
K.M.G. Gehan Jayasuriya
Department of Botany, University of Peradeniya, Peradeniya, Sri Lanka
Jerry M. Baskin
Department of Biology, University of Kentucky, Lexington, KY, USA
Carol C. Baskin
Department of Biology, University of Kentucky, Lexington, KY, USA
M. Thilina R. Fernando
National Herbarium, Royal Botanic Gardens, Peradeniya, Sri Lanka
Research Journal of Seed Science
Year: 2012 | Volume: 5 | Issue: 1 | Page No.: 1-18







ABSTRACT
Non dormancy, three of the five classes of dormancy and orthodox and recalcitrant storage behavior occur in seeds of Fabaceae. The aim of the study was to characterize whole-seed dormancy and storage behavior in seeds of three tropical species of Derris (Fabaceae), which are lianas. Seed Moisture Content (MC); effects of drying and low temperature on viability; water-uptake of intact and scarified seeds; and effects of scarification, fruit coat removal and GA3 on germination were determined. Seed coat anatomy was studied to check for evidence of physical dormancy. Seeds of D. parvifolia and D. scandens had low MC and those of D. trifoliata high MC. D. trifoliata seeds were sensitive to both drying and low temperature storage. Seeds of D. scandens were water-impermeable and those of D. parvifolia and D. trifoliata water-permeable. D. parvifolia seeds germinated without treatment, whereas those of D. scandens required scarification. Removal of fruit coat and application of GA3 overcame dormancy in D. trifoliata seeds. A palisade layer was present only in the seed coat of D. scandens. D. trifoliata seeds are recalcitrant and those of the other two species orthodox. Seeds of D. parvifolia are nondormant and those of D. scandens and D. trifoliata have Physical (PY) and Physiological (PD) dormancy, respectively. The ecological implications of nondormancy/dormancy in relation to orthodoxy/recalcitrant seed storage behavior in tropical lianas are discussed.
PDF Abstract XML References Citation
Received: September 29, 2011;
Accepted: October 20, 2011;
Published: January 21, 2012
How to cite this article
K.M.G. Gehan Jayasuriya, Jerry M. Baskin, Carol C. Baskin and M. Thilina R. Fernando, 2012. Variation in Seed Dormancy and Storage Behavior of Three Liana Species of Derris (Fabaceae, Faboideae) in Sri Lanka and Ecological Implications. Research Journal of Seed Science, 5: 1-18.
URL: https://scialert.net/abstract/?doi=rjss.2012.1.18
URL: https://scialert.net/abstract/?doi=rjss.2012.1.18
INTRODUCTION
Although much information is available on seed dormancy of Fabaceae species in the temperate zone, relatively few detailed studies have been done on tropical members of the family (Baskin and Baskin, 1998). Seed dormancy in most members of the family in temperate and arctic zones of the world is caused by a water impermeable seed coat (physical dormancy, PY) (e.g., some Medicago spp. and Trifolium spp. (Uzun and Aydin, 2004; Taylor, 2005; Travlos and Economou, 2006; Balouchi and Sanavy, 2006) and in a few taxa by PY in combination with a Physiologically Dormant (PD) embryo, i.e., (PY+PD), e.g., Medicago radiata (Balouchi and Sanavy, 2006). In tropical and subtropical zones, on the other hand, seeds of Fabaceae species have PY e.g., Sesbania rostrata (Sarker et al., 2000), Crotolaria retusa (Alderete-Chavez et al., 2010) and Bauhinia spp. (Alderete-Chavez et al., 2010; Asiedu et al., 2011), PD, e.g., Humboldtia laurifolia (Jayasuriya et al., 2010) or (rarely) (PY+PD), or they are Non-Dormant (ND) (Baskin and Baskin, 1998; Sautu et al., 2007).
With regard to storage behavior, seeds of Fabaceae species are orthodox (desiccation-tolerant), intermediate or recalcitrant (desiccation-intolerant) (Dickie and Pritchard, 2002). Although the majority of Fabaceae species thus far investigated produce orthodox seeds, the storage behavior of only a relatively few tropical members of this family has been determined (Dickie and Pritchard, 2002). Thus, neither seed storage behavior nor seed dormancy has been studied well in tropical Fabaceae.
To add to the knowledge of seed dormancy of tropical Fabaceae species, dormancy and storage behavior of seeds of this family in Sri Lanka were studied. To our surprise, initial observations and experiments suggested that seeds of three Derris species differed in kind of dormancy and in storage behavior. Thus, special attention was given to seed dormancy and storage behavior of seeds of these three Derris species. There are no previous reports of PY, PD and ND and of both orthodox and recalcitrant seeds in any genus of the 17 families known to have water-impermeable seeds.
Within the 17 angiosperm families containing species that produce seeds with PY, only a few records in the literature are available on genera with species that produce seeds with PY as well as species that produce seeds with other kinds of dormancy. Meisert (2002) found that seeds of some Erodium and Pelargonium species have PY, while seeds of a few Erodium and Pelargonium species have no dormancy. Further, Meisert et al. (1999) showed that seeds of nondormant Erodium and Pelargonium species have a palisade layer in the seed coat and that water enters the seed through an opening analogous to the water gap in seeds with PY (Gama-Arachchige et al., 2010, 2011). Observations of Jayasuriya et al. (2008) on Bonamia are in agreement with Meisert (2002). Jayasuriya et al. (2008) showed that seeds of B. grandiflora have PY, while those of B. menziesii have no dormancy. The seed coat of B. menziesii consists of palisade cells, but seeds imbibe water through a permanently-open hilar fissure. However, genera consisting of species that produce seeds with nondormancy and PD, e.g., Planchonella spp. (Ng, 1978), Santiria spp. (Ng, 1973, 1978), Scordocarpus spp. (Ng, 1980; Gilbert, 1952) and Teijsmanniodendron spp. (Ng, 1978) or species that produce seeds with Morphological Dormancy (MD) and Morphophysiological Dormancy (MPD), e.g., Euterpe spp. (Mullett et al., 1981; Bannister, 1970), Aristolochia spp. (Adams et al., 2005) do occur among seed plants. However, in general species in the same genus have the same dormancy class (Baskin and Baskin, 1998).
Seed storage behavior is also consistent within a genus with few exceptions. Seeds of Coffea species have a continuum of seed storage behaviour that ranges from recalcitrant to intermediate and then to orthodox (Eira et al., 2006). Acer saccharum (Jones, 1920) and A. opalus (Gleiser et al., 2004) produce orthodox seeds, while A. saccharinum (Jones, 1920) produce recalcitrant seeds (Greggains et al., 2000).
A single genus containing species that produce seeds with different kinds of dormancy or storage behaviors is important from several aspects of seed biology. In particular, these species provide good opportunities to study the evolution of seed dormancy, the ecological significance of seed dormancy and the comparative biochemistry and molecular biology of different kinds of dormancy.
The main objective of this research was to characterize dormancy and storage behavior of seeds of the three tropical Derris species, i.e., D. trifoliata, D. scandens and D. parvifolia and to discuss the significance of the kinds of seed dormancy and of storage behavior of the three Derris species in relation to their ecology.
MATERIALS AND METHODS
Study organisms: Our study organisms belong to the genus Derris (Fabaceae, subfamily Faboideae, tribe Dalbergieae). There are about 50 species of Derris, with most of them distributed from south Asia to northern Australia; the distribution of D. trifoliata extends to east Africa and to the western Pacific (Mabberley, 1997, 2008). Three of the five native Derris species in Sri Lanka were included in this study and all three are woody lianas. Derris parvifolia is endemic to Sri Lanka and occurs in dry mixed semi-evergreen forests in the dry zone. Derris trifoliata occurs in mangrove swamps in both the dry and wet zones and D. scandens in mangrove swamps as well as in other saline or fresh water marshes in the dry zone (Rudd, 1991).
Collection and description of fruits and seeds: Fruits/seeds of Derris trifoliata and D. scandens were collected from numerous plants at different locations in the wet zone (Matara, Thangalla, Galle and Ambalangoda) and in the dry zone (Hambanthota), while fruits/seeds of D. parvifolia were collected from numerous plants in Polonnaruwa and Naula located in the dry zone of Sri Lanka. Derris scandens and D. parvifolia fruits/seeds were collected in December 2008 and September 2009 and those of D. trifoliata in May 2009 and May 2010. Fruits/seeds were collected from lianas of the three species by shaking the fruiting branches gently. Only brown, mature fruits were collected. During maturation drying on parent plants, fruit color changes from green to yellow to brown and fruits are dispersed as soon as they become brown (Jayasuriya K.M.G.G., personal observation). Fruits/seeds were placed in polythene bags and transported to the Department of Botany, University of Peradeniya, Peradeniya, Sri Lanka and experiments were initiated within 2 weeks after collection.
The fruit of Derris trifoliata is a one-seeded (rarely two-seeded) flattened legume with a mass (Mean±SD) of 148±14 mg. Fruits are oval-shaped and do not have any special appendages to support dispersal; however, they are buoyant in water. Seeds of this species have a very thin seed coat and the whole fruit acts as the dispersal as well as the germination unit. Fruits of D. scandens and D. parvifolia are three-seeded (rarely two-seeded) flattened legumes with a mass of 157±11 and 136±8 mg, respectively. They are linear in shape and resemble wings. The fruit coat of D. parvifolia is papery, which aids the fruit in wind dispersal. The whole fruit is the dispersal and germination unit for D. parvifolia. The fruit coat of D. scandens is thicker than that of D. parvifolia, but it can aid in short distance wind dispersal. The fruit is buoyant; therefore, it also can float and thus be dispersed by water. The fruit is the dispersal unit of D. scandens. However, the fruit coat splits during dispersal, releasing the seeds, which are the germination unit of this species. Seeds of D. scandens and D. parvifolia resemble a “typical” bean seed with a clear hilum and a lens. Derris trifoliata seeds are also bean-shaped, but they do not have a clear hilum or a lens. Seed mass (Mean±SD) of D. scandens, D. parvifolia and D. trifoliata is 47±7, 42±6 and 103±17 mg, respectively.
Moisture content and imbibition of seeds: If seeds are recalcitrant, their initial MC probably is >15%, whereas if MC is <15% there is a good possibility that they are orthodox. Fifteen fresh seeds of each species were weighed individually to the nearest 0.0001 g using a digital analytical balance and then dried separately in an oven at 110°C. Seeds were retrieved from the oven and weighed after 3 h and then at 1 h intervals until they reached a constant mass, which was at 22, 24 and 28 h for D. trifoliata, D. parvifolia and D. scandens, respectively. Seeds of D. scandens were scarified prior to drying. Seed Moisture Content (%MC) was calculated as [(fresh seed mass-oven-dry seed mass)/fresh seed mass]x100 (Hong and Ellis, 1996).
If seeds or fruits have a water-impermeable coat, they will not imbibe, whereas manually-scarified seeds and fruits will do so. Fifteen nontreated (intact) and 15 manually scarified (individually with a razor blade) seeds and fruits (with seeds inside) of D. parvifolia and D. trifoliata and of only seeds (germination unit) of D. scandens were weighed individually to the nearest 0.0001 g with a digital analytical balance. These germination units were placed on moistened filter paper (Whatman No. 1) in separate 9-cm-diameter Petri dishes, retrieved at the time intervals shown in Fig. 1, blotted with filter paper, reweighed and returned to the Petri dish for a total of 30 days or until all of them were germinated. Percentage water uptake (%Ws) was calculated as [(Wi-Wd)/Wd]x100, where Ws = increase in seed mass, Wi = seed mass after water uptake (imbibition) for a given period of time and Wd = initial seed mass.
Effect of drying or of storage at low temperatures on germination of D. trifoliata seeds: If seeds are recalcitrant, they probably will lose viability when dried to a MC <15 % and also when stored at -1 or 5°C. Three samples consisting of three replicates of 15 seeds each of D. trifoliata were air-dried at ambient laboratory conditions to 30, 20 and 10% MC. Then, they were incubated at ambient laboratory conditions on filter papers moistened with a 100 ppm gibberellic acid solution to help ensure that seeds would germinate if they were viable. Seeds were checked for germination at 2-day intervals for 46 days. Four samples consisting of three replicates of 15 seeds each of D. trifoliata were stored at -1 and 5°C for 1 or 2 months and checked for germination as described above. Seeds were kept in sealed ziplock plastic bags during storage to minimize water loss. Radicle protrusion was the criterion for germination.
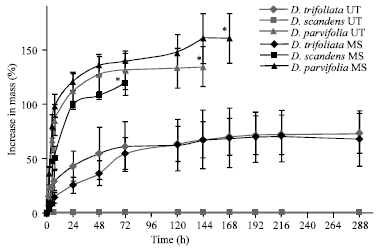 | |
| Fig. 1: | Imbibition of intact and manually scarified seeds of Derris trifoliata, D. scandens and D. parvifolia collected in 2009. Data are shown only for a maximum of 12 days although the experiment was conducted for 30 days. However, no significant increase or decrease in mass was observed after the last data points shown on the graph. *Imbibition test was terminated because all the tested seeds germinated. Error bars are±1 SD. UT, untreated; MS, manually scarified |
Effect of fruit coat on rate of drying of D. trifoliata seeds: Seeds were collected in 2009. Each of 14 samples containing 15 fruits was weighed individually and 12 of them were placed on dry filter paper in Petri dishes and allowed to air-dry under laboratory conditions. Two samples each were retrieved after 1, 2, 3, 4, 6 and 8 weeks. For time 0 fruits and for those retrieved after various drying intervals, fruit coats were removed and the seeds weighed. Then, the seeds were oven-dried to a constant mass at 110°C. In the trial for seeds with fruit coats removed, 15 D. trifoliata seeds were weighed individually at time 0 and after 1, 2, 3, 4 and 6 weeks of drying on filter paper in open Petri dishes. After 6 wk, seeds were dried to constant mass at 110°C. Seed moisture content (%MC) for a given period of drying was calculated as:
Germination of fresh seeds: If seeds are dormant, fresh intact seeds will not germinate. Three replicates of 15 nontreated (intact-fresh) 2010-collected seeds of D. trifoliata and 2009-collected seeds of the other two species, intact fruits (with seeds inside) of D. trifoliata and D. parviflora and manually-scarified (individually with a razor blade) seeds of D. parviflora and D. scandens were placed on moistened filter paper in 9-cm-diameter Petri dishes and incubated at ambient laboratory temperature (c. 25°C) under artificial room fluorescent light + diffuse sunlight for about 10 h per day or in continuous darkness. Darkness was provided by wrapping Petri dishes with aluminum foil. Germination experiments for D. trifoliata were conducted in 2010, while experiments on the other species were conducted in 2009. Seeds incubated in room-light conditions were checked for germination at 2-day intervals until all of them germinated, while those incubated in darkness were checked only after 30 days, at the end of the experiment. Radicle protrusion was the criterion for germination. For D. parvifolia, the fruit was counted as germinated only when radicles had emerged from all of the three seeds within it. However, when one seed in a D. parvifolia fruit germinated the others germinated within 1-2 days. Fruits of D. trifoliata have only one seed.
Initial site of water entry into seeds as shown by dye tracking: Fifteen untreated seeds each of D. parvifolia, D. trifoliata and D. scandens and of 15 boiled seeds (made nondormant by boiling them in water for 30 sec) of D. scandens were immersed in saturated methylene blue solution. Three seeds of each species were retrieved after 15 and 30 min and 1, 2 and 3 h. Transverse cuts by hand were made and observed under a dissecting microscope. Photographs were taken using a Leica L2 stereomicroscope camera and used to determine the initial site of water entry into the seeds.
Fruit and seed coat anatomy: Hand sections of seeds of the three Derris species at the hilum/lens area and at a position on the seed coat away from this area (regular seed coat) were made using a razor blade. Sections of the fruit coat of D. parvifolia and D. trifoliata also were made as described above, since the fruit is the germination unit of these two species. Sections were observed under a light microscope and drawings prepared. Photomicrographs were taken using an Olympus CX21 light microscope connected to an Olympus DP 20 SE camera.
Effect of gibberellic acid and removal of fruit coat on seed germination of D. trifoliata: Using seeds and fruits collected in 2010, three replicates of 15 fruits and of 15 intact seeds freed from the fruit coat were placed on filter paper moistened with 0 (distilled water control), 100 or 500 ppm GA3 in 9-cm-diameter Petri dishes. Seeds and fruits were incubated at laboratory temperature and light/dark conditions (see above) and checked for germination at 2-day intervals until all of them had germinated. Radicle protrusion was the criterion for germination.
Effect of dry vs. wet storage and of immersion in water on germination of D. scandens seeds: A water-immersion experiment was performed because some seeds of this species are dispersed by water and thus immersion in water may have an effect on dormancy break during the dispersal period. To test the effect of dry storage, three replicates each containing 25 D. scandens seeds (germination unit) collected in 2009 were placed on dry filter paper in open 9-cm-diameter Petri dishes and stored at ambient laboratory temperature for 2, 4 or 6 months. Then, they were transferred to moistened filter paper and incubated at ambient laboratory temperatures in light/dark. Seeds were checked for germination at 2-day intervals for 30 days. The effect of wet storage was determined by placing three replicates of 25 seeds each on moistened filter paper in 9-cm-diameter Petri dishes and monitoring germination at 5-day intervals for 180 days. To test the effect of immersion, three replicates of 100 seeds were immersed in distilled water at ambient laboratory conditions and checked for germination at 5-day intervals for 180 days. Radicle protrusion was the criterion for germination.
Analysis of data: All experiments were carried out in a completely randomized design. Pooled t-tests were done to determine differences in imbibition data between intact and manually scarified seeds. One way ANOVA was conducted to determine significant differences between untreated seeds, manually scarified seeds and intact fruits during the imbibition test. Regression analysis was performed to analyze the trend between moisture content and germination of D. trifoliata seeds. Cumulative germination progress curves for D. trifoliata seeds were modeled by the Weibull distribution function and germination curves were used to determine time taken for 50% germination (T50) for each treatment. One-way ANOVA was used to analyze data on effect of drying and storage at low temperatures and on the effect of GA3 and other manipulations of the seeds and fruits of D. trifoliata on germination. A two-way ANOVA was used to analyze data collected from the germination experiment of the three species and the experiment on dormancy break of D. scandens seeds. In the analysis of the data from the germination experiment of the three species, treatment (untreated seeds, untreated fruits and manually-scarified seeds), light condition (light/dark or dark) and species were used as factors. In the analysis of dormancy breaking of D. scandens seeds, treatments (dry storage, wet storage, immersion in water) and storage time were the two factors. Pooled t-tests were carried out to determine the differences between moisture content of seeds stored with and without fruit coat for the same period of time. All germination data were arc-sine square root transformed prior to analysis. Duncan’s multiple mean separation procedure was used to separate means. SAS statistical software was used to analyze the data.
RESULTS
Moisture content and imbibition of seeds: Freshly matured seeds of D. parvifolia and D. scandens had a MC of 10.1 and 10.4%, respectively, while the MC of D. trifoliata seeds was 57.1%. All untreated and manually scarified seeds of D. parvifolia germinated within 7 and 6 days, respectively. Increase in mass of both untreated and manually scarified seeds of D. parvifolia during this period was >120 % (Fig. 1), whereas increase in mass of intact fruits of this species was >200 % (data not shown). There was no significant difference in mass increase between untreated and manually scarified seeds of D. parvifolia (t = 1.35, df = 14, p = 0.104), but mass increase of the fruits differed significantly from that of the other two species (F = 3.34, p = 0.048). However, this was due to the absorption of water by the fruit wall. None of the D. trifoliata untreated or manually scarified seeds germinated within 30 days. However, during this imbibition period mass of both untreated and manually-scarified D. trifoliata seeds increased about 70%, whereas that of intact fruits increased >200% (data not shown) (F = 38.45, p<0.001). In contrast to the other two species, mass of manually-scarified D. scandens seeds increased >115%, while that of intact untreated seeds increased <0.5% (t = -21.12, df = 14, p<0.001) (Fig. 1). All manually scarified D. scandens seeds germinated within 3 days, while none of the untreated D. scandens seeds germinated during the 30 day imbibition period.
Effect of drying or of storage at low temperatures on germination of D. trifoliata seeds: Seeds dried to 30, 20 and 10% MC germinated to 56.7, 40 and 6.7%, respectively. All nongerminated seeds died and rotted within 3-5 days. There was a significant 3rd degree polynomial relationship (R2 = 0.98) between seed MC vs. germination (= viability in this situation) (Fig. 2). The predicted equation suggests that all of the seeds die at 8% MC. None of the D. trifoliata seeds germinated after 1 or 2 months of storage at either -1 or 5°C.
Effect of fruit coat on rate of drying of D. trifoliata seeds: Derris trifoliata seeds without the fruit coat lost moisture more rapidly than those with a fruit coat (Fig. 3). Seeds with and without a fruit coat reached about 15% MC between 5 and 6 weeks and between 2 and 3 weeks, respectively (arrows in Fig. 3). After 6 weeks, MC of seeds without a fruit coat was 9%, while even after 8 weeks the MC of seeds with a fruit coat had not reached 10% (Fig. 3) (t = 2.71, df = 14, p = 0.024).
Germination of fresh seeds: After 30 days, 92% and 19% of the D. trifoliata seeds with and without fruit coat removed, respectively, had germinated and 100 and 26% of scarified and non-scarified (nontreated) seeds of D. scandens, respectively, had germinated (Fig. 4).
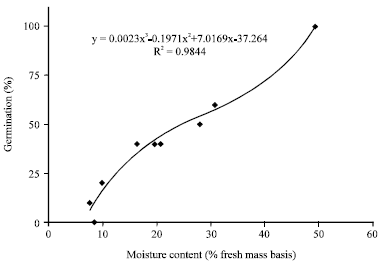 | |
| Fig. 2: | Germination of 2009-collected seeds of Derris trifoliata dried to different seed moisture levels. Line represents the 3rd degree polynomial regression for the data shown in solid diamonds |
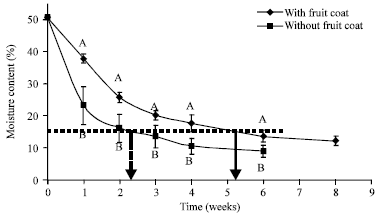 | |
| Fig. 3: | Moisture loss of Derris trifoliata seeds with and without fruit coat. Different uppercase letters indicate significant differences between seeds with and without fruit coat for the same period of drying. Error bars are ±1 SD. Arrows depict time taken for seeds with and seeds without fruit coat to reach 15% MC |
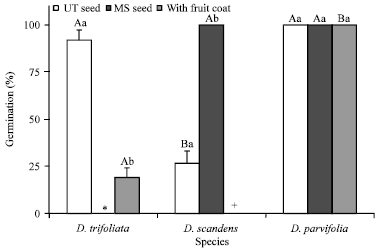 | |
| Fig. 4: | Germination of 2010-collected intact fruits and fresh untreated and manually scarified seeds of Derris trifoliata, D. scandens and D. parvifolia under ambient laboratory temperature and light conditions. Different uppercase letters indicate significant differences between species within the same treatment and different lowercase letters significant differences between treatments within the same species. Error bars are +1 SD. UT, untreated; MS, manually scarified; * treatment not conducted for D. trifoliata; +treatment not conducted for D. scandens |
For D. parvifolia, 100% of nontreated and manually-scarified individual seeds and all seeds inside the fruits germinated within 30 days. There were no significant differences between seeds germinated in light/dark versus dark conditions for any of the three species (data not shown).
Initial site of water entry into seeds as shown by dye tracking: No staining was observed in nontreated (lens closed) D. scandens seeds throughout the 3 h period of the dye-tracking experiment (Fig. 5a). After 15 min, blue-stained tissue was observed in boiled (for 30 sec., to open the lens) seeds of D. scandens (Fig. 5b) and in nontreated seeds of D. parvifolia (not shown) and D. trifoliata (Fig. 5d).
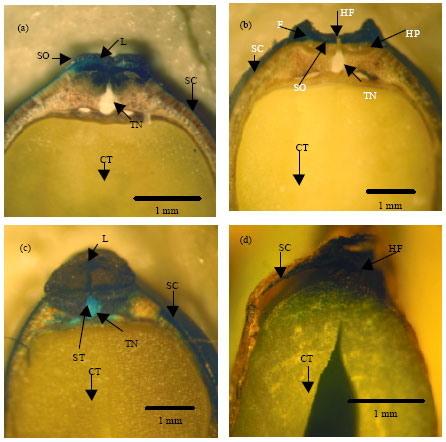 | |
| Fig. 5(a-d): | Uptake of dye (indicated by blue color) after 15 min by untreated (a) and treated (boiled for 30 sec) (b, c) seeds of Derris scandens and untreated seeds (d) of D. trifoliata. CT: Cotyledon; F: Remnants of the funiculus; HF: Hilar fissure; HP: Hilar pad; L: Lens; SC: Seed coat; SO: Staining on outside of seed coat; ST: Staining on inside of seed coat; TN: Tracheid nest |
Stain first appeared in the tissues below the hilum scar in seeds of D. parvifolia and D. trifoliata (Fig. 5d). In treated seeds of D. scandens (Fig. 5b, c), stain was observed first in the tissues below the lens. After 30 min, stain was observed in the tissues below the regular seed coat away from the hilum/lens region in seeds of D. trifoliata and D. parvifolia (not shown). However, even after 2 h no stain was observed in the tissues below the seed coat away from hilar region in boiled D. scandens seeds; instead, staining spread to the tissues below the hilar scar. Even after 3 h, no stain was observed in non-treated D. scandens seeds.
Fruit and seed coat anatomy: Anatomy of the fruit coat of D. parvifolia (Fig. 6a) and D. trifoliata (Fig. 6b) differed. Three main layers could be seen in the fruit coat of D. parvifolia (Fig. 6a). The inner and outer layers consist of parenchyma cells with brown depositions, while the middle layer consists of a tissue containing fibers without any pigment depositions. The outermost layer of the fruit coat of D. trifoliata has a distinct epidermal layer (Fig. 6b). Below this epidermal layer, there are 5-6 layers of tissue consisting of loosely packed parenchyma cells. The innermost layer consists of a separate layer of fibers that easily detaches from the other parts of the fruit coat. No palisade layer of cells was observed in the fruit coat of either D. parvifolia or D. trifoliata.
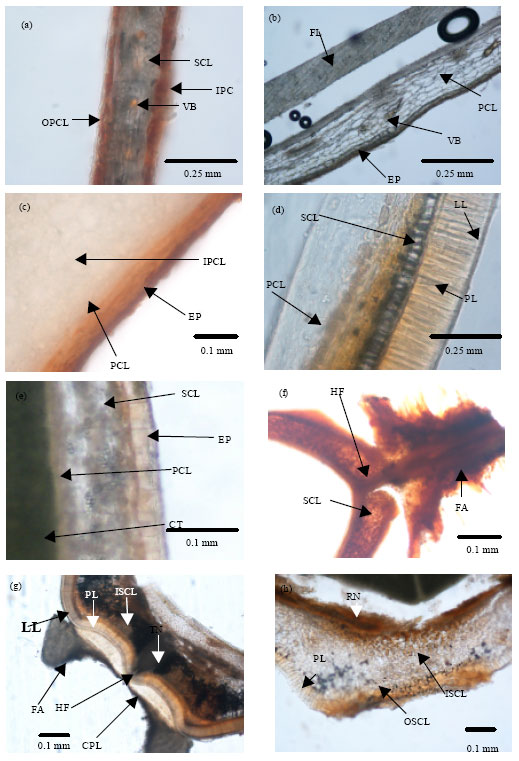 | |
| Fig. 6 (a-h): | Cross sections through fruit coat of Derris parvifolia (a) and D. trifoliata (b); of seed coat away from the hilum area of seeds of Derris parvifolia (c), D. scandens (d) and D. trifoliata (e); and of seed coat in the hilum area of seeds of Derris parvifolia (f), D. scandens (g) and D. trifoliata (h). CT: Cotyledons; CPL: Counter palisade cell layer; EP: Epidermis; FA: Remnants of funiculus; FL: Fiber layer; HF: Hilar fissure; IPCL: Inner parenchyma layer; ISCL: Inner sclereid cell layer; LL: Light line; OPCL: Outer parenchyma layer; OSCL: Outer sclereid layer; PCL: Parenchyma layer; PL: Palisade layer; RN: Remnants of the nucellus; SCL: Sclereid layer; TN: Tracheid nest and VB: Vascular bundle |
Anatomy of the seed coat away from hilum: Anatomy of the seed coat away from the hilum differs between the three species. In D. parvifolia, it consists of two layers and the innermost one is the epidermis, which consists of 3-4 layers of flattened cells with brown-colored (presumably) phenolic depositions (Fig. 6c). Beneath this layer, there are 8-9 layers of parenchyma cells, which may be remnants of the nucellus, endosperm and inner integument. No palisade cell layer was observed in the seed coat of D. parvifolia away from the hilum (Fig. 6c).
The seed coat anatomy of D. scandens is typical of that of physically dormant seeds (Fig. 6d). Thus, the outermost layer of the seed coat is a distinct palisade layer. There is a clear light line in the palisade layer, which is also typical for seed coats of physically dormant seeds. Beneath the palisade layer, there is a sclereid layer that appears as a dark-colored region with square-shaped cells (Fig. 6d). The innermost layer consists of parenchyma cells that may be remnants of the nucellus, endosperm and inner integument. Brown-colored depositions also could be seen in these cells.
The seed coat of D. trifoliata away from the hilum consists of three cell layers (Fig. 6e). The outermost layer is the epidermis, which is made up of rectangular-shaped cells with brown-colored depositions. Beneath the epidermis, there are 6-8 sclereid cell layers with spherical to irregular shaped cells. The innermost layer consists of parenchyma cells, which may be a mixture of remnants of the nucellus, endosperm and inner integument (Fig. 6e). No palisade cell layer was observed in the seed coat of D. trifoliata away from the hilum.
Anatomy of the hilum area: The anatomy of the seed coat is not the same throughout its entirety. Thus, the anatomy of the hilum area of the seeds of all three species differs from that of the seed coat away from the hilum. In the hilum area of the seed coat of D. parvifolia, remnants of the funiculus could be seen clearly (Fig. 6f). There is no considerable anatomical change near the hilum region and no palisade cells are present near the hilum (Fig. 6f). In D. scandens, several changes could be seen in seed coat anatomy near the hilum region compared to the seed coat away from the hilum (Fig. 6g). The palisade layer in the seed coat away from the hilum continues in the hilum region. In addition to this palisade layer, a counter palisade layer occurs above the palisade layer in the hilum pad; there is no light line in the counter palisade layer (Fig. 6g). A clear hilar fissure could be seen as a suture in the middle of the hilum pad. A tracheid nest is located below the hilum fissure (Fig. 6g). Moreover, the innermost parenchyma in the seed coat away from the hilum has been replaced by a sclereid layer in the hilum region. In the seed coat of D. trifoliata, the outermost layer of rectangular cells changes to a palisade cell layer near the hilum region (Fig. 6h). However, it gradually disappears at the hilum scar. The hilum scar region is filled with sclereids and parenchyma cells. Thus, in the seed coat of D. trifoliata palisade cells are present in the hilum region (Fig. 6h).
Effect of gibberellic acid and removal of fruit coat on germination of D. trifoliata: Only 19% of the seeds in intact (nontreated) D. trifoliata fruits collected in 2010 had germinated after 30 days, whereas 91% of those with the fruit coat removed germinated within 30 days (Fig. 7). Seeds of D. trifoliata with fruit coat intact took 71 days to reach 50 % germination, whereas the T50 value of seeds with fruit coat removed was 8.5 days (Table 1). Seeds of D. trifoliata with fruit coat intact collected in 2010 treated with 100 ppm and 500 ppm GA3 germinated to 55 and 70%, respectively, within 30 days. T50 values of seeds in fruits treated with 500 ppm GA3 were significantly lower (i.e., germination rate faster) (F = 206.8, p<0.001) than those treated with 100 ppm (Table 1).
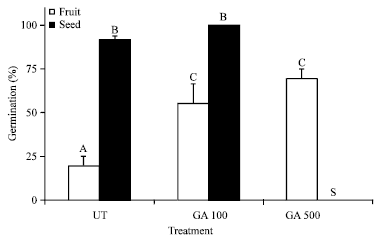 | |
| Fig. 7: | Effect of GA3 on germination of D. trifoliata (2010-collected) intact seeds (seed) and seeds within fruits (fruit). Different uppercase letters indicate significant differences between treatments. Error bars are +1 SD. S, Fruit coat removal treatment was not conducted for 500 ppm gibberellic acid treatment |
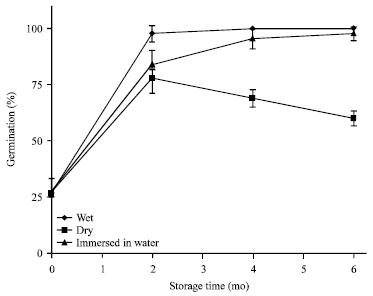 | |
| Fig. 8: | Effect of storage treatments on germination of 2009-collected seeds of D. scandens. Error bars are ±1 SD |
Seeds with fruit coat removed treated with 100 ppm GA3 germinated to 100% within 30 days compared to 91% for nontreated seeds (Fig. 7). Although, GA3-treated seeds germinated faster (i.e., T50 was lower) than nontreated ones, the difference was not significant (Table 1).
Effect of wet or dry storage and of immersion in water on germination of D. scandens: Fresh nontreated D. scandens seeds germinated to 27%, whereas those stored dry for 2 months germinated to 78 % (Fig. 8). Seeds of D. scandens stored wet for 2 and 4 months germinated to 98 and 100%, respectively. Seeds immersed in water for 2 and 4 months germinated to 84 and 95%, respectively (Fig. 8).
| Table 1: | T50 values calculated for germination of D. trifoliata seeds in different treatments. Different lowercase letters indicate significant difference between treatments. |
 | |
| Different lowercase letters depict significant statistical differences between treatments | |
DISCUSSION
Seeds of D. parvifolia and D. scandens had a low (c. 10% fresh mass basis) initial seed MC, whereas those of D. trifoliata had a high (c. 57%) MC. These data suggested that seeds of D. parvifolia and D. scandens were orthodox and that those of D. trifoliata were recalcitrant. Recalcitrancy for seeds of D. trifoliata was confirmed by results of the seed drying experiments, which showed that they could not tolerate drying. Further, none of the D. trifoliata seeds germinated even after only 1 month of storage at -1 or at 5°C. Since seeds were in sealed ziplock bags, loss of viability during low temperature storage could not be due to water loss. Although seeds of several other species of Fabaceae have been shown to be recalcitrant, only a very small proportion of species in this large family produce desiccation-intolerant seeds (Dickie and Pritchard, 2002). These authors reported that 1.2% of Fabaceae species studied were recalcitrant and 0.2% intermediate in storage behavior. Thus, the seeds of >98% of Fabaceae species are orthodox. Recently, two species of Humboldtia (Fabaceae) have been shown to produce recalcitrant seeds (Saba et al., 2008; Jayasuriya et al., 2010).
There are only a few reports in the literature of both recalcitrant and orthodox seeds within the same genus. Storage behavior of Coffea species varies from orthodox to recalcitrant to intermediate (Eira et al., 2006). Dickie and Pritchard (2002) analyzed the Kew Millennium Seed Bank database and reported that both orthodox and recalcitrant storage behavior occur in seeds of Acer, Agathis, Araucaria, Calophyllum, Castanopsis, Citrus, Coprosma, Diospyros, Garcinia, Magnolia, Pittosporum, Spondias and Vitex; none of these is a legume. Apparently, then, our study is the first one to report both orthodox and recalcitrant seeds in a genus of the Fabaceae. Further, since none of the genera listed above by Dickie and Pritchard (2002) is in a family known to have PY (Baskin and Baskin, 2004b), our study also is the first one to report both orthodox and recalcitrant seed behavior in a genus in a family that contains species with PY.
Available information suggests that seeds of most tropical Fabaceae species have PY. However, some species produce seeds with PD (e.g., Prionia copaifera, Tachigalia versicolor, Andira inermis and Dipteryx oleifera (Sautu et al., 2007) and seeds of Humboldtia laurifolia even have physiological epicotyl dormancy (Jayasuriya et al., 2010). Other species produce seeds with no dormancy, e.g., Pongamia pinnata (Ramdeo, 1970; Kumar et al., 2007), Millettia ferruguinea (Teketay, 1998), Inga spp. and Albizia adinocephala (Sautu et al., 2007).
Intact seeds of D. parvifolia and D. trifoliata imbibed water equally well with or without the fruit coat intact. Thus, seeds of both species have a water permeable seed and fruit coat and thus do not have PY. Both intact and manually scarified D. parvifolia seeds and fruits (germination unit) germinated equally well in light/dark and in dark, indicating that the seeds are nondormant. In physically dormant seeds, the palisade layer in the seed coat is responsible for water impermeability (Baskin et al., 2000). Neither seeds of D. parvifolia nor D. trifoliata have a palisade layer(s) in their seed coat. Also, dye-tracking experiments showed that water enters the seed throughout the entire seed coat of these two species, although high amounts of water initially entered through the hilar fissure. On the other hand, fresh intact seeds of D. scandens did not take up water at all, whereas manually scarified seeds imbibed well, indicating the presence of a water impermeable seed coat, thus PY.
Fresh mechanically scarified D. scandens seeds germinated well, whereas only a few intact seeds imbibed water and germinated during a 30-day incubation period. These results confirm that seeds of this species have a water-impermeable seed coat and that embryos are not physiologically dormant. Thus, the seeds have PY and not combinational dormancy (PY+PD). Dye tracking experiments showed that intact D. scandens seeds could not absorb the dye. However, when seeds of this species were boiled (30 sec) they imbibed the dye (only) through the lens, showing that the lens is the water gap in seeds of D. scandens, as it is in other hard seeded faboid legumes (Baskin, 2003). When intact seeds of D. scandens were stored dry or wet for 2 months, some of them came out of dormancy and germinated. However, when seeds were stored dry for 4 or 6 months, germination percentage decreased, whereas wet storage for 4 or 6 months increased the germination percentage. These 4 and 6 month periods of dry storage did not cause the seeds to lose viability; nongerminated seeds germinated after they were scarified (data not shown). Dry storage may have reduced the sensitivity of seeds to dormancy breaking treatment (Jayasuriya et al., 2009). In which case, D. scandens seeds may be capable of sensitivity cycling.
Seeds of D. trifoliata with the fruit coat removed germinated to 91% within 30 days, whereas those with the fruit coat intact germinated to only 19%. The dispersal and germination unit of D. trifoliata is the fruit. Thus, the dispersal unit of this species has PD. That is, the embryo in fresh seeds does not have enough growth potential to overcome the mechanical resistance of the fruit coat (Baskin and Baskin, 1998, 2004a). With time (warm stratification), however, dormancy is lost in the germination unit. Seeds with fruit coat intact reached 100% germination in 14 weeks. Further, seeds with fruit coat intact treated with 100 or 500 ppm GA3 germinated to 100% within 30 days. These results can be interpreted to mean that time and GA3 overcame dormancy in D. trifoliata via an increase in growth potential of the embryo. Embryos of D. trifoliata germinated and produced healthy seedlings after removal of the fruit coat as well as after removal of both fruit and seed coat. These results clearly indicate that seeds of this species have the nondeep level of PD (Baskin and Baskin, 2004a).
As discussed above, D. trifoliata seeds are recalcitrant in seed storage behavior and further they have PD. Most species that produce recalcitrant seeds occur in year-round mesic environments or disperse seeds into mesic environments. Thus, seeds have no dormancy and they germinate soon after dispersal (Farnsworth, 2000; Pammenter and Berjak, 2000; Kermode and Finch-Savage, 2002; Asomaning et al., 2011). Although according to Kermode and Finch-Savage (2002), desiccation sensitive seeds are adapted to a regeneration strategy of rapid germination, some recalcitrant seeds are dormant (Farmer, 1977; Wigston, 1987; Carvalho et al., 1998; Tweddle et al., 2003; Ghasemi and Khosh-Khui, 2007; Jayasuriya et al., 2010) However, no recalcitrant seeds can have PY, because drying of the seed below the MC that recalcitrant seeds generally can tolerate is required for development of water impermeability of the seed (or fruit) coat (Qu et al., 2010).
The T50 value of D. trifoliata seeds with fruit coat intact was 71 days (Table 1). Thus, seeds with fruit coat have slow germination, which may be unfavorable for a species with recalcitrant seeds. However, D. trifoliata occurs in mangrove swamps, where water is present throughout the year. Further, the buoyant fruits of this species fall into the water and they are dispersed by water (Gehan Jayasuriya, personal observation). Fruits that fall onto the soil also are protected from desiccation by the fruit coat. Under ambient laboratory conditions, seeds enclosed by the fruit coat took 38 days to dry down to 15% MC. Thus, D. trifoliata should have a significant advantage by its seeds being physiologically dormant and recalcitrant. This combination of dormancy and recalcitrancy should allow seeds to persist in the seed bank for a period of time and to tolerate the high water content in its mangrove swamp habitat.
Derris is a widely distributed genus in southern and southeastern Asia (Mabberley, 1997); D. parvifolia is the only species in the genus endemic to Sri Lanka (Rudd, 1991). Thus, D. parvifolia may have been derived from an ancestor of Derris after the ancestor of D. parvifolia came to the island of Sri Lanka. This species produces nondormant seeds but occurs in a seasonal-dry climatic zone. However, the fruit is dispersed just before the rainy season begins and thus the nondormant seeds can germinate soon after they mature/disperse. Further, the fruit coat of D. parvifolia is flattened, papery and indehiscent and the fruits have a low mass. Thus, the seeds would be dispersed away from the mother plant by wind.
Derris trifoliata and D. scandens occur in marshy areas. Both produce a large number of seeds and although most seeds are dispersed by water a significant quantity of them falls onto the ground below the parent lianas. Thus, dormancy in seeds of these two species may be advantageous in dispersing germination over time. Further, dormancy of water-dispersed seeds would allow separation between the dispersal event and germination. When seeds fall into water and have no dormancy, they could imbibe and germinate. There is very little difference in temperature between seasons in the habitats of D. trifoliata and D. scandens. Further, prevailing temperatures are favorable for seed germination year-round. Consequently, if the seeds had no dormancy they would germinate on water during the dispersal event, where conditions for seedling growth are unfavorable. Thus, these two species use different mechanisms to avoid germination in conditions unfavorable for seedling establishment.
Fresh D. scandens seeds have a water impermeable seed coat; therefore, the embryo is not hydrated and cannot germinate. On the other hand, fresh seeds of D. trifoliata have a water-permeable seed coat and thus can imbibe water; however, the embryo in fresh seeds has physiological dormancy (PD, low growth potential) and cannot overcome the resistance of the fruit coat and germinate. Further, seed storage behaviour of D. trifoliata is recalcitrant and that of D. scandens is orthodox. Thus, seed dormancy in these two species seems to be connected to storage behaviour. The recalcitrant seeds of D. trifoliata cannot be physically dormant and the physically dormant seeds of D. scandens cannot be recalcitrant.
CONCLUSION
Seeds of D. parvifolia and D. scandens are orthodox in storage behavior, while those of D. trifoliata are recalcitrant. D. parvifolia seeds have no dormancy, whereas seeds of D. scandens have PY and those of D. trifoliata PD. Although D. scandens and D. trifoliata share similar habitats, they have evolved two different dormancy strategies to prevent seeds from germinating, while they are being dispersed via water. The dormancy strategies of these two species are determined by storage behavior. Derris is the only known genus with species producing seeds that belong to three different dormancy classes and we are not aware of any genus containing species that produce seeds with PY as well as species that produce seeds with PD.
REFERENCES
- Adams, C.A., J.M. Baskin and C.C. Baskin, 2005. Trait stasis versus adaptation in disjunct relict species: Evolutionary changes in seed dormancy-breaking and germination requirements in a subclade of Aristolochia subgenus Siphisia (Piperales). Seed Sci. Res., 15: 161-173.
CrossRef - Alderete-Chavez, A., L. Aguilar-Marin, N. De la Cruz-Landero, J.J. Guerra-Santos, R. Brito, E. Guevara and R. Gelabert, 2010. Effects of scarification chemical treatments on the germination of Crotalaria retusa L. seeds. J. Biol. Sci., 10: 541-544.
CrossRefDirect Link - Asomaning, J.M., N.S. Olympio and M. Sacande, 2011. Desiccation sensitivity and germination of recalcitrant Garcinia kola Heckel seeds. Res. J. Seed Sci., 4: 15-27.
Direct Link - Balouchi, H.R. and S.A.M.M. Sanavy, 2006. Effect of gibberellic acid, prechilling, sulfuric acid and potassium nitrate on seed germination and dormancy of annual Medics. Pak. J. Biol. Sci., 9: 2875-2880.
CrossRefDirect Link - Baskin, C.C., 2003. Breaking physical dormancy in seeds-focussing on the lens. New Phytol., 158: 229-232.
CrossRefDirect Link - Baskin, J.M. and C.C. Baskin, 2004. A classification system for seed dormancy. Seed Sci. Res., 14: 1-16.
CrossRefDirect Link - Baskin, J.M., C.C. Baskin and X. Li, 2000. Taxonomy, anatomy and evolution of physical dormancy in seeds. Plant Species Biol., 15: 139-152.
CrossRefDirect Link - Carvalho, J.E.U., C.H. Muller and N.V.M. Leao, 1998. Chronology of morphological events associated to germination and desiccation sensitivity of Platonia insignis Mart. seeds: Clusiaceae. Rev. Brasil. Sementes, 20: 236-240.
Direct Link - Farmer, R.E., 1977. Epicotyl dormancy in white and chestnut oaks. For. Sci., 23: 329-332.
Direct Link - Farnsworth, E., 2000. The ecology and physiology of viviparous and recalcitrant seeds. Annu. Rev. Ecol. Syst., 31: 107-138.
CrossRefDirect Link - Gama-Arachchige, N.S., J.M. Baskin, R.L. Geneve and C.C. Baskin, 2010. Identification and characterization of the water gap in physically dormant seeds of Geraniaceae, with special reference to Geranium carolinianum. Ann. Bot., 105: 977-990.
CrossRef - Gama-Arachchige, N.S., J.M. Baskin, R.L. Geneve and C.C. Baskin, 2011. Acquisition of physical dormancy and ontogeny of the micropyle-water-gap complex in developing seeds of Geranium carolinianum (Geraniaceae). Ann. Bot., 108: 51-64.
CrossRef - Gilbert, G., 1952. Contribution a la biologie des essences forestieres congolaises. Bull. R. Bot. Soc. Belgium, 84: 289-296.
Direct Link - Jayasuriya, K.M.G.G., J.M. Baskin and C.C. Baskin, 2008. Dormancy, germination requirements and storage behaviour of seeds of Convolvulaceae (Solanales) and evolutionary considerations. Seed Sci. Res., 18: 223-237.
CrossRef - Jayasuriya, K.M.G.G., J.M. Baskin and C.C. Baskin, 2009. Sensitivity cycling and its ecological role in seeds with physical dormancy. Seed Sci. Res., 19: 3-13.
CrossRefDirect Link - Jayasuriya, K.M.G.G., A.S.T.B. Wijetunga, J.M. Baskin and C.C. Baskin, 2010. Recalcitrancy and a new kind of epicotyl dormancy in seeds of the understory tropical rainforest tree Humboldtia laurifolia (Fabaceae, Ceasalpinioideae). Am. J. Bot., 97: 15-26.
CrossRefDirect Link - Kumar, S., J. Radhamani, A. Singh and K.S. Varaprasad, 2007. Germination and seed storage behavior in Pongamia pinnata. Curr. Sci., 93: 910-911.
Direct Link - Meisert, A., D. Schulz and H. Lehmann, 1999. Structural features underlying hardseededness in geraniaceae. Plant Biol., 1: 311-314.
CrossRef - Mullett, J.H., D.V. Beardsell and H.M. King, 1981. The effect of seed treatment on the germination and early growth of Euterpe edulis (family Palmae). Scient. Hortic., 15: 239-244.
CrossRef - Pammenter, N.W. and P. Berjak, 2000. Some thoughts on the evolution and ecology of recalcitrant seeds. Plant Spec. Biol., 15: 153-156.
CrossRefDirect Link - Qu, X., J.M. Baskin and C.C. Baskin, 2010. Whole-seed development in Sicyos angulatus (Cucurbitaceae, Sicyeae) and a comparison with that of water-impermeable seeds in five other families. Plant Spec. Biol., 258: 185-192.
CrossRefDirect Link - Sarker, P.C., S.M.A. Hossain, M.S.U. Bhuiya and M. Salim, 2000. Breaking seed dormancy in Sesbania rostrata. Pak. J. Biol. Sci., 3: 1801-1802.
CrossRefDirect Link - Sautu, A., J.M. Baskin, C.C. Baskin, J. Deago and R. Condit, 2007. Classification and ecological relationships of seed dormancy in a seasonal moist tropical forest, Panama, Central America. Seed Sci. Res., 17: 127-140.
CrossRef - Taylor, G.B., 2005. Hardseededness in Mediterranean annual pasture legumes in Australia: A review. Aust. J. Agric. Res., 56: 645-661.
CrossRef - Teketay, D., 1998. Germination of Acacia origena, A. pilispina and Pterolobium stellatum in response to different pre-sowing seed treatments, temperature and light. J. Arid Environ., 38: 551-560.
CrossRefDirect Link - Travlos, I.S. and G. Economou, 2006. Optimization of seed germination and seedling emergence of Medicago arborea L. Int. J. Bot., 2: 415-420.
CrossRefDirect Link - Tweddle, J.C., J.B. Dickie, C.C. Baskin and J.M. Baskin, 2003. Ecological aspects of seed desiccation sensitivity. J. Ecol., 91: 294-304.
CrossRefDirect Link - Eira, M.T.S., E.A.A. da Silva, R.D. de Castro, S. Dussert, C. Walters, J.D. Bewley and H.W.M. Hilhorst, 2006. Coffee seed physiology. Braz. J. Plant Physiol., 18: 149-163.
Direct Link - Uzun, F. and I. Aydin, 2004. Improving germination rate of Medicago and Trifolium species. Asian J. Plant Sci., 3: 714-717.
CrossRefDirect Link - Ng, F.S.P., 1980. Germination ecology of Malaysian woody plants. Malaysian Forester, 43: 406-438.
Direct Link - Gleiser, G., M.C. Picher, P. Veintimilla, J. Martinez and M. Verdu, 2004. Seed dormancy in relation to seed storage behaviour in Acer. Botanical J. Linnean Soc., 145: 203-208.
CrossRef - Ghasemi, M. and M. Khosh-Khui, 2007. Effect of stratification and growth regulators on seed germination and seedling growth of Quercus ilex L. J. Plant Sci., 2: 341-346.
CrossRef
Dr.P.k.Nagar Reply
As such paper is good but the authors should have conducted some more physiol studies like moisture content ABA and related dormancy stidies. I would suggest that the email addrss of corresponding author may also b given