
V. Manonmani
Department of Seed Science and Technology, Tamil Nadu Agricultural University, Coimbatore, 641 003, Tamil Nadu, India
M. Ameer Junaithal Begum
Department of Seed Science and Technology, Tamil Nadu Agricultural University, Coimbatore, 641 003, Tamil Nadu, India
M. Jayanthi
Department of Seed Science and Technology, Tamil Nadu Agricultural University, Coimbatore, 641 003, Tamil Nadu, India
Research Journal of Seed Science
Year: 2014 | Volume: 7 | Issue: 1 | Page No.: 1-13







ABSTRACT
Seed priming is a commercially used technique for improving seed germination and vigour. It is a technique which involves uptake of water by the seed followed by drying to initiate the early events of germination upto the point of radicle emergence. The benefits include rapid, uniform and increased germination, improved seedling vigour and growth under a broad range of environments resulting in better stand establishment. The common feature in these priming is the controlled uptake of water. Also, the seeds used for priming elicit specific subcellular responses. This review article summarises the recent information available on the various physiological and subcellular processes associated with priming which lead to seed enhancement.
PDF Abstract XML References Citation
Received: September 14, 2013;
Accepted: October 30, 2013;
Published: February 27, 2014
How to cite this article
V. Manonmani, M. Ameer Junaithal Begum and M. Jayanthi, 2014. Halo Priming of Seeds. Research Journal of Seed Science, 7: 1-13.
URL: https://scialert.net/abstract/?doi=rjss.2014.1.13
URL: https://scialert.net/abstract/?doi=rjss.2014.1.13
INTRODUCTION
Crop production depends heavily on planting of high quality seeds. Rapid and uniform emergence is almost important, it is the foundation on which stand establishment is based and potential yield is determined. Absolute longevity depends on initial seed quality, vigour and proper storage. In ancient days, various seed treatments were practiced as initial production techniques for improved productivity. One programmatic approach to increase crop production is seed invigouration (Farooq et al., 2006). Seed invigouration strategies include hardening, osmohardening, osmoconditioning, hydropriming, hormonal priming, matri-priming and others (Windauer et al., 2007). In the last two decades, seed priming is an effective seed invigouration method has become a common seed treatment to increase the rate and uniformity of emergence and crop establishments in most of the crops especially in advanced countries. Seed priming is a controlled hydration process that involves exposing the seeds to low water potentials that restrict germination, but permits pregerminative physiological and biochemical changes to occur (Khan, 1992). Upon rehydration, primed seeds may exhibit faster rates of germination, more uniform emergence, greater tolerance to environmental stress and reduced dormancy in many species (Khan, 1992).
Several literatures (Windauer et al., 2007) revealed that seed priming could advance germination, improve the initial quality characters, improve field emergence, better establishment, crop stand and increase yields in many diverse environments. Generally, seed storage caused a decrease in the protein content which may be related to oxidation of the amino acids, due to the increase in the respiratory activity and advance in the deterioration process of the stored seeds. Seed deterioration is associated with loss of membrane integrity, changes in enzymatic activities, decline in protein and nucleic acid synthesis and lesions in DNA (McDonald, 1999). Priming can reverse some of the aging-induced deteriorative events and thus improve seed performance (Taylor et al., 1998). Priming is responsible to repair the age related cellular and subcellular damage of low vigor seeds that may accumulate during seed development (Bray, 1995). Priming of seed promotes germination by repair of the damaged proteins, RNA and DNA (Koehler et al., 1997). Many seed priming treatments have been used to reduce the damage of aging and invigourate their performance in many crops (Farooq et al., 2009).
However, a strong reduction in longevity is generally associated with primed seeds in many species, eventhough longevity is enhanced in some cases (Taylor et al., 1998). The exact causes of faster deterioration of primed seeds remain unclear and still not established. Decreased longevity of primed seeds has been attributed to reduced DNA repair upon subsequent germination (Redfearn and Osborne, 1997). The promotion of germination with seed priming may take place for several reasons, but changes in metabolic levels are important during seed priming. It has been assumed that the onset of germination is associated with a rapid resumption of RNA and protein biosynthesis (Osborne, 1983). Limited numbers of investigations into the biochemical changes occurring in primed and subsequently germinated seeds have noted changes of pattern of RNA, protein and DNA synthesis (Dell'Aquila and Taranto, 1986). The goal of this article is to update the information on physilogical and biochemical basis of seed priming including recent references which have not been covered by earlier reviews.
Effect of seed priming on physiological seed quality parameters: In seed management, the success in application of any seed management technique depends upon the type of test, method of application, selection of crop, initial performance of the crop, selection of chemical, its concentration, duration of treatment and the purpose of implication. Seed priming is one of the pre-sowing seed management techniques where the seeds are partially soaked and subsequently dried back for invigorative effect that expresses on field emergence and extend up to yield. In rainfed areas, germination and subsequent seedling growth can be inhibited by adverse conditions in the field. Priming is helpful in reducing the risk of poor stand establishment under a wide range of environmental conditions. Researchers had evaluated the influence of various priming treatments in different crops and recommended the suitable seed priming techniques both for field establishment and for improved productivity even under wider variations of growing atmosphere.
Cantliffe et al. (1984) primed lettuce seed (Lactuca sativa L. cv. Minetto) in aerated solutions of 1% K3PO4 and water at 15°C in the dark for various periods of time and reported that priming for 20 h in 1% K3PO4 improved the germination up to 86% . They also reported that cell division occurred at 21 h in water and at 27 h in 1% K3P04 prior to radicle protrusion. Bradford (1986) revealed that seed priming has been successfully demonstrated to improve germination and emergence in many crops particularly seeds of vegetables and small seeded grasses. Furutani et al. (1986) expressed that onion seeds primed for 8 days at 10°C in -1.1 MPa of mannitol reduced the days to 50% germination by 46%. Odell and Cantliffe (1986) reported that tomato seeds primed with 1.5% K3PO4+1% KNO3 solution at 35°C had germinated more rapidly as compared to unprimed seeds. Basra et al. (1988) primed the maize seed in solutions of 2.5% K2HPO4 and 2.5% K2HPO4+KNO3 found that germination accelerated at a chilling temperature and the effect of priming was largely retained after seeds had been dried back. Cantliffe (1991) revealed that lettuce seeds primed in one percent K3PO4 for 20 h in the dark with the addition of 100 ppm of cytokinin reduced thermo dormancy and increased germination percentage. In summer squash Cavallaro et al. (1994) primed the seeds in aerated solution of KNO3+K2HPO4 at 3% for 2 days revealed that priming improved the seedling emergence (97%) where as untreated seeds recorded 72% emergence. Anuradha et al. (1995) revealed that the hydration of freshly harvested cabbage seeds for two days at low temperature (10°C) had increased the emergence percent, speed of germination and days to 50% germination than the control.
Harris et al. (1999) revealed that on-farm seed priming (soaking seeds overnight in water) markedly improved the establishment and early vigor of maize resulting in faster development, earlier flowering, maturity and higher yields. McDonald (2000) reported that sunflower seeds primed with osmotic solution of PEG 800 improved the seedling length and dry mass of both shoot and root. Subbarao et al. (2001) reported that priming of black gram seeds with sodium molybdate (0.5 g L-1) improved the seed yield of rice fallow situation in 19 trials conducted in 5 villages in Jharkhand and West Bengal. Maize seeds were subjected to hydropriming, osmotic priming (PEG-10,000) and matriconditioning with compost, pressmud and GA3. Results revealed that early germination was recorded in seeds matriconditioned with compost, pressmud and GA3 in laboratory and early emergence under field condition (Afzal et al., 2002). Kaur et al. (2003) observed that priming chickpea seeds with water caused early germination, increased seedling length, maximum biomass of root and shoot under salt stressed conditions when compared to non-primed seeds. Demer and Mavi (2004) haloprimed the watermelon seeds in KNO3 3% for 6 days at 20°C and found that priming decreased mean emergence time and increased seedling weight and hypocotyl length. Subedi and Ma (2005) evaluated the effect of seed priming with water, osmotic solution (2.5% KCl) and plant growth regulators (Indole acetic acid, cytokinin, ethephon and gibberellic acid) on emergence, seedling vigor, nitrogen response and grain yield of corn. The results revealed that seed soaking with 20 ppm gibberellic acid (GA3) solution for 30 min improved the seedling vigour.
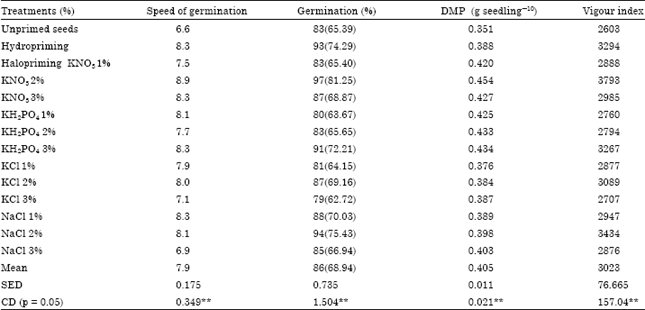
Basra et al. (2003a) in wheat (Triticum aestivum cv. Auqa) found that hydropriming for 24 h and hardening for 12 h (one cycle hydropriming, hardening and matriconditioning) were found better as expressed by germination and all other vigour parameters compared to matriconditioning with press mud for 24 h or halopriming with 100 mol m-3 CaCl2 for 24 h. Farooq et al. (2006) reported in sunflower for seedling elongation H2O2, NaCl and GA3 were the best while for shoot and root dry weight, salicylic acid and H2O2 were promising. Mavi et al. (2006) found that improvement in plumule length was increased due to earlier germination induced by NaCl priming treatment. Sunflower seeds were hydro primed for 24 h, matriprimed for 24 and 48 h, osmoprimed with 0.5% KNO3 for 12 h and 0.1% NaCl for 12 h. Hydropriming and osmopriming with NaCl resulted in early 50% emergence, increased plant population, achene yield and achene proteins, but plant height and achene oil contents were not affected significantly by different seed priming (Hussain et al., 2006). Kaya et al. (2006) reported that hydroprimed seeds of sunflower and wheat could germinate faster and produced longer seedling under salinity stress compared with untreated seeds. Ramzan et al. (2010) reported that gladiolus seeds primed with distilled water achieved 92% germination followed by 80% in KNO3 1 and 70% in KNO3 2%. It also reduced time required for 50% germination and it increased the seedling length of 14 cm. Mohan Kumar and Manonmani (2011) reported that sunflower hybrid seeds haloprimed with 2% KNO3 for 6 h increase the germination speed, maximize the germination, vigour and field emergence (Table 1).
Effect of seed priming on biochemical parameters: Smith and Comb (1991) found that soluble protein content increased to 109 and 120% in pepper seeds primed in -0.90 and -1.35 MPa NaCl solutions respectively after 12 days of priming and also revealed that there was no significant difference in the soluble protein content between two priming treatments.
| Table 1: | Influence of halopriming on seed germination and seedling vigour of sunflower hybrid (KBSH 44) seeds (Mohan Kumar and Manonmani, 2011) |
 | |
Davison and Bray (1991) reported that the rate of protein synthesis in 4 day germinated unprimed leek seeds is equivalent to 2 day germinated osmoprimed leek seeds for both embryo and endosperm tissue. They also observed that five polypeptides were found to be synthesized in embryonic tissue of leek seeds after 4 days of priming in a -1.0 MPa polyethylene glycol solution which were not present at fourth day of germination of seeds. Khan (1992) found that two amino acids were incorporated in proteins during the first 24 h of imbibitions of sweet pepper seeds in PEG solutions. In contrast, Dell`Aquila and Spada (1992) revealed that the synthesis of a group of proteins associated with radicle emergence was reduced during imbibition of wheat seeds in NaCl. Fujikura and Karssen (1992) found that cauliflower seeds subjected to controlled deterioration, osmopriming and both, the proteins expression correlated with the rate of germination which was reduced in controlled deterioration and enhanced in osmopriming. Chiu et al. (1995) reported that improvement in germination by priming might be due to enhanced repair of membrane due to reduced leakage of electrolytes in primed watermelon seeds than in control. Job et al. (1997) observed that the priming of sugarbeet seeds with PEG 600 has increased the basic â-subunit of 11-S globulin storage protein correspond to 22 kDa polypeptide in primed sugarbeet seeds when compared with the level seen in unprimed seeds. Bailly et al. (1998) reported that priming of aged sunflower seeds in -2 MPa PEG progressively restored the initial germinability and resulted in marked decrease in the level of conjugated dienes indicating a fall in lipid peroxidation processes. Lin and Sung (2001) found that thermo priming at 20°C increases the activities of isocitrate lyase and malate synthase and these increases in the activities of glyoxysome enzymes which were linked to the improved emergence responses in primed bitter gourd seeds. Gonzalez-Zertuche et al. (2001) observed that the priming of Wigandia urens buried seeds showed increased protein concentration and also induced the synthesis of heat stable proteins of 14 and 23 kDa in buried seeds and proteins of high molecular weight of 43 kDa in primed seeds that were not detected in control seeds.
Cruz-Garcia et al. (2003) revealed that changes in sunflower seeds primed with H2O2 diverted a greater part of the cotyledonary resources towards the shoot which was crucial to its earlier establishment and photosynthesis for vigorous growth. Since, this signaling molecule reprogrammed the gene expression leading to de novo protein synthesis, a membrane repair mechanism and other substrates available for improved and synchronized germination. Basra et al. (2003b) reported that wheat seeds were subjected to hydropriming for 6, 12 or 24 h and matriconditioned with gunny bags. The best priming treatment was found to be hydropriming for 24 h which registered maximum emergence percentage and lesser Electrical Conductivity (EC) than control. Naglreiter et al. (2005) observed that priming of Pinus sylvestris seeds with PEG+gibberelic acid (200 mg kg-1 of seeds) showed higher free radical content than in unprimed seeds. Whereas seeds primed with K+ salts observed only minor changes in the free radical levels. However, both priming treatments showed faster germination rates compared to control without changing the germination percent. Wahid et al. (2008) revealed that priming of sunflower seeds with salicylic acid, H2O2 and NaCl induced the de novo synthesis of peptides with low molecular weight of 37 and 57 kDa and high molecular weight of 157 and 167 kDa. Mohan Kumar and Manonmani (2011) reported that sunflower hybrid seeds primed with 2% KNO3 recorded the high α-amylase activity and protease activity during course of germination (Fig. 1 and 2).
Effect of seed priming on molecular parameters: Although priming has been found to improve both the rate and uniformity of germination and emergence in many species, little is known about the biochemical and molecular mechanisms.
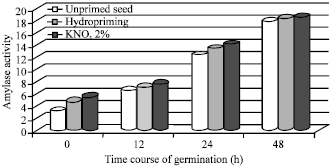 | |
| Fig. 1: | Influence of seed priming on α-amylase activity (mg maltose min-1) of sunflower hybrid (KBSH 44) seeds during course of germination (Mohan Kumar and Manonmani, 2011) |
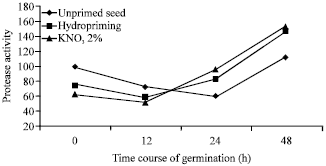 | |
| Fig. 2: | Influence of seed priming on protease activity (OD value) of sunflower hybrid (KBSH 44) seeds during course of germination (Mohan Kumar and Manonmani, 2011) |
A few reported evidence on molecular studies indicated that the increased rate of metabolic process was involved in germination when primed seeds are rehydrated. The beneficial effect of priming has been related to the physiological changes occurring in the partially hydrated embryos and on subsequent germination.
The improved performance of seedling after priming has been explained by the completion of DNA repair mechanisms during the priming period, qualitative and quantitative increase in protein content and rapid declining of reserve materials like phytate and micronutrients during germination of primed seeds (Coolbear and Grierson, 1979; Bray et al., 1989; Job et al., 1997). Dell'Aquila and Taranto (1986) found that the delayed onset of cell division and the low rate of DNA synthesis and its content during osmopriming of wheat seeds were followed by a fast increase of these processes during the early hours of germination in water. They also concluded that in aged seeds, osmoregulation induced more advantageous metabolic changes and enabled this kind of seeds to restore their germinability and vigour more effectively than fresh wheat seeds. Bray et al. (1989) revealed that DNA synthesis was detectable in leek embryos during priming in PEG at -1.0 MPa even in the absence of any cell division and upon subsequent germination there was a five fold increase in the rate of DNA synthesis after a 6 to 12 h lag period of germination when compared with the level seen in unprimed seeds.
Coolbear et al. (1990) found that in tomato, var. Moneymaker, there was a large increase in nucleic acid content especially rRNA, within seeds obtained by low temperature presowing treatment and revealed that this was unlikely to be the immediate cause of more rapid seed germination. In contrast, Clarke and James (1991) reported that there was no net increase in RNA and DNA content of leek seeds during osmopriming. Bino et al. (1992) and Lanteri et al. (1993) studied the effects of priming on nuclear replication activity in tomato and pepper seeds by means of flow cytometry. The amount of 4°C nuclei in tomato seed increased to 60% after 21 days of osmopriming treatment. A positive correlation was found between the induction of DNA synthesis measured as the increase in 4°C cells and the efficiency of osmotic treatments to reduce the mean germination time. In leek seeds, Ashraf and Bray (1993) reported that replicative DNA synthesis and nuclear DNA had occurred during osmopriming and germination of leek seeds. Lanteri et al. (1994) revealed that priming of pepper and tomato seeds in -1.1, -1.3 and -1.5 MPa PEG solution for 14 days at 25°C reduced the mean time to germination. An induction of 4°C signals were also found after priming, indicating that during priming the cells of the embryonic root tip had replicated their DNA and arrested at the G2 phase of the cell cycle. Liu et al. (1996) reported that ‘fresh PEG priming’ of freshly harvested tomato (Lycopersican esculentum Mill) cv. Moneymaker seeds neither alleviated seed dormancy nor promoted DNA replication. However, the addition of 10 μm GA3 to the osmotic priming solution triggered the replicative DNA synthesis of fresh priming seeds and further enhanced the germination process.
Gurusinghe et al. (1999) reported that radicle tip cells of tomato seeds advanced through the cell cycle during hydrothermal priming resulted in an increase in the percentage of nuclei having 4C DNA content in tomato. Gao et al. (2002) reported that germination of Brassica napus was enhanced if the seeds were primed either with water for 12 h or with ABA for 40 h with concomitant reduction in transcript level of BnPER1 transcript (peroxiredoxin-antioxidant protein) when compared to control seeds. Hudson et al. (2007) found that during seed priming one protease gene (At 5 g 67360, a cucumisin-like serine protease) was significantly expressed and two other protease messages (At 5 g 58870, an FtsH protease and At 4 g 39910, an ubiquitin-specific protease) were significantly repressed.
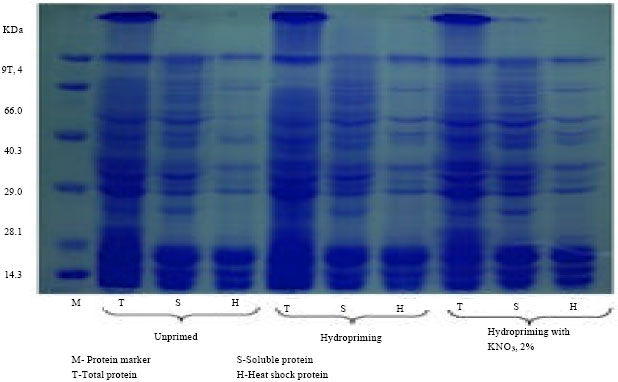 | |
| Fig. 3: | Protein profiles of sunflower hybrid (KBSH 44) seeds (Mohan Kumar and Manonmani, 2011) |
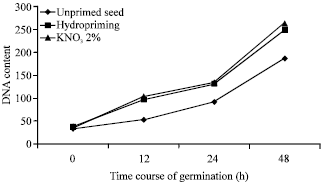 | |
| Fig. 4: | Influence of seed priming on DNA content (μg g-1) of sunflower hybrid (KBSH 44) seeds during course of germination (Mohan Kumar and Manonmani, 2011) |
Mohan Kumar and Manonmani (2011) reported that sunflower hybrid seeds primed with 2% KNO3 recorded the high utilization of proteins and synthesis of new proteins with increased DNA content during course of germination (Fig. 3 and 4).
Effect of seed priming on storability of seeds: Seed storage caused a decrease in the protein content which may be related to oxidation of the amino acids due to the increase in the respiratory activity and advance in the deterioration process of the stored seeds. Thus, prolonged seed storage would increase the metabolic activity of the seeds and consequently decrease the reserve substance content and reduce the dry material weight of the seeds (Bewley and Black, 1982). A number of studies have indicated that relatively short prehydration treatments (either brief imbibition in water or exposure to high relative humidity) can either improve the tolerance of seeds to subsequent adverse storage condition or improve the vigour of aged seeds (Basu and Pal, 1980; Perl, 1979; Burgass and Powell, 1984; Rao et al., 1987). Argerich and Bradford (1989) reported that primed tomato seeds exhibited delayed germination and a lower mean germination when stored at 30°C for 6 months as compared with the control. Tarquis and Bradford (1992) reported that hydropriming of lettuce seeds improved seed germination rate decreased the longevity faster than the nonprimed control seed under controlled deterioration conditions even under mild storage conditions (45°C and 50% relative humidity). Zheng et al. (1994) found that osmopriming of fresh and stored seeds and hydropriming of stored seeds were better than control. Priming induced rapid and uniform germination and rapid emergence of seedlings of canola seeds, which is responsible to repair the age related cellular and subcellular damage of low vigor seeds that may accumulate during seed development (Bray, 1995). Bruggink et al. (1999) reported that the longevity of primed seed was considerably less compared to that of the nonprimed seed lot. Whereas in tomato seeds, a partial restoration of longevity could be obtained by keeping the seeds, after the priming treatment under a mild water and/or temperature stress for a period of several hours to days. Priming temperature influences the success of priming and subsequently affects the longevity of primed seeds (Van Pijlen et al., 1996). The 10°C primed sweetcorn seeds were found to have increased anti-oxidative activity and decreased protein modification and therefore reduced seed deterioration (Murthy and Sun, 2000). Chiu et al. (2002) found that the sweetcorn sh-2 seeds primed at 10°C for 36 h had better storability compared to 20°C-primed seeds when they were stored at 25°C for 12 months. Gurusinghe and Bradford (2001) concluded that osmopriming enhanced the seed germination but longevity of primed seeds in storage was often reduced. However, postpriming heat treatment at 37°C to tomato seeds extended the potential longevity of primed seeds. Basra et al. (2003b) found that canola seeds primed with polyethylene glycol (PEG-10,000) for 4 or 8 h and stored in sealed containers. Among the treatments, osmopriming for 4 h stored seeds resulted more leaf area index, dry matter accumulation, crop growth rate and ultimately higher seed yield than control. Yeh et al. (2005) proved that partial vaccum storage of primed bitter gourd seeds maintain viability upto 12 months. However, primed seeds accumulate more total peroxide than non-primed control after 12 months in non-vaccum storage and this led to a marked decrease in seed longevity. Hill et al. (2007) revealed that primed lettuce seeds stored at 9% moisture content at 38 °C deteriorate faster than primed seeds stored at 6% moisture content at 48°C. Mohan Kumar and Manonmani (2011) reported that haloprimed sunflower seeds with 2% KNO3 maintain the storage potential by recording maximum germination and field emergence after six months of storage than unprimed seeds (Fig. 5-6).
 | |
| Fig. 5: | Effect of seed priming on field emergence potential of sunflower hybrid |
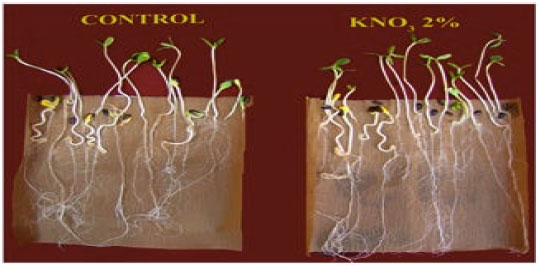 | |
| Fig. 6: | Effect of seed priming in germination and seedling vigour of sunflower hybrid |
CONCLUSION
There is evidence that in most field and horticultural crops have priming led to improvement of germination and seedling establishment. Furthermore, the technique of halopriming having other advantages such as feasibility and low cost. The challenge of technology is to enhance seed performance provides an opportunity for more indepth studies on physiological and biochemical changes that occur during seed treatments. Extending shelf life in storage of primed seeds should be tackled to ensure consistent performance over a wide range of environmental conditions.
REFERENCES
- Afzal, I., S.M.A. Basra, N. Ahmad, M.A. Cheema, E.A. Warraich and A. Khaliq, 2002. Effect of priming and growth regulator treatments on emergence and seedling growth of hybrid maize (Zea mays L.). Int. J. Agric. Biol., 2: 303-306.
Direct Link - Argerich, C.A. and K.J. Bradford, 1989. The effects of priming and ageing on seed vigour in tomato. J. Exp. Bot., 40: 599-607.
Direct Link - Ashraf, M. and C.M. Bray, 1993. DNA synthesis in osmoprimed leek (Allium porrum L.) seeds and evidence for repair and replication. Seed Sci. Res., 3: 15-23.
Direct Link - Bailly, C., A. Benamar, F. Corbineau and D. Come, 1998. Free radical scavenging as affected by accelerated aging and subsequent priming in sunflower seeds. Physiol. Plant., 104: 646-652.
CrossRefDirect Link - Basra, A.S, S. Bedi and C.P. Malik, 1988. Accelerated germination of maize seeds under chilling stress by osmotic priming and associated changes in embryo phospholipids. Ann. Bot., 61: 635-639.
Direct Link - Basra, S.M.A., E. Ullah, E.A. Warraich, M.A. Cheema and I. Afzal, 2003. Effect of storage on growth and yield of primied canola (Brassica napus) seeds. Int. J. Agric. Biol., 5: 117-120.
Direct Link - Basra, S.M.A., I.A. Pannu and I. Afzal, 2003. Evaluation of seedling vigor of hydro and matriprimed wheat (Triticum aestivum L.) Seeds. Int. J. Agric. Biol., 2: 121-123.
Direct Link - Bino, R.J., J.N. De Vries, H.L. Kraak and J.G. Van Pijlen, 1992. Flow cytometric determination of nuclear replication stages in tomato seeds during priming and germination. Ann. Bot., 69: 231-236.
Direct Link - Bradford, K.J., 1986. Manipulation of seed water relations via osmotic priming to improve germination under stress conditions. HortScience, 21: 1105-1112.
Direct Link - Bray, C.M., P.A. Davison, M. Ashraf and R.M. Taylor, 1989. Biochemical changes during osmopriming of leek seeds. Ann. Bot., 63: 185-193.
Direct Link - Bruggink, G.T., J.J.J. Ooms and P. van der Toom, 1999. Induction of longevity in primed seeds. Seed Sci. Res., 9: 49-53.
CrossRefDirect Link - Burgass, R.W. and A.A. Powell, 1984. Evidence for repair process in the invigouration of seeds by hydration. Ann. Bot., 53: 735-757.
Direct Link - Cantliffe, D.J., 1991. Benzyladenine in the priming solution reduces thermodormancy of lettuce seeds. Hortic. Technol., 1: 95-97.
Direct Link - Cantliffe, D.J., J.M. Fischer and T.A. Nell, 1984. Mechanism of seed priming in circumventing thermodormancy in lettuce. Plant Physiol., 75: 290-294.
Direct Link - Chiu, K.Y., C.L. Chen and J.M. Sung, 2002. Effect of priming temperature on storability of primed sh-2 sweet corn seed. Crop Sci., 42: 1996-2003.
Direct Link - Chiu, K.Y., C.S. Wang and J.M. Sung, 1995. Lipid peroxidation and peroxide-scavenging enzymes associated with accelerated aging and hydration of watermelon seeds differing in ploidy. Physiol Plant., 94: 441-446.
CrossRefDirect Link - Clarke, N.A. and P.E. James, 1991. The effects of priming and accelerated ageing upon the nucleic acid content of leek seeds and their embryos. J. Exp. Bot., 42: 261-268.
CrossRefDirect Link - Coolbear, P. and D. Grierson, 1979. Studies on the changes in the major nucleic acid components of tomato seeds (Lycopersicon esculentum Mill.) resulting from osmotic presowing treatments. J. Exp. Bot., 30: 1153-1162.
CrossRef - Coolbear, P., R.J. Slater and J.A. Bryant, 1990. Changes in nucleic acid levels associated with improved germination performance of tomato seeds after low temperature presowing treatment. Ann. Bot., 65: 187-195.
Direct Link - Cruz-Garcia, F., A. Gomez, J.J. Zuniga, J. Plasencia and J.M. Vazquez-Ramos, 2003. Cloning and characterization of a COBRA-like gene expressed de novo during maize germination. Seed Sci. Res., 13: 209-217.
CrossRef - Davison, P.A. and C.M. Bray, 1991. Protein synthesis during osmopriming of leek (Allium porrum L.) seeds. Seed Sci. Res., 1: 29-35.
CrossRef - Dell`Aquila, A. and P. Spada, 1992. Regulation of protein synthesis in germinating wheat embryos under polyethylene glycol and salt stress. Seed Sci. Res., 2: 75-80.
CrossRefDirect Link - Dell'Aquila, A. and G. Taranto, 1986. Cell division and DNA-synthesis during osmopriming treatment and following germination in aged wheat embryos. Seed Sci. Technol., 14: 333-341.
Direct Link - Demer, I. and K. Mavi, 2004. The effect of priming on seedling emergence of differentially matured watermelon (Citrullus lanatus (Thunb.) Matsum and Nakai) seeds. Scient. Hortic., 102: 467-473.
CrossRefDirect Link - Farooq, M., S.M.A. Basra and Hafeez-ur-Rehman, 2006. Seed priming enhances emergence, yield and quality of direct-seeded rice. Int. Rice Res. Notes, 31: 42-44.
Direct Link - Farooq, M., S.M.A. Basra, A. Wahid, N. Ahmad and B.A. Saleem, 2009. Improving the drought tolerance in rice (Oryza sativa L.) by exogenous application of salicylic acid. J. Agron. Crop Sci., 195: 237-246.
CrossRefDirect Link - Fujikura, Y. and C.M. Karssen, 1992. Effects of controlled deterioration and osmopriming on protein synthesis of cauliflower seeds during early germination. Seed Sci. Res., 2: 23-31.
CrossRef - Furutani, S.C., B.H. Zandstra and H.C. Price, 1986. The effect of osmotic solute composition and duration and temperature of priming on onion seed germination. Seed Sci. Technol., 14: 545-551.
Direct Link - Gao, Y.P., P.C. Bonham-Smith and L.V. Gusta, 2002. The role of peroxiredoxin antioxidant and calmodulin in ABA-primed seeds of Brassica napus exposed to abiotic stresses during germination. J. Plant Physiol., 159: 951-958.
Direct Link - Gonzalez-Zertuche, L., C. Vazquez-Yanes, A. Gamboa, M.E. Sanchez-Coronado, P. Aguilera and A. Orozco-Segovia, 2001. Natural priming of Wigandia urens seeds during burial: Effect on germination growth and protein expression. Seed Sci. Res., 11: 27-34.
CrossRefDirect Link - Gurusinghe, S. and K.J. Bradford, 2001. Galactosyl-sucrose oligosaccharides and potential longevity of primed seeds. Seed Sci. Res., 11: 121-134.
Direct Link - Harris, D., A. Joshi, P.A. Khan, P. Gothkar and P.S. Sodhi, 1999. On-farm seed priming in semi-arid agriculture: Development and evaluation in maize, rice and chickpea in India, using participatory methods. Exp. Agric., 35: 15-29.
Direct Link - Hill, H.J., J.D. Cunningham, K.J. Bradford and A.G. Taylor, 2007. Primed lettuce seeds exhibit increased sensitivity to moisture content during controlled deterioration. HortScience, 42: 1436-1439.
Direct Link - Hudson, M.E., T. Bruggink, S.H. Chang, W.J. Yu and B. Han et al., 2007. Analysis of gene expression during Brassica seed germination using a cross-species microarray platform. Crop Sci., 47: 96-112.
Direct Link - Hussain, M., M. Farooq, S.M.A. Basra and N. Ahmad, 2006. Influence of seed priming techniques on the seedling establishment, yield and quality of hybrid sunflower. Int. J. Agric. Biol., 8: 14-18.
Direct Link - Job, C., A. Kersulec, L. Ravasio, S. Chareyre, R. Pepin and D. Job, 1997. The solubilisation of the basic subunit of sugarbeet seed 11-S globulin during priming and early germination. Seed Sci. Res., 7: 225-244.
CrossRef - Kaya, M.D., G. Okcu, M. Atak, Y. Cikili and O. Kolsarici, 2006. Seed treatments to overcome salt and drought stress during germination in sunflower (Helianthus annuus L.). Eur. J. Agron., 24: 291-295.
CrossRefDirect Link - Khan, A.A., 1992. Preplant Physiological Seed Conditioning. In: Horticultural Reviews, Volume 13, Janick, J. (Ed.). John Wiley and Sons, Oxford, UK., pp: 131-181.
CrossRef - Lanteri, S., F. Saracco, H.L. Kraak and R.J. Bino, 1994. The effects of priming on nuclear replication activity and germination of pepper (Capsicum annuum) and tomato (Lycopersicon esculentum) seeds. Seed Sci. Res., 4: 81-87.
CrossRef - Lanteri, S., H.L. Kraak, C.H.R. de Vos and R.J. Bino, 1993. Effects of osmotic pre conditioning on nuclear replication activity in seeds of pepper (Capsicum annum L.). Physiol. Plant., 89: 433-440.
CrossRef - Lin, J.M. and J.M. Sung, 2001. Pre-sowing treatments for improving emergence of bitter gourd seedlings under optimal and sub-optimal temperatures. Seed Sci. Technol., 29: 39-50.
Direct Link - Liu, Y.Q., R.J. Bino, W.J. Van der Burg, S.P.C. Groot and H.W.M. Hilhorst, 1996. Effects of osmotic priming on dormancy and storability of tomato (Lycopersicon esculentum Mill.) seeds. Seed Sci. Res., 6: 49-55.
CrossRef - Mavi, K., S. Ermis and I. Demir, 2006. The effect of priming on tomato rootstock seeds in relation to seedling growth. Asian J. Plant Sci., 5: 940-947.
CrossRefDirect Link - McDonald, M.B., 1999. Seed deterioration: Physiology, repair and assessment. Seed Sci. Technol., 27: 177-237.
Direct Link - Murthy, U.M.N. and W.Q. Sun, 2000. Protein modification by Amadori and Maillard reactions during seed storage: Roles of sugar hydrolysis and lipid peroxidation. J. Exp. Bot., 51: 1221-1228.
Direct Link - Naglreiter, C., T.G. Reichenauer, B.A. Goodman and H.R. Bolhar-Nordonkampf, 2005. Free radical generation in Pinus sylvestris and Larix deciduas seeds primed with polyethylene glycol or potassium salt solution. Plant Physiol. Biotechnol., 43: 117-123.
Direct Link - Perl, M., 1979. Invigoration of cotton seedlings by treatment for pregermination activities. J. Exp. Bot., 114: 183-192.
CrossRefDirect Link - Ramzan, A., I.A. Hafiz, T. Ahmad and N.A. Abbasi, 2010. Effect of priming with potassium nitrate and dehusking on seed germination of Gladiolus (Gladiolus alatus). Pak. J. Bot., 42: 247-258.
Direct Link - Rao, N.K., E.H. Roberts and R.H. Ellis, 1987. The influences of pre-and post-storage hydration treatments on chromosomal aberrations, seedling abnormalities and viability of lettuce seeds. Ann. Bot., 60: 97-108.
Direct Link - Redfearn, M. and D.J. Osborne, 1997. Effects of advancement on nucleic acids in sugarbeet (Beta Vulgaris) seeds. Seed Sci. Res., 7: 261-268.
CrossRef - Smith, P.T. and B.G. Comb, 1991. Physiological and enzymatic activity of pepper seeds (Capsicum annuum) during priming. Physiol. Plant, 82: 433-439.
CrossRef - Subedi, K.D. and B.L. Ma, 2005. Seed priming does not improve corn yield in a humid temperate environment. Agron. J., 97: 211-218.
Direct Link - Tarquis, A.M. and K.J. Bradford, 1992. Prehydration and priming treatments that advance germination also increase the rate of deterioration of lettuce seeds. J. Exp. Bot., 43: 307-317.
Direct Link - Taylor, A.G., P.S. Allen, M.A. Bennett, K.J. Bradford, J.S. Burris and M.K. Misra, 1998. Seed enhancements. Seed Sci. Res., 83: 245-256.
CrossRef - Van Pijlen, J.G., S.P.C. Groot, H.L. Kraak and R.J. Bino, 1996. Effect of pre-storage hydration treatments on germination performance, moisture content, DNA synthesis and controlled deterioration tolerance of tomato (Lycopersicon esculentum Mill) seeds. Seed Sci. Res., 6: 57-63.
CrossRefDirect Link - Wahid, A., A. Noreen, S.M.A. Basra, S. Gelani and M. Farooq, 2008. Priming-induced metabolic changes in sunflower (Helianthus annuus) achenes improve germination and seedling growth. Bot. Stud., 49: 343-350.
Direct Link - Windauer, L., A. Altuna and R. Benech-Arnold, 2007. Hydrotime analysis of Lesquerella fendleri seed germination responses to priming treatments. Ind. Crop Prod., 25: 70-74.
CrossRefDirect Link - Yeh, Y.M., K.Y. Chiu, C.L. Chen and J.M. Sung, 2005. Partial vacuum extends the longevity of primed bitter gourd seeds by enhancing their anti-oxidative activities during storage. Sci. Hortic., 104: 101-112.
CrossRef - Zheng, G.H., R.W. Wilen, A.E. Slinkard and L.V. Gusta, 1994. nhancement of canoloa seed germination and seedling emergence at low temperature by priming. Crop Sci., 34: 1589-1593.
Direct Link