
K.P. Yoga
Department of Seed Science and Technology, Tamil Nadu Agricultural University, Coimbatore, 641 003, Tamil Nadu, India
R. Jerlin
Department of Seed Science and Technology, Tamil Nadu Agricultural University, Coimbatore, 641 003, Tamil Nadu, India
M. Ameer Junaithal Begum
Department of Seed Science and Technology, Tamil Nadu Agricultural University, Coimbatore, 641 003, Tamil Nadu, India
Research Journal of Seed Science
Year: 2014 | Volume: 7 | Issue: 1 | Page No.: 21-25







ABSTRACT
Effect of different growth retardants viz., MH at 5000, 10000, 15000 ppm, ccc at 1000, 2000, 3000 ppm and ABA at 250 and 500 ppm on inducing dormancy in non-dormant groundnut cv. TMV 7 was studied by foliar application at 70 and 80 days after sowing. It was evident from the results that ABA 500 ppm applied at 70 days after sowing was most effective in inducing dormancy in the resultant seeds as well as recorded the lowest in situ germination of pods.
PDF Abstract XML References Citation
Received: September 12, 2013;
Accepted: October 29, 2013;
Published: February 27, 2014
How to cite this article
K.P. Yoga, R. Jerlin and M. Ameer Junaithal Begum, 2014. Standardization of Dormancy Induction Treatments in Groundnut cv. TMV 7. Research Journal of Seed Science, 7: 21-25.
URL: https://scialert.net/abstract/?doi=rjss.2014.21.25
URL: https://scialert.net/abstract/?doi=rjss.2014.21.25
INTRODUCTION
Dormancy is the natural evolutionary consequence, however, depending on the situation dormancy may be a boon or evil in groundnut. In general, in groundnut bunch types are non-dormant while spreading and semi spreading types are having a varied period of dormancy.
The groundnut crop by its very nature of fruiting underground cannot be harvested without sufficient moisture in the soil. Rain at the time of harvest are often either too low resulting in imperfect harvest at increased cost or excessive when harvesting has to be delayed till the fields come to a workable condition. In the latter circumstances, considerable loss by the sprouting of the seed occurs in the bunch varieties. The fully germinated seed is quite useless while the partially sprouted one is of poor quality with low oil content and has a tendency for rapid deterioration. Specially in black soil regions, the higher percentage of sprouting of pods is commonly noticed due to high moisture retention capacity of the soil. About 20% loss in bunch groundnut pod yields has been reported due to in situ germination (Nagarjun and Radder, 1983). Eighty percent of groundnut seeds had germinated in the field due to heavy rain (John et al., 1948) and 20-50% pods germinate due to rains at pod maturity stage. Therefore, the non-dormant nature of seed in bunch groundnut is considered to be the major problem in its cultivation. To overcome this kind of situation, in most of the bunch groundnut growing areas, there is an important need to search for a method of inducing seed dormancy to save the produce and to retain the seed quality against field sprouting.
MATERIALS AND METHODS
To induce dormancy in non-dormant cv. TMV 7, the plants were raised in pots and were sprayed with different growth retardants at 70 and 80 days after sowing in addition to control which was not sprayed with growth retardants.
Treatments
| T0 | = | Unsprayed control |
| T1 | = | Maleic hydrazide (MH) 5000 ppm |
| T2 | = | Maleic hydrazide (MH) 10000 ppm |
| T3 | = | Maleic hydrazide (MH) 15000 ppm |
| T4 | = | Cycocel (CCC) 1000 ppm |
| T5 | = | Cycocel (CCC) 2000 ppm |
| T6 | = | Cycocel (CCC) 3000 ppm |
| T7 | = | Abscisic acid (ABA) 250 ppm |
| T8 | = | Abscisic acid (ABA) 500 ppm |
Stages of application
| S1 | = | 70 days after sowing |
| S2 | = | 80 days after sowing |
The experiment was conducted as factorial completely randomized design at the Department of Seed Science and Technology, Tamil Nadu Agricultural University, Coimbatore. After the attainment of maturity, the plants were left in pots for 20 days with regular watering for germination of pods in situ. After 20 days, the pods were lifted and the number of pods sprouted in each treatment were counted. Germination test was conducted in sand medium using 4x100 seeds and germination percent of resultant seeds were taken at 10th day.
RESULTS AND DISCUSSION
In groundnut, bunch type cultivars lack seed dormancy which cause considerable loss due to sprouting of pods in the field. The loss due to this pre-harvest sprouting is nearly 20-40% depending upon the variety as reported by John et al. (1948). To overcome this kind of loss, the present experiment was conducted to find out a suitable method for inducing seed dormancy so as to save the produce from sprouting in the field. To induce the dormancy, the crop of non-dormant groundnut cultivar TMV 7 was sprayed with Maleic Hydrazide (MH), Cycocel (CCC) and Abscisic Acid (ABA) at different concentrations in different days viz., 70 and 80 days after sowing. From the results, it is evident that dormancy was induced in groundnut cv. TMV 7 by spraying of ABA 500 ppm on 70 days after sowing, MH 15000 ppm on 70 days after sowing and CCC 3000 ppm on 80 days after sowing. The above treatments recorded the lowest in situ germination of 2% while unsprayed control recorded the maximum of 44% (Table 1). ABA 500 ppm applied at 70 days after sowing registered superiority over other treatments in inducing dormancy of resultant kernels to the extent of 81% over control while the resultant kernels of control plants recorded 93% germination (Fig. 1).
The result of the present study was evinced by Ketring (1973) and Narasimha Narasimhareddy and Swamy (1976) by expressing the inhibitory action of ABA on germination of groundnut seed. Sengupta et al. (1979) observed the inhibition of groundnut seed germination by ABA and CCC but the extent of inhibition was more due to ABA than CCC. The inhibitory effect imposed by ABA was reversed by kinetin application and they concluded that ABA and kinetin control the seed dormancy and germination in groundnut, respectively. ABA is a naturally occuring organic substance that inhibit seed germination (Ketring, 1973). Veldstra (1945) proposed that the inhibition of germination would result from changes in membrane permeability.
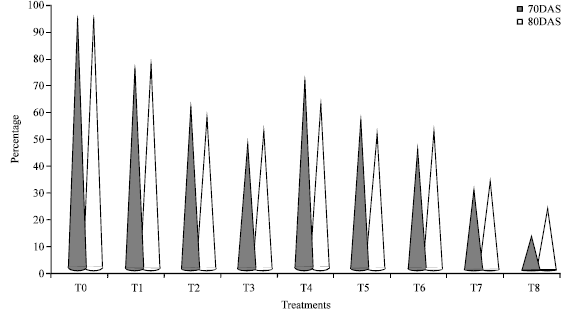 | |
| Fig. 1: | Effect growth retardants on germination (%) of resultant seeds of groundunt cv. TMV 7 |
| Table 1: | Effect of foliar application of growth retardants on in situ germination (%) of groundnut cultivar TMV 7 |
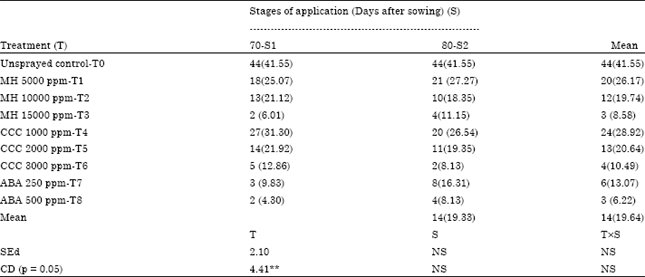 | |
| Figures in parenthesis indicate arcsine values | |
ABA inhibited groundnut seed germination and the factor which appeared limiting in the presence of ABA was low level of ethylene production by the seed. Thus, ABA may prevent germination and cause dormancy by preventing ethylene synthesis (Ketring and Morgan, 1971). ABA is an auxin antagonist and the mode of action of this auxin antogonisms have been shown to be non competitive. The suggested mechanisms of action for the observed inhibition of growth are: Combination of IAA with a protein and competition at a particular site by the structurally similar trans-cinnamic acid; formation of loose combination or easily dissociated coumarin compounds of enzymes or plant cell metabolites; reactions with sulf hydryl groups of enzymes by the unsaturated lactones, coumarin and protoanemonin. Thus, the machanism of action of the inhibitors in preventing growth and germination was at the molecular level (Ketring, 1973). The stimulation of plant growth and seed germination by GA is counteracted or inhibited by ABA. Haber et al. (1969) have shown that the promotion of germination by GA was due to the expansion of existing cells and this expansion was inhibited by ABA. Chen and Osborne (1970) stated that GA activates preformed mRNA stored in the embryo and ABA blocks the existing messages but perhaps at sites different from those of GA. GA did not stimulate DNA synthesis but ABA inhibited it once it was initiated. The inhibition of expression of ot-amylase genes by ABA had been reported by Pagano et al. (1997) in barley. Le Page-Degivry et al. (1990) observed that the dormancy of Helianthus annus appeared to be dependent on ABA synthesis but not concomitant with its accumulation. Seshu and Dadlani (1991) reported that seed dormancy in rice is a result of accumulation of ABA in the hull and pericarp during maturation which strongly inhibits amylase activity in the germinating seeds. ABA inhibit cell division and cell elongation and this could occur by breaking synthesis of a particular RNA molecule (Ketring, 1973).
The inhibitory effect of CCC on germination of groundnut seed in the present study is in continuity with Ketring (1973) and they reported that CCC is a synthetic growth retardant which inhibits seed germination. In the presence of CCC, diffusible auxin from pea plant apices was reduced. CCC prevents synthesis of a GA like substances in non-dormant embryo of wild oats, wheat, barley. Inhibition of groundnut seed germination by CCC was reported earlier by Sengupta et al. (1979) which was in confirmation with the present study. The induction of seed dormancy in groundnut by MH in the present investigation was in confirmity with Gupta et al. (1985). Nagarajan and Gopalakrishnan (1958) reported that the presence of auxin like substances in the groundnut seed is responsible for germination and making it non-dormant. The introduction of antiauxins to the seed by means of foliar application at the time of kernel development may suppress the auxin formation and induce dormancy. Maleic hydrazide (diethanolamine salt of 1,2-dihydroxy-3,6-pyridazinedione) a growth and respiratory inhibitor, possesses the characteristics of an anti-auxin, has been found to be capable of inducing dormancy by antagonizing with auxin in groundnut. Since MH is an auxin-antagonist, the primary effect of MH on inducing dormancy seems to be through interference in the tryptophan metabolism, as the tryptophan is the precursor in the synthesis of auxins (Karivaratharaju and Rao, 1972). Besides this, MH was found to increase the content of another amino acid, hydroxy proline which inhibit the auxin induced cell elongation. Nagarjun and Radder (1983) observed that the moisture content and catalase enzyme activity reduced during induction of dormancy by MH. Seed germination is an energy requiring process and is therefore dependent upon the moisture content of the seed. In addition to this, dormancy is also related to control of enzyme formation and inactivation of seed and they concluded that the reduction in moisture content of seed also indicates that the application of MH prevented the pods to absorb moisture and thereby it inhibits imbibition which is the primary phase in seed germination.
REFERENCES
- Gupta, R.K., S.S. Singh and M.M. Verma, 1985. Introduction of dormancy in groundnut variety T64 by Maleic Hydrazide. Indian J. Agric. Res., 19: 82-86.
Direct Link - Haber, A.H., D.E. Foard and S.W. Perdue, 1969. Actions of gibberellic and abscisic acids on lettuce seed germination without actions on nuclear DNA synthesis. Plant Physiol., 44: 463-467.
Direct Link - John, C.M., C.R. Seshadri and Bhavani M. Shankar Rao, 1948. Dormancy of the seed in the groundnut. Madras Agric. J., 35: 159-167.
Direct Link - Ketring, D.L. and P.W. Morgan, 1971. Physiology of oil seeds II. Dormancy release in virginia-type peanut seeds by plant growth regulators. Plant Physiol., 47: 488-492.
Direct Link - Le Page-Degivry, M.T., P. Barthe and G. Garello, 1990. Involvement of endogenous abscisic acid in onset and release of Helianthus annus embryo dormancy. Plant Physiol., 92: 1164-1168.
CrossRefPubMedDirect Link - Nagarjun, P. and G.D. Radder, 1983. Studies in induction of seed dormancy in Bunch type groundnut. Seed Res., 11: 24-31.
Direct Link - Narasimhareddy, S.B. and P.M. Swamy, 1976. Effect of various growth regulators on the germination of dormant groundnut (Arachis hypogaea) seeds. Indian J. Plant Physiol., 19: 226-229.
Direct Link - Sengupta, U.K., M. Pandey and G.S. Sirohi, 1979. Effect of growth regulators on seed germination in groundnut: Interaction studies. Seed Res., 7: 114-119.
Direct Link - Seshu, D.V. and M. Dadlani, 1991. Mechanism of seed dormancy in rice. Seed Sci. Res., 1: 187-194.
CrossRefDirect Link