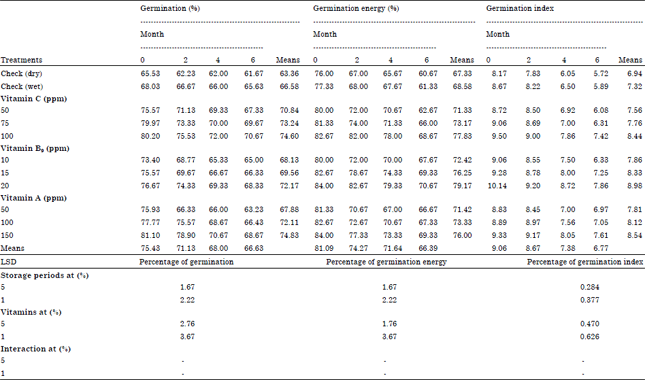 | |
| Table 3: | Dead and rotted soybean seeds as affected by the tested some vitamins at different storage periods and their interactions |
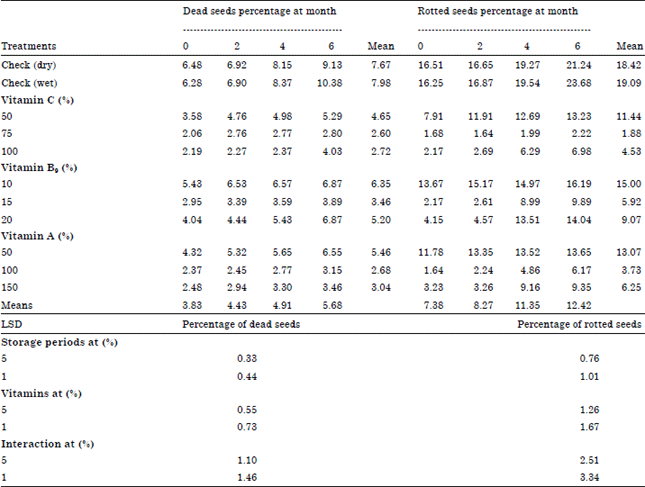 | |
Moreover, the high level of Vitamin C gave the highest values of root length followed by high level of Vitamin B9. Treatment of Vitamin A at high level recorded highest seedlings dry weight followed by Vitamin B9 then Vitamin C. Meanwhile, the interaction between storage periods and vitamin treatments had no significant effect on seedling characters.
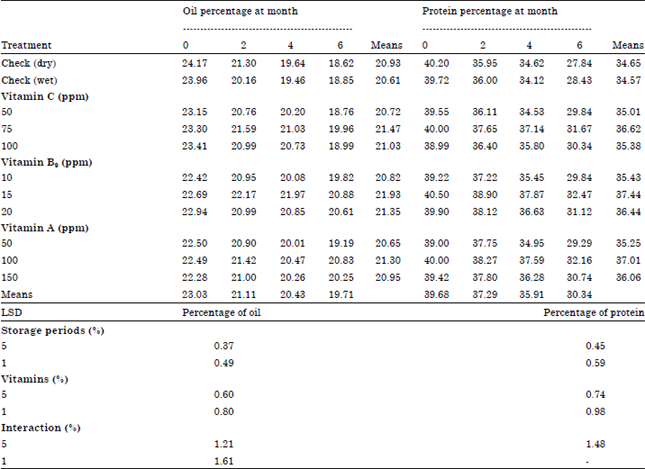
Seed chemical analysis: Results in Table 6 showed that significant decrease in oil and protein percentage in soybean seeds with increase of storage periods from 0-6 months. On the other hand, all levels of vitamins increased significantly soybean seed oil and protein contents. The moderate level of Vitamin B9 followed by moderate Vitamin C level recorded maximum oil content. Moreover, the highest values of protein occurred under the application of moderate level of Vitamin B9 followed by Vitamin A then Vitamin C. The interaction between storage periods and vitamins significantly increased soybean seed oil and protein content. Generally, vitamin treatments counteracted the harmful effect of storage periods on soybean seed quality.
Greenhouse experiment: In this experiment, samples of each treatment were taken after six months storage and planted in plastic pots to study the role of vitamins on the following characters:
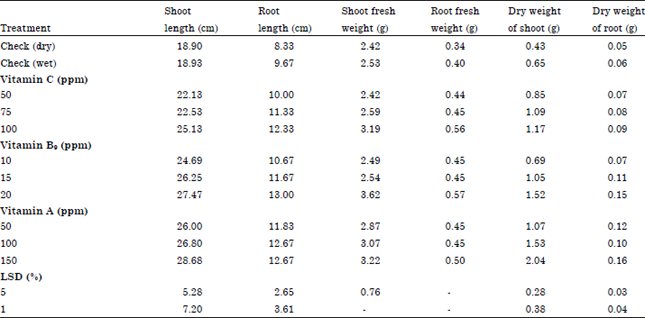
| • | Seedling morphological characters: The effect of soybean seed treatments with vitamins for six month storage periods on seedling morphological characters (shoot length, root length, fresh and dry weight of shoot and root) is presented in Table 7 |
| Table 4: | Rotted and survival seedlings as affected by the tested some vitamins at different storage periods and their interactions |
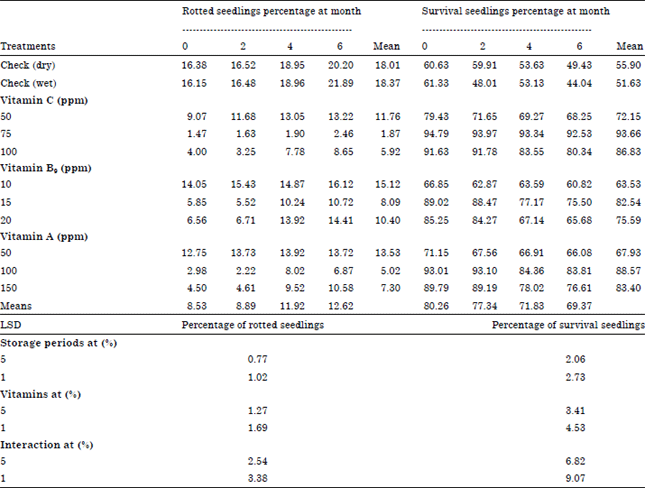 | |
It can be observed that all treatments at any level used significantly the above mentioned characters except, root fresh weight which showed no significant effect. Taking in consideration that there are positive relationship between the concentrations and their effect on seedling morphological characters. The highest values of shoot length and dry weight of shoot and root appeared under application of Vitamin A followed by Vitamin B9 but Vitamin B9 followed by Vitamin A were the most effective on root length and shoot fresh weight.
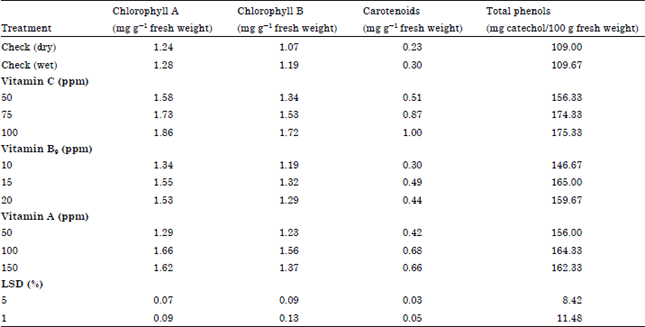
| • | Physiological characters: Concerning the effects of vitamins on photosynthetic pigments (chlorophyll a, b and carotenoids) and total phenols in Table 8 showed that all treatments at any level used significantly increased photosynthetic pigments and total phenol content except, chlorophyll b and carotenoids under low level of Vitamin B9 (10 ppm) were found to be similar to that of wet control. High and moderate levels of Vitamin C (100 and 75 ppm) gave the highest values of chlorophyll a, carotenoids and total phenols |
| Table 5: | Variation in morphological characters of soybean seedlings as affected by the tested some vitamins at different storage periods and their interactions |
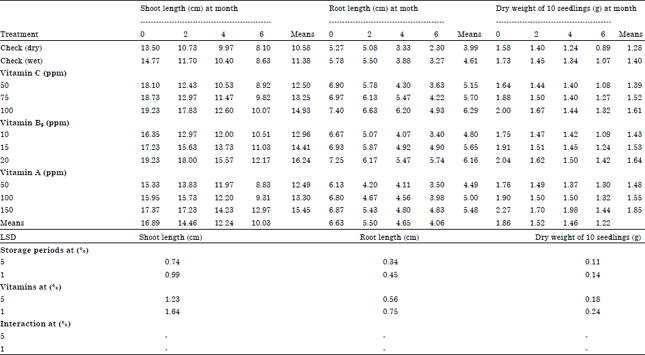 | |
| Table 6: | Chemical composition of soybean seeds as affected by the tested some vitamins at different storage periods and their interactions |
 | |
| Table 7: | Variation in morphological characters of soybean seedlings under greenhouse as a function of the tested some vitamins |
 | |
| Table 8: | Variation in physiological characters of soybean seedling under greenhouse as a function of the tested some vitamins |
 | |
DISCUSSION
The impact of environmental regulations that have either banned or restricted the use of older fungicides and the development of biological and chemical control of plant pathogens and provide plant protection will into the growing season so, the uses and expectations of seed treatments are greater.
Soybean is listed as poor store due to a quick deterioration by auto oxidation of lipids and increase of content of free fatty acids during storage period which led to decrease in seed viability and vigor under warm and humid condition (Maheshbabu and Hunje, 2008).
The rapid loss of seed viability and vigor during storage is related to environmental conditions during seed production, pests, diseases, seed oil content, seed moisture content, mechanical damages of seed in processing, storage longevity, packaging, pesticides, air temperature, relative air humidity in storage and biochemical injury of seed tissue (Guberac et al., 2003; Heatherly and Elmore, 2004). In old seeds, the loss of germination and vigor is positively correlated with the electrolytic leaching which increases with the decrease of phospholipids content of the membrane (Vieira et al., 2002).
Two types of fungi invade seeds: field fungi and storage fungi. The major deleterious effects of storage fungi are to: Decrease viability, cause discoloration, produce mycotoxins, cause heat production and develop mustiness and caking (Bewley and Black, 1994). Intrinsic nutritional factors, extrinsic factors, processing factors and implicit factors affect fungal population that could change throughout the storage period followed by seed deterioration (Magan et al., 2004).
The inhibitory effects of storage periods on germination shoot length, root depth and seedling dry weight might be due to their genetic differences age induced deterioration, inherent differences in seed structure and composition as well as seed quality (Kandil et al., 2013). In addition, seed storage would increase the metabolic activity of seeds and decrease the reserve substance content due to oxidation of amino acids, increase in respiratory activity and advance in the deterioration process of stored seeds (Bewley and Black, 1994).
In this investigation, seed treatment with vitamins overcome the depressing effects of storage periods on germination characters and seedling morphological characters through its effects are decreasing on seed fungal population, increasing seed quality as well as increasing photosynthetic pigments and phenol content. Chlorophyll and phenol compounds are a good parameters reflecting the health condition of plant and induce the resistance (Reuveni et al., 1992). Also, a photosynthetic pigment increases carbohydrate contents which comprises structural polysaccharides and pectin as well as phenol compounds. These compounds play important role in plant defense (Hahlbrock and Scheel, 1989). Moreover, a rapid accumulation of phenols at the infection site is the first step of the defense mechanism in plant which restricts or slows the pathogen growth (Gogoi et al., 2001). In addition, vitamins increase carotenoids content which act as a major endogenous plant antioxidant.
Vitamin B complex and Vitamin C appear to be suitable growth regulators for increasing seed vigor and phenolic elicitation due to their inherent antioxidant potential (McCue et al., 2000). Total phenols have been considered as important defense-related compounds (Gogoi et al., 2001; Khaleifa et al., 2006). Phenolics play important role in the regulation of plant metabolic processes, plant growth and lignin synthesis (Lewis and Yamamoto, 1990). Burguieres et al. (2007) reported that the treatment of pea seeds with folic acid or Vitamin C enhances phenolic content. Vitamins also could potentially serve as indirect stimulators of proline biosynthesis that has been linked to pentose phosphate pathway activity during stress (Shetty and Wahlqvist, 2003). Proline used as storage compound for energy, hence reduced carbon needs.
Generally, the positive effect of vitamins may be due to its role as antioxidant. Which intercept and neutralize the harmful effect of destructive oxygen free radicals (Benzie, 2003). The antimicrobial action of vitamins as antioxidants is due to inhibition of several enzymes function by oxidized compounds and interferes with protein, RNA and DNA synthesis (Nesci et al., 2003). There was an increase in Super Oxide Dismutase (SOD) activity and Catalase (CAT) with stimulation of phenolic-linked antioxidant and enhancement of seed vigor under treated with folic acid and Vitamin C (Burguieres et al., 2007). Catalase and superoxide dismutase are important antioxidant enzymes with the ability to repair oxidation damage caused by reactive oxygen species and in the decomposition of hydrogen peroxide to give water and oxygen (Babilha et al., 2002; Apel, 2004).
CONCLUSION
It could be concluded that the application of Vitamin A at 100 ppm or Vitamin C at 75 ppm as seed treatment is recommended for reducing soybean seed deterioration during storage.
REFERENCES
- Abd El-Hai, K.M., M.A. El-Metwally and S.M. El-Baz, 2010. Reduction of soybean root and stalk rots by growth substances under salt stress conditions. Plant Pathol. J., 91: 149-161.
CrossRefDirect Link - AOSA., 1983. Seed Vigor Testing Handbook: Contribution No. 32 to the Handbook on Seed Testing. Association of Official Seed Analysis (AOSA), Springfield, IL., USA., Pages: 93.
Direct Link - Maheshbabu, H.M. and R. Hunje, 2008. Effect of seed treatment with botanicals on storability of soybean. Karnataka J. Agric. Sci., 21: 357-360.
Direct Link - Benzie, I.F.F., 2003. Evolution of dietary antioxidants. Comp. Biochem. Physiol. A: Mol. Integr. Physiol., 136: 113-126.
CrossRefPubMedDirect Link - Burguieres, E., P. McCue, Y.I. Kwon and K. Shetty, 2007. Effect of vitamin C and folic acid on seed vigour response and phenolic-linked antioxidant activity. Bioresour. Technol., 98: 1393-1404.
CrossRefDirect Link - Gogoi, R., D.V. Singh and K.D. Srivastava, 2001. Phenols as a biochemical basis of resistance in wheat against karnal bunt. Plant Pathol., 50: 470-476.
CrossRefDirect Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedures for Agricultural Research. 2nd Edn., John Wily and Sons, Hoboken, New Jersey, ISBN: 9780471870920, Pages: 704.
Direct Link - Guberac, V., S. Maric, A. Lalic, G. Drezner and Z. Zdunic, 2003. Hermetically sealed storage of cereal seeds and its influence on vigour and germination. J. Agron. Crop Sci., 189: 54-56.
CrossRefDirect Link - Hahlbrock, K. and D. Scheel, 1989. Physiology and molecular biology of phenylpropanoid metabolism. Annu. Rev. Plant Physiol. Plant Mol. Biol., 40: 347-369.
CrossRefDirect Link - Kandil, A.A., A.E. Sharief and M.S. Sheteiwy, 2013. Seedling parameters of soybean cultivars as influenced with seed storage periods, conditions and materials. Int. J. Agric. Sci., 5: 330-338.
CrossRefDirect Link - Khaleifa, M.M.A., C.R. Azzam and S.A. Azer, 2006. Biochemical markers associated with disease resistance to damping-off and root-rot diseases of peanut mutants and their productivity. Egypt. J. Phytopathol., 34: 53-74.
Direct Link - Khaliliaqdam, N., A. Soltani, N. Latifi and F.G. Far, 2012. Quantitative response of the longevity of soybean seed under controlled conditions. Am. Eurasian J. Agric. Environ. Sci., 12: 224-230.
Direct Link - Lewis, N.G. and E. Yamamoto, 1990. Lignin: Occurrence, biogenesis and biodegradation. Annu. Rev. Plant Physiol., 41: 455-496.
CrossRefDirect Link - Mackinney, G., 1941. Absorbtin of light by chlorophyll solution. J. Biol. Chem., 140: 315-322.
Direct Link - McCue, P., Z. Zheng, J.L. Pinkham and K. Shetty, 2000. A model for enhanced pea seedling vigour following low pH and salicylic acid treatments. Process Biochem., 35: 603-613.
CrossRef - Ndimande, B.N., H.C. Wien and E.A. Kueneman, 1981. Soybean seed deterioration in the tropics. I. The role of physiological factors and fungal pathogens. Field Crops Res., 4: 113-121.
CrossRefDirect Link - Nesci, A., M. Rodriguez and M. Etcheverry, 2003. Control of Aspergillus growth and aflatoxin production using antioxidants at different conditions of water activity and pH. J. Applied Microbiol., 95: 279-287.
CrossRefDirect Link - Reuveni, R., M. Shimoni, Z. Karchi and J. Kuc, 1992. Peroxidase activity as abiochemical marker of resistance of muskmelon (Cucumic melon) to Pseudoperonospora cubensis. Phytopathology, 82: 749-753.
Direct Link - Robinson, J.M. and S.J. Britz, 2000. Tolerance of a field grown soybean cultivar to elevated ozone level is concurrent with higher leaflet ascorbic acid level, higher ascorbate-dehydroascorbate redox status and long term photosynthetic productivity. Photosynth. Res., 64: 77-87.
CrossRefDirect Link - Randhir, R. and K. Shetty, 2003. Light-mediated fava bean (Vicia faba) response to phytochemical and protein elicitors and consequences on nutraceutical enhancement and seed vigour. Process Biochem., 38: 945-952.
CrossRef - Ruan, S., Q. Xue and K. Tylkowska, 2002. The influence of priming on germination of rice (Oryza sativa L.) seeds and seedling emergence and performance in flooded soil. Seed Sci. Technol., 30: 61-67.
Direct Link - Shetty, K. and M.L. Wahlqvist, 2003. A model for the role of the proline-linked pentose-phosphate pathway in phenolic phytochemical bio-synthesis and mechanism of action for human health and environmental applications. Asia Pac. J. Clin. Nutr., 13: 1-24.
PubMed - Singleton, V.L. and J.A. Rossi, 1965. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic., 16: 144-158.
CrossRefDirect Link - Solomons, N.W. and M. Orozco, 2003. Alleviation of vitamin A deficiency with palm fruit and its products. Asia Pac. J. Clin. Nutr., 12: 373-384.
PubMedDirect Link - Vieira, R.D., A.L. Penariol, D. Perecin and M. Panobianco, 2002. Electrical conductivity and initial water content of soybean seeds. Pesquisa Agropecuaria Brasileira, 37: 1333-1338.
CrossRef