
A. Alderete-Chavez
Centro de Investigacion de Ciencias Ambientales, Universidad Autonoma del Carmen, Calle 56 No. 4, Carmen, Campeche, Mexico CP 24180, Mexico
N. De la Cruz-Landero
Centro de Investigacion de Ciencias Ambientales, Universidad Autonoma del Carmen, Calle 56 No. 4, Carmen, Campeche, Mexico CP 24180, Mexico
J.J. Guerra-Santos
Centro de Investigacion de Ciencias Ambientales, Universidad Autonoma del Carmen, Calle 56 No. 4, Carmen, Campeche, Mexico CP 24180, Mexico
E. Guevara
Centro de Investigacion de Ciencias Ambientales, Universidad Autonoma del Carmen, Calle 56 No. 4, Carmen, Campeche, Mexico CP 24180, Mexico
R. Gelabert
Centro de Investigacion de Ciencias Ambientales, Universidad Autonoma del Carmen, Calle 56 No. 4, Carmen, Campeche, Mexico CP 24180, Mexico
Lizbeth R. De la Cruz-Magana
Centro de Investigacion de Ciencias Ambientales, Universidad Autonoma del Carmen, Calle 56 No. 4, Carmen, Campeche, Mexico CP 24180, Mexico
E. Nunez-Lara
Centro de Investigacion de Ciencias Ambientales, Universidad Autonoma del Carmen, Calle 56 No. 4, Carmen, Campeche, Mexico CP 24180, Mexico
R. Brito
Centro de Investigacion de Ciencias Ambientales, Universidad Autonoma del Carmen, Calle 56 No. 4, Carmen, Campeche, Mexico CP 24180, Mexico
Research Journal of Seed Science
Year: 2011 | Volume: 4 | Issue: 1 | Page No.: 51-57







ABSTRACT
The effectiveness of scarification treatments in the promotion of germination of Bauhinia divaricata was determine to suggest an effective and practical method for breaking seed dormancy. Seeds were collected on March 2010, obtained from disturbed forests and tested with different exposition times (0, 7, 15, 20 and 30 min) by their immersion into sulfuric acid during April and May 2010, assays were done in the Botanical Garden of the Universidad Autonoma del Carmen, day/night regime of 32/18°C was taken into consideration along with a 12 h photoperiod. The germination was observed daily early in the morning during 30 days. All treatments started their germination three days after the beginning of the experiment and reached their maximum value between 25 and 30 days after the exposition. Seeds treated with sulphuric acid, responded positively in all cases but, the immersion in acid for 15 or 20 min were the treatment that showed the best results, for this reason these treatments are recommended to improve and to efficient the germination of seeds of this species.
PDF Abstract XML References Citation
Received: September 07, 2010;
Accepted: October 16, 2010;
Published: February 26, 2011
How to cite this article
A. Alderete-Chavez, N. De la Cruz-Landero, J.J. Guerra-Santos, E. Guevara, R. Gelabert, Lizbeth R. De la Cruz-Magana, E. Nunez-Lara and R. Brito, 2011. Promotion of Germination of Bauhinia divaricata L. Seeds by Effects of Chemical Scarification. Research Journal of Seed Science, 4: 51-57.
URL: https://scialert.net/abstract/?doi=rjss.2011.51.57
URL: https://scialert.net/abstract/?doi=rjss.2011.51.57
INTRODUCTION
Bauhinia is a large pantropical genus representative of the tribe Cercideae that includes more than 50 species in Mexico and Central America, several are endemic, their species can be clearly distinguished form the other sorts of the subfamily Caesalpinioideae, the genus Bauhinia is the only native representative of the tribe Cercideae at Yucatan peninsula with 5 wild species. In Campeche State there is only one species, Bauhinia divaricata, growing in areas degraded by erosion on infertile and bad quality soils in secondary vegetation characterized by a mosaic of habitats where environmental factors have been changed (Auda, 2010). Bauhinia divaricata is an shrub from 1 to 8 m height. The leaves are wider than long and sometimes divided in two lobules seemed to a goat leg, pale green color. The flowers are white. The fruits of 5 to 12 cm of length, with 5 to 10 seeds; species can be used as ornamental due its high economical value; with useful for respiratory diseases like cold, pulmonary affections and asthma. The leaves are used on wounds, like healing and in bruises like anti-inflammatory disinfectant and, also it is used in cases of fright and the on childbirth and distributed throughout in Yucatan peninsula (Colin, 2006).
Seed propagation is the most common method for production of many ligneous plants; however, commonly latency problems exist that make difficult the germination and emergence. The seminal latency is one of the main problems for plant production (Reyes and Casal, 2006). In many species the low rate of germination is associate to the hardening of the seminal covers (Dayamba et al., 2010), what it makes almost impermeable to the entrance of oxygen and water and in some species the covers can be a mechanical barrier for germination or can contain chemical compound that inhibits the process (Ruprecht et al., 2008).
Most species of Fabaceae produce seeds with physical dormancy which was broken by some environmental factors in the field, yet for the mechanism of physical dormancy release in the natural condition is still poorly understood (Wen et al., 2009; Baskin et al., 2006). Physical dormancy and the conditions under which it is released under natural conditions, remains poorly understood, especially in tropical regions (Rieks et al., 2006). Seed dormancy in different plant families could be endogenous or exogenous in origin. In endogenous dormancies some characteristic of the embryo like endosperm, fruit walls or seed coats covering the embryo or embryo immaturity prevents germination. In exogenous dormancies some physical, chemical or mechanical factors such as impermeable seed coats, germination inhibitors and woody structures inhibit seed germination (Razavi and Hajiboland, 2009).
In the wild, physical dormancy release could be not only the effects of the gastric juice of birds. Frugivorous birds disperse the seeds of many fruit-bearing plants, but the fate of seeds after defecation or regurgitation is often unknown. Some rodents gather and scatter hoard seeds and some of these may be overlooked, germinate and establish plants (Ness and Bressmer, 2009; Stephen et al., 2005; Cote et al., 2003).
The function of the seed coat is to protect the embryo and endosperm from desiccation, mechanical injury, unfavorable temperatures and insects or diseases (Liu et al., 2009). Similarly to many other taxa of legumes, the seeds of Bauhinia plants have a low and irregular germination which is attributed mainly to the impermeability of the seeds to water (Zhi-Quan et al., 2007; Taylor, 2005; Mena-Ali and Rocha, 2005; Tauro et al., 2009), the seeds remain in physical dormancy until the seed coat is permeable by some environmental factors in natural conditions (Jayasuriya et al., 2007). Many investigations have been done by testing methods to overcome the physical dormancy of Bauhinia seeds (Scott et al., 2010; Bhagirath and Johnson, 2009).
The purpose of this study was to know the effect of scarification with sulphuric acid on the germination potential of Bauhinia divaricata, to propose an effective and practical method for breaking seed dormancy.
MATERIALS AND METHODS
The seeds of Bauhinia divaricata were obtained from disturbed forests in, Municipality of Carmen, Campeche State, Mexico (18° 37’ 54 N and 91° 54’ 14” W), at 1.84 m above sea level, on slopes of 4-7%. The mature seeds were collected from twenty four plants on March 2010.
The laboratory assays were done in the Seed laboratory of Natural Sciences School, Universidad Autónoma del Carmen, Campeche State, Mexico, from April to May 2010. A day/night regime of 32/18°C was taken into consideration along with a 12 h photoperiod. Pregerminative treatment considered the immersion of seeds in 98% Sulphuric acid (H2SO4), during 0 (control), 7, 15, 20 and 30 min after which seeds were washed with distilled water.
A total of 300 seeds (ten seeds with 6 replications for treatments) were sown in each experimental unit (Petri dishes with Agrolite as substrate). The germination was observed daily early in the morning for a period of 30 days. The seeds were considered germinated when the radicle reached at least the length of the seed. Seeds with roots or cotyledons malformed were not considered for the test. The seeds were irrigated with distilled water and a 3% Captan solution.
The experimental design was completely randomized blocks with six replications. Germination was registered by replica per treatment during the experiment. Germination frequencies for the same treatment were grouped and expressed as percentage. The SAS program (v. 2003) for microcomputers was employed to conduct the ANOVA test (Proc mixed) and mean comparisons LSD (Less Significant Difference). The treatments were considered as fixed effects, the blocks as random effects.
RESULTS
The results obtained on the final percentage and time of emergence in B. divaricata, after to apply the germinative treatments (Table 1). Percentage of emergency was affected by chemical scarification highest values corresponded to the application of sulfuric acid by 15 min. The time in which the 50 % of the total emergency (E50) observed was smaller in the control.
All treatment applied were statistically significant (p = 0.05). The radicles of B. divaricata started their germination three days after they were treated with acid solution. The seeds with immersion in 98% H2SO4, shown positively effect in all cases compared with the control when the temperature was under 32/18°C day/night. When the H2SO4 was used, germination percentage is over 57% at day 29, in all seeds treated in contrast with 20% obtained at the control (Fig. 1).
In concordance with statistical analysis, seeds exposed to immersion for 7, 15, 20, or 30 min showed different behavior at the end of the experiment, however when the seed are treated for 15 min, the range of germination percentages by replica are very short in contrast with the other treatments and germination rate is increased, reaching more than 13% at day 3 while the same result were obtained after 5 and 7 days in the remaining assays. The other treatments got no more than 57% of germination at the end of the assay. All treated seeds had significantly higher germination percentages than the control which only obtained 20% of germination at the end of the assays (Fig. 1).
The trend observed at different treatments shown that conditions proposed for the experiments are adequate for germination and the application of 98% H2SO4 for 15 and 20 min, significantly increased the germination of B. divaricata.
| Table 1: | Effect of seeds scarification on the final percentage and times of emergency in Bauhinia divaricata |
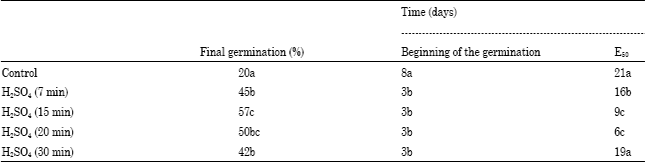 | |
| Separation of averages in the columns according to the test of Tukey (p≤0.05). Means with some letter(s) are not significantly different | |
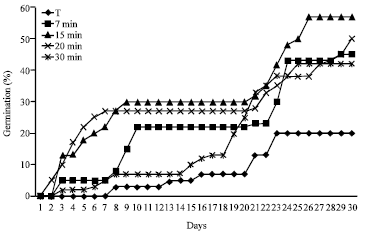 | |
| Fig. 1: | Curves of accumulated germination frequency (%) of B. divaricata with chemical Scarification during 30 days of experiment |
DISCUSSION
The seeds of Bauhinia obtained from a natural environment and the H2SO4 scarification treatments simulated pass of the seeds through the digestive tract of animals (v. gr. birds and rodents) (Amerah et al., 2007), The effects of ingestion by birds can be due to the grinding effect in the gizzard, the acidic treatment in the gut or both (Santamaria et al., 2002), which under natural conditions chemical scarification is needed (Chavez et al., 2010).
Seed dormancy is an innate seed property that defines the environmental conditions in which the seed is able to germinate, dormancy is a common attribute of many weed seed populations and this hampers the task of predicting emergence from weed seed banks under field conditions (Batlla and Benech-Arnold, 2007). Otherwise, characteristics of seeds as coat thickness and coat hardness, time and conditions where they are stored and treatment previously offered are involved at sprouting (Martinez et al., 2007).
Effect of dormancy breaking method on germination rate depends on the plant species, thus it can be suggested that the most effective method in breaking the dormancy of forage legumes should be determined (Balouchi and Sanavy, 2006) by the application or each method with different period, concentration or degree combinations to the seeds in order to maximum germination rate. Our results demonstrate that H2SO4 treating were able to enhance germination because seed dormancy in B. divaricata is reduced when the acid act over the seeds coat improving germination percentage (Gama-Arachchige et al., 2010).
Higher percentage of germination with chemical scarification (Sulphuric acid) for 15 min at temperatures of 25/15°C with light (82.3% of germination) were reported in Lupinus leptophyllus (Chavez et al., 2010). Chemical germination was found in seeds of Calystegia soldanella pretreated with H2SO4 for 3 h (Ko et al., 2004). Chemical applications also improve germination in Calystegia soldanella. A positive effect of H2SO4 was observed when the seeds were pretreated, reaching about 90 % of germination (Baskin et al., 1998). Pretreatments applications of H2SO4 for 20 min, also improve germination in Capparis spinosa (Sozzi and Chiesa, 1995).
The results of this research that simulates the effects of the pass throughout animal digestive tract on seeds germination and from the literature reviewed it is inferred that B. divaricata have about 20% germinative efficiency in natural conditions (without treatment) and when the seeds are consumed by rodents and other animals, the digestion process helps scarification and the dispersion of the seeds throughout the ecosystem.
The positive response to sulfuric acid treatments indicates that the impermeable coat is responsible of low germination rates from intact seeds as showed at experimental untreated controls. By overcoming the physical dormancy with chemical pretreatment, seed coats are softened and water uptake is enabled. Both actions are crucial to sustaining the life cycle of hard-seeds species. These results demonstrate that the seeds of this species and some others in the same sort, present/display a condition of tegument latency. This type of latency is frequent in legumes.
CONCLUSIONS
The treatments of chemical scarification accelerated and standardized the emergency process and increased their final values. Germination was affected by all H2SO4 scarification treatments. The treatment of immersion in sulphuric acid for 15 min gave 57% of germination after 30 days seeds. Therefore it is recommendable to apply treatments with sulfuric acid, to increase the percentage of germination of B. divaricata. These results can be due to that the sulfuric acid maybe perforates the layer of macroesclereidas allowing that imbibitions happen.
ACKNOWLEDGMENTS
Special thanks to PROMEP granting the internal project number PROMEP/103.5/09/416 and thanks to CONACyT for financing project FORDECyT No. 137942 without financing the development of this research would not be possible and Universidad Autónoma of Carmen by the granted facilities to carry out this work.
REFERENCES
- Amerah, A.M., V. Ravindran, R.G. Lentle and D.G. Thomas, 2007. Feed particle size: Implications on the digestion and performance of poultry. Worlds Poult. Sci. J., 63: 439-455.
CrossRefDirect Link - Auda, M.M.A., 2010. Contribution to the plant ecology and the most palatable species for grazing in the gaza strip mediterranean coast, Palestine. Asian J. Plant Sci., 9: 88-93.
CrossRef - Balouchi, H.R. and S.A.M.M. Sanavy, 2006. Effect of gibberellic acid, prechilling, sulfuric acid and potassium nitrate on seed germination and dormancy of annual Medics. Pak. J. Biol. Sci., 9: 2875-2880.
CrossRefDirect Link - Baskin, C.C., K. Thompson and J.M. Baskin, 2006. Mistakes in germination ecology and how to avoid them. Seed Sci. Res., 16: 165-168.
CrossRefDirect Link - Baskin, J.M., N. Xiaoying and C.C. Baskin, 1998. A comparative study of seed dormancy and germination in an annual and a perennial species of Senna (Fabaceae). Seed Sci. Res., 8: 501-512.
Direct Link - Batlla, D. and R.L. Benech-Arnold, 2007. Predicting changes in dormancy level in weed seed soil banks: Implications for weed management. Crop Protection, 26: 189-196.
CrossRef - Bhagirath, S.C. and D.E. Johnson, 2009. Germination, emergence and dormancy of Mimosa pudica. Weed Biol. Manage., 9: 38-45.
CrossRefDirect Link - Alderete-Chavez, A., D.A. Rodriguez-Trejo, V. Espinosa-Hernandez, E. Ojeda-Trejo and N. de la Cruz-Landero, 2010. Effects of different scarification treatments on the germination of Lupinus leptophyllus Seeds. Int. J. Botany, 6: 64-68.
CrossRefDirect Link - Colin, T.R., 2006. Two new species of Bauhinia (fabaceae, caesalpinioideae, cercideae) from Southeastern Mexico and Mesoamerica. Novon J. Botanical Nomenclature, 16: 533-537.
CrossRef - Cote, M., J. Ferron and R. Gagnon, 2003. Impact of seed and seedling predation by small rodents on early regeneration establishment of black spruce. Can. J. For. Res., 33: 2362-2371.
CrossRef - Dayamba, D.S., L. Sawadogo, M. Tigabu, P. Savadogo, D. Zida, D. Tiveau and P. C. Oden, 2010. Effects of aqueous smoke solutions and heat on seed germination of herbaceous species of the Sudanian savanna-woodland in Burkina Faso. Flora Morphol. Distribution Functional Ecol. Plants, 205: 319-325.
CrossRefDirect Link - Gama-Arachchige, N.S., J.M. Baskin, R.L. Geneve and C.C. Baskin, 2010. Identification and characterization of the water gap in physically dormant seeds of Geraniaceae, with special reference to Geranium carolinianum. Ann. Bot., 105: 977-990.
CrossRef - Jayasuriya, K.M.G.G., J.M. Baskin, R.L. Geneve and C.C. Baskin, 2007. Morphology and anatomy of physical dormancy in Ipomoea lacunosa: Identification of the water gap in seeds of Convolvulaceae (Solanales). Ann. Bot., 100: 13-22.
CrossRefDirect Link - Ko, J.M., H.J. Park, B.M. Min and H.C. Cha, 2004. Effects of various pretreatments on seed germination of Calystegia soldanella (Convolvulaceae), a coastal sand dune plant. J. Plant Biol., 47: 396-400.
Direct Link - Liu, H., L. Yu-Jun, Y. Ming-Feng and S. Shi-Hua, 2009. A comparative analysis of embryo and endosperm proteome from seeds of Jatropha curcas. J. Integrative Plant Biol., 51: 850-857.
CrossRef - Martinez, I., D. Garcia and J.R. Obeso, 2007. Allometric allocation in fruit and seed packaging conditions the conflict among selective pressures on seed size. Evol. Ecol., 21: 517-533.
CrossRef - Mena-Ali, I.J. and O.J. Rocha, 2005. Selective seed abortion affects the performance of the offspring in Bauhinia ungulate. Ann. Bot., 95: 1017-1023.
CrossRef - Ness, H.J. and K. Bressmer, 2009. Abiotic influences on the behaviour of rodents, ants, and plants effect an ant-seed mutualism. Ecoscience, 12: 76-81.
CrossRef - Razavi, S.M. and R. Hajiboland, 2009. Dormancy breaking and germination of Prangos ferulaceae seeds. Eur. Asian J. BioSci., 3: 78-83.
CrossRef - Reyes, O. and M. Casal, 2006. Seed germination of Quercus robur, q. pyrenaica and q. ilex and the effects of smoke, heat, ash and charcoal. Ann. For. Sci., 63: 205-212.
CrossRef - Rieks, D., V. Klinken, L.K. Flack and W. Pettit, 2006. Wet-season dormancy release in seed banks of a tropical leguminous shrub is determined by wet heat. Ann. Bot., 98: 875-883.
CrossRef - Ruprecht, E., T.W. Donath, A. Otte and R.L. Eckstein. 2008. Chemical effects of a dominant grass on seed germination of four familial pairs of dry grassland species. Seed Sci. Res., 18: 239-248.
CrossRef - Santamaria, L., I. Charalambidou, J. Figuerola and A.J. Green, 2002. Effect of passage through duck gut on germination of fennel pondweed seeds. Arch. Hydrobiol., 156: 11-22.
CrossRef - Scott, K., S. Setterfield, M. Douglas and A. Andersen, 2010. Soil seed banks confer resilience to savanna grass-layer plants during seasonal disturbance. Acta Oecol., 36: 202-210.
CrossRef - Sozzi, G.O. and A. Chiesa, 1995. Improvement of caper (Capparis spinosa L.) seed germination by breaking seed coat-induced dormancy. Scient. Hortic., 62: 255-261.
CrossRefDirect Link - Stephen, B., V. Wall, K.M. Jun and J.R. Gworek, 2005. Two-phase seed dispersal: Linking the effects of frugivorous birds and seed-caching rodents. Oecologia, 145: 281-286.
CrossRef - Taylor, G.B., 2005. Hardseededness in Mediterranean annual pasture legumes in Australia: A review. Aust. J. Agric. Res., 56: 645-661.
CrossRef - Tauro, T.P., H. Nezomba, F. Mtambanengwe and P. Mapfumo, 2009. Germination, field establishment patterns and nitrogen fixation of indigenous legumes on nutrient-depleted soils. Symbiosis, 48: 92-101.
Direct Link - Wen, H.X., P.Y. Wu and R.Y. Wang, 2009. Different requirements for physical dormancy release in two populations of Sophora alopecuroides relation to burial depth. Ecol. Res., 24: 1051-1056.
CrossRef - Zhi-Quan, C.A., P. Lourens, C.A. Kun-Fang and F. Bongers, 2007. Seedling growth strategies in bauhinia species: Comparing lianas and trees. Ann. Bot., 100: 831-838.
CrossRef