
S. Mondal
Department of Plant Physiology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi-221005, India
P. Vijai
Department of Plant Physiology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi-221005, India
B. Bose
Department of Plant Physiology, Institute of Agricultural Sciences, Banaras Hindu University, Varanasi-221005, India
Research Journal of Seed Science
Year: 2011 | Volume: 4 | Issue: 3 | Page No.: 157-165







ABSTRACT
In the present study the effects of hydro and halo hardening of seeds with distilled water and Mg(NO3)2, respectively, were observed on the performance of rice (Oryza sativa L.) var. Swarna (MTU 7029) starting from its germination to vegetative growth. Maximum amylase activity in endosperm correlated well with an increase in soluble sugar content were noted in halo hardened seeds of rice, whereas in the same treatment the insoluble content was noted to reduce maximum in comparison to hydro hardened and non-hardened sets. Studied growth parameters like plumule/shoot length, radicle/root length, fresh and dry weights of seedling, root number, proline content, total nitrogen content in seedlings and nitrate reductase activity in leaves were found maximum in Mg(NO3)2 hardened one followed by other two treatments.
PDF Abstract XML References Citation
Received: February 20, 2011;
Accepted: April 25, 2011;
Published: July 09, 2011
How to cite this article
S. Mondal, P. Vijai and B. Bose, 2011. Role of Seed Hardening in Rice Variety Swarna (MTU 7029). Research Journal of Seed Science, 4: 157-165.
URL: https://scialert.net/abstract/?doi=rjss.2011.157.165
URL: https://scialert.net/abstract/?doi=rjss.2011.157.165
INTRODUCTION
Rice (Oryza sativa L.) is the prime source of food for nearly half of the World’s population and it is one of the most important major food crops (Kumar et al., 2008). Rice contributes 43% of total food grain production and 46% of total cereal production in India. It continues to play vital role in the national food grain supply. It is a staple food of nearly half of the world population. Rapid and uniform field emergences of seedlings are two essential pre-requisites to increase yield and quality in a number of field crops (Krishnotar et al., 2009). Seed priming/hardening (pre sowing imbibition treatment) is widely used to enhance seed performance with respect to rate and uniformity of germination (De Lespinay et al., 2010). These treatments are based upon controlled hydration of the seeds. During priming, seeds are brought up to and eventually held at the end of phase II of water uptake of the germination process without entering the initial growth phase characterized by radical protrusion through the seed coat.
The beneficial effects of seed harderning are primarily due to pre-enlargement of the embryo, biochemical changes like enzyme activation and improvement of germination rate (Lee and Kim, 2000; Basra et al., 2005). Recently Bose and Mishra (2001), Sharma and Bose (2006) and Bose et al. (2007) did some work with nitrate seed priming/hardening technology where the seeds of various field crops like wheat, maize and mustard were hardened with different nitrate salts [(Mg(NO3)2 and KNO3] before sowing. They observed an improvement in germination physiology, vegetative growth and yield of these crops with hardening of seeds with nitrate salts; hence it is known as nitrate seed hardening technology. Further, Farooq et al. (2006a, b) also introduced a technique for rice seed invigoration that successfully integrated hardening and osmoconditioning. The process was named osmohardening of rice. However, this type of technology is of immense importance because it minimizes the use of chemicals and avoids discarding materials those may be undesirable to the environment (McDonald, 2000). Harris et al. (2001), Ghassemi-Golezani et al. (2008a, b) did a lot of studies in form of participatory research work with farmers and suggested that this type of seed technology is simple, low cost and low risk intervention and also had positive impacts on the wider farming system and livelihood. Seed priming also helps to restore/activate the performance of various stress regulating enzymes/systems (osmolites) (Bailly et al., 1997; Omidi, 2010). Therefore, this nitrate seed hardening technology may also be introduced to improve and overcome the various constraints related to rice crop growth. Hence, in the present investigation the most popular and highest yielding variety Swarna (MTU 7029) was taken into consideration with an objective to study the impact of seed hardening treatments with distil water and magnesium nitrate salt on some important parameters, regulating germination as well as early seedling growth.
MATERIALS AND METHODS
The present study was carried out in Seed Physiology Laboratory at Department of Plant Physiology, I.Ag.Sc., BHU, India, in the year 2008-09 in a Randomized Block Design (RBD) with 4 replication. Seeds were procured from the Genetics and Plant Breeding Department of the same institute.
This experiment was conducted in the presence of diffused light condition at an average temperature of about 30-35°C during kharif season.
Healthy bold seeds were surface sterilized with 0.1% HgCl2 (Mercuric chloride) solution for 2 min then washed thoroughly with distilled water for 5-6 times. For hardening treatment the seeds were either kept in beaker having distilled water (T2) (hydrohardening) or varying concentrations of Mg(NO3)2 (halohardening) [2, 4 and 8 mM (T3, T4 and T5)] for 20 h. After that the seeds were taken out and gently washed with distilled water once and then dried back to its initial weight at the room temperature by placing them under fan. After that these seeds were packed in paper bags and were used as per requirement but within one month of treatment. The seeds without any treatment referred as control (T1).
The morphophysiological and biochemical observation during germination and vegetative growth in hydro and halo hardened seeds were measured in different days according to the requirement. Rate of germination was measured by using following formula:
Radicle/root length of the longest root was measured from its physiological base to tip by using scale after proper washing of root. The number of roots was counted by placing the root part on a glass plate with the help of a needle. Plumule/shoot length was measured from its physiological base to the top of the longest leaf. Dry weight of seedlings were obtained by drying at 60-70°C till constant weight was achieved. Amylase, soluble and insoluble sugar content of endosperm, nitrate reductase activity of leaves, proline and total nitrogen content of seedlings were measured by using the method of Bernfeld (1955), Dubois et al. (1956), Srivastava (1974), Bates et al. (1973) and Lang (1958), respectively.
Statistical analysis: The data obtained from various observation were subjected to statistical analysis by adopting appropriate method of Analysis of Variance for Randomized Block Design. The significance of treatment effect was tested with the help of F- test and the differences between treatments by Critical Difference (C.D) at 1 and 5% level of significance were determined.
RESULTS AND DISCUSSION
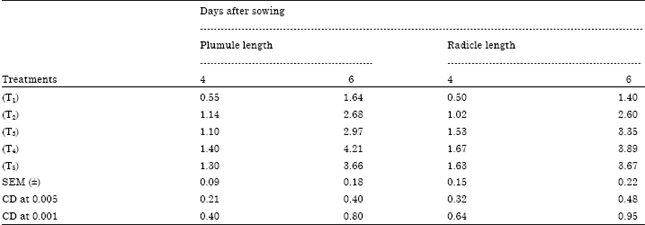
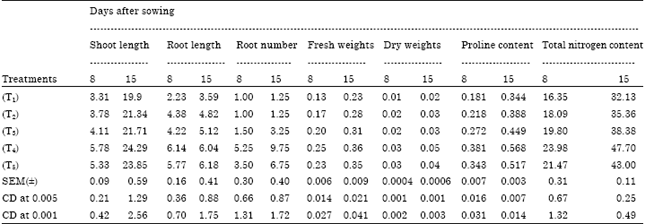
The germination of rice seeds was found to increase gradually upto 72 h. The T4 sets achieved the maximum germination percentage at 48 h and 72h (95 and 98.75%, respectively) where as control sets reached to its maximum level only at 72 h (92 %) (Fig. 1). Plumule and radicle lengths in 4 and 6 day old germinating rice seeds were found to maximum in T4 hardened sets (1.4, 4.21, 1.67 and 3.89 cm, respectively). That was followed by T5, T3 and T2 sets but the control represented the poorest performance among all (Table 1). The amylase activity of endosperm obtained from 3, 5 and 7 days old seedling was assayed. Maximum activity of amylase enzyme at 3 days old seedling was represented by T4 (298.89 mg maltose g-1 h-1) treatment followed by T5 (274.07 mg maltose g-1 h-1) and T3 (265.42 mg maltose g-1 h-1) whereas in 5 and 7 days old seedlings the maximum activity was observed in control sets (239.59 mg maltose g-1 h-1 and 118.33 mg maltose g-1 h-1) (Fig. 2). Soluble sugar content (Fig. 3) showed an improvement in Mg(NO3)2 invigorated seeds with increasing hours of germination followed by distilled water treated one; control was always found poor performer in this respect (5.67, 17.27 and 22.97 mg g-1 of 3, 5 and 7 days old endosperm). The insoluble sugar content was found highest in case of control sets (36.7, 23.82 and 17.75 mg g-1) in respect to other treatments at all the study periods (Fig. 4). Shoot and root length, root number, fresh and dry weights, proline content, total nitrogen content in seedlings (shoot+root) and nitrate reductase activity of leaves were studied at 8 and 15 days after sowing, respectively (Table 2, Fig. 5). For all the parameters the halo hardened sets performed better than the control one in both the days. Beside this, T4 was best performer than all other treatments and control for all studied parameters; followed by T5 but T5 showed at par results in many of the cases.
It is well known that Swarna is a popular variety but sometimes it suffers due to lack of proper irrigation and late sowing of seeds in its cropping season both of which may affect the growth of the crop by various means. Hence, better rates of germination followed by vigorous seedlings are desirable characters for the proper vegetative and reproductive growth of these plants.
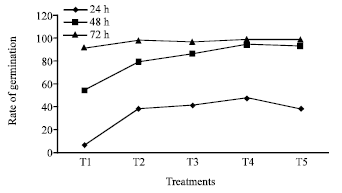 | |
| Fig. 1: | Effect of seed hardening with distilled water and different concentrations of Mg(NO3)2 on rate of germination of rice var. Swarna at different study period [C.D. at 1%= 12.29 (24 h), 2.23 (48 h), 6.05 (72 h)]. *T1- Control (non-hardened); T2- distilled water hardened; T3, T4 and T5- 2, 4 and 8 mM Mg(NO3)2 hardened |
| Table 1: | Effect of seed hardening with distilled water and different concentrations of Mg(NO3)2 on plumule and radicle length (cm) of seedlings of rice var. Swarna at different days after sowing (DAS) |
 | |
| *T1: Control (non-hardened), T2: Distilled water hardened, T3, T4 and T5: 2, 4 and 8 mM Mg (NO3)2 hardened | |
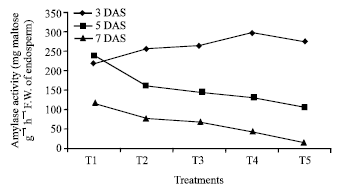 | |
| Fig. 2: | Effect of seed hardening with distilled water and different concentrations of Mg(NO3)2 on Amylase Activity of rice var. Swarna at different study periods of germination [C.D. at 1%= 5.17 (3 DAS), 6.63 (5 DAS), 15.34 (7 DAS)]. *T1- Control (non-hardened); T2- distilled water hardened; T3, T4 and T5- 2, 4 and 8 mM Mg(NO3)2 hardened |
Results of present investigation revealed that when the seeds of rice variety Swarna hardened with distilled awater and various concentrations (2-8 mM) of Mg(NO3)2 showed an improvement in rate of germination (%) as well as plumule and radicle length of the seedlings (Table 1). The study of amylase enzyme was also found to be positively correlated with the seed germination where the germination % of the nitrate hardened seeds reached to its maximum level at early hours (48 h) but control sets failed to achieve this status at that period. From the study it has also been realised that the amylase activity in control sets achieve their maximum level in 6 day old germinating seeds whereas it was found to decline in all the sets of hardened seeds at that time (Fig. 2). This might be possible due to an exhaustive utilization of amylase enzyme activity during the early and enhanced rate of germination in hardened sets as compared to control one. The increased activity of enzyme amylase further established a positive correlation with the increasing amount of soluble sugar in germinating seeds of hardened sets (Fig. 3). The study regarding the change in insoluble sugar, the reserve food material of cereal seed showed a promising correlation by reducing their amount with the increasing level of soluble sugar at all the studied period (Fig. 4).
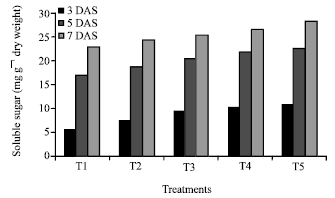 | |
| Fig. 3: | Effect of seed hardening with distilled water and different concentrations of Mg(NO3)2 on soluble sugar content of endosperm of rice var. Swarna at different study periods of germination [CD at 1% = 0.14 (3 DAS), 0.12 (5 DAS), 0.45 (7 DAS)]. *T1- Control (non-hardened); T2- distilled water hardened; T3, T4 and T5- 2, 4 and 8 mM Mg(NO3)2 hardened |
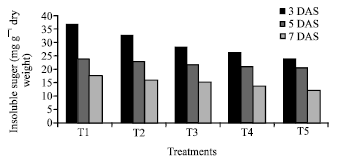 | |
| Fig. 4: | Effect of seed hardening with distilled water and different concentrations of Mg(NO3)2 on insoluble sugar content of endosperm of rice var. Swarna at different study periods of germination[CD at 1% = 2.15 (3 DAS), 0.24 (5 DAS), 0.16 (7 DAS)]. *T1- Control (non-hardened); T2- distilled water hardened; T3, T4 and T5- 2, 4 and 8 mM Mg(NO3)2 hardened |
The finding establishes that during hydro as well as halo hardening of rice seeds did not loss their internal metabolic activities of seeds. A correlative increment in the activity of amylase enzyme with reducing sugar content has also been reported by Farooq et al. (2010) in germinating rice seeds primed with CaCl2, ascorbate, and KCl. The present study therefore clearly showed that Mg(NO3)2 is a promising chemical for seed hardening which in turn improves the process of germination in rice seed. However, Alboresi et al. (2005) while working with mutants of Arabidopsis nitrate reductase genes suggested that nitrate acts as a signalling molecule and favours the germination and this signalling pathway interact either with abscisic acid or gibberellic acid biosynthesis. Bose et al. (1992) reported that the priming of maize seeds with Ca(NO3)2 (7.5 mM) and GA3 alone induced germination, length of root and shoot of seed/seedling in comparison to control. They further noted that while Ca(NO3)2+GA3 supplied to the seed in combination then the rate of enhancement was more. This may represent an indirect interpretation that nitrate ions induce germination via gibberellins. However magnesium is also here introduced with nitrate which has its own importance being the secondary macro-essential element. Logan et al. (2001) reported that magnesium improves respiration and biogenesis of mitochondria in seed cells after its influx during the process of seed soaking (a part of hardening). Bose and Mishra (1999) also reported an increment in the status of magnesium content in the seeds of brassica primed with different salts of magnesium.
In the present investigation fresh and dry weights and plant height were noted to be maximum in T4 treatment followed by T5, T3 and T2 hardened seeds. But control was inferior among all (Table 2). The number of roots of the growing seedlings was more in the plants raised from all the hardened sets. However, it is well established that root being the uptake machinery of the plant may provide better nourishment to the growing crop if it is more evolved. Therefore, an encouraging result regarding an increment in the fresh weight as well as number of roots might be influential in improving the growth of shoot in the present study. The same type of observation was made by Anaytullah and Bose (2007) while working with nitrate hardened wheat varieties (HUW-234 and HUW-468). However, close correlation ship between improved seedling growth characteristics and greater production of roots are assignable to an increased level of cell division at the post germinative phase of plant growth (Srivastava, 2002). This cell division might be the resultant of the presence of magnesium ion taken up by the seeds during soaking which stimulates the process of cell division by aiding the ATP formation (Kiss, 1979).
| Table 2: | Effects of seed hardening with distilled water and different concentrations of Mg(NO3)2 on shoot and root length (cm), root number, fresh and dry weights (g), proline content (mg g-1 dry wt.), total nitrogen content (mg g-1 dry wt.) of seedlings of rice var. Swarna at different days after sowing |
 | |
| T1: Control (non-hardened), T2: Distilled water hardened, T3, T4 and T5: 2, 4 and 8 mM Mg(NO3)2 hardened | |
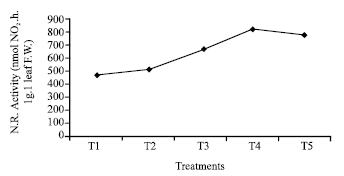 | |
| Fig. 5: | Effect of seed hardening with distilled water and different concentrations of Mg(NO3)2 on nitrate reductase activity of rice var. Swarna at 15 DAS (CD at 1% = 11.3). *T1- Control (non-hardened); T2- distilled water hardened; T3, T4 and T5- 2, 4 and 8 mM Mg(NO3)2 hardened |
The studies regarding some biochemical parameters related to stress/nitrogen metabolism like proline content, nitrogen content and nitrate reductase activity increased in the growing seedlings raised from all hardened sets in general in which 4 mM Mg(NO3)2 was found to achieve maximum level in this respect (Table 2, Fig. 5). It is well established that proline is an osmoregulating compatible solute which provides a stress ameliorating character to any crop by its presence more than threshold level (Shah and Dubey, 1997; Atreya et al., 2009). On the basis of present study it may be hypothesized that the soaking and drying of the seeds (hardening) may generate/modulate some mechanism inside the seed system which carried over to the vegetative phase of the plants life consequently that improves the stress ameliorating character in the growing plant even in the normal condition by improving the status of proline. Again, Amirjani (2010) showed that increasing NaCl concentrations lead to increase of proline content of soybean. Therefore, in the present case nitrate seed treatment also offers a novel fact towards the improvement in stress regulating capacity in rice var. Swarna. Enhanced amount of nitrogen content and increased level of nitrate reductase activity, obtained in the present study suggest that nitrate moiety must be influxed in the seed during soaking part of seed hardening process which further showed their impact in the germination time as well as vegetative growth phase. However, Bose and Pandey (2003) observed that the nitrate treated sets have more nitrogen content and nitrate reductase activity in okra cotyledons during germination. Nitric oxide functions as a major signalling molecule in plants. Both biotic and abiotic stresses can induce the synthesis of NO in plants but the biosynthetic origin of NO in plants have not yet been fully resolved (Durner and Klessig, 1999). However, study suggests that synthesis of NO depends on the activity of nitrite-NO reductase on the plasmamembrane of roots. Mitochondria also support nitrate dependent NO synthesis (Jasid et al., 2006). Hence, the present study suggests that during priming nitrate as well as magnesium both accumulated in the vacuole and their effect carried over to the vegetative phase of crop where this nitrate may induce the nitrate reductase activity. An increment in nitrate reductase activity in leaves and an improvement in root growth in seedlings raised from nitrate hardened sets may improve the chances of NO generation which in turn induced the stress ameliorating in growing crops by improving proline content. However, the effect is concentration specific in respect to Mg(NO3)2 salt.
REFERENCES
- Alboresi, A., C. Gestin, M.T. Leydecker, M. Bedu, C. Meyer and H.N. Truong, 2005. Nitrate, a signal relieving seed dormancy in Arabidopsis. Plant Cell Environ., 28: 500-512.
PubMed - Amirjani, M.R., 2010. Effect of salinity stress on growth, mineral composition, proline content, antioxidant enzymes of soybean. Am. J. Plant Physiol., 5: 350-360.
CrossRefDirect Link - Atreya, A., V. Vartak and S. Bhargava, 2009. Salt priming improves tolerance to dessication stress and to extreme salt stress in Bruguiera cylindrical. Int. J. Integrat. Biol., 6: 68-72.
Direct Link - Bailly, C., A. Benamar, F. Corbineau and D. Come, 1997. Changes in Superoxide Dismutase, Catalase and Glutathione Reductase Activities in Sunflower Seeds during Accelerated Ageing and Subsequent Priming. In: Basic and Applied Aspects of Seed Biology, Ellis, R.H., M. Black, A.J. Murdoch and T.D. Hong (Eds.). Kluwer Academic, Dordrecht, pp: 665-672.
- Basra, S.M.A., M. Farooq, R. Tabassam and N. Ahmad, 2005. Physiological and biochemical aspects of pre-sowing seed treatments in fine rice (Oryza sativa L.). Seed Sci. Technol., 33: 623-628.
CrossRefDirect Link - Bose, B. and T. Mishra, 2001. Effect of seed treatment with magnesium salts on growth and chemical attributes of mustard. Indian J. Plant Physiol., 6: 431-434.
Direct Link - Bose, B. and M.K. Pandey, 2003. Effect of nitrate pre soaking of okra (Abelmoschus esculentus L.) seeds on growth and nitrate assimilation of seedlings. Physiol. Mol. Biol. Plants, 9: 287-290.
Direct Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Durner, J. and D.F. Klessig, 1999. Nitric oxide as a signal in plants. Curr. Opin. Plant Biol., 2: 369-374.
CrossRef - Farooq, M., S.M.A. Basra, A. Wahid and N. Ahmad, 2010. Changes in nutrient-homeostasis and reserves metabolism during rice seed priming: Consequences for seedling emergence and growth. Agric. Sci. China, 9: 191-198.
CrossRefDirect Link - Farooq, M., S.M.A. Basra and K. Hafeez, 2006. Seed invigoration by osmohardening in fine and coarse rice. Seed Sci. Technol., 34: 181-187.
Direct Link - Farooq, M., S.M.A. Basra, I. Afzal and A. Khaliq, 2006. Optimization of hydropriming techniques for rice seed invigoration. Seed Sci. Technol., 34: 507-512.
CrossRefDirect Link - Ghassemi-Golezani, K., A.A. Aliloo, M. Valizadeh and M. Moghaddam, 2008. Effects of different priming techniques on seed invigoration and seedling establishment of lentil (Lens culinaris Medik). J. Food Agric. Environ., 6: 222-226.
Direct Link - Ghassemi-Golezani, K., P. Sheikhzadeh-Mosaddegh and M. Valizadeh, 2008. Effects of hydropriming duration and limited irrigation on field performance of chickpea. Res. J. Seed Sci., 1: 34-40.
CrossRefDirect Link - Harris, D., B.S. Raghuwanshi, J.S. Gangwar, S.C. Singh, K.D. Joshi, A. Rashid and P.A. Hollington, 2001. Participatory evaluation by farmers of on-farm seed priming in wheat in India, Nepal and Pakistan. Exp. Agric., 37: 403-415.
CrossRefDirect Link - Jasid, S., M. Simontacchi, C.G. Bartoli and S. Puntarulo, 2006. Chloroplasts as a nitric oxide cellular source. Effect of reactive nitrogen species on chloroplastic lipids and proteins. Plant Physiol., 142: 1246-1255.
CrossRef - Kumar, G.S., A. Rajarajan, N. Thavaprakash, C. Babu and R. Umashankar, 2008. Nitrogen use efficiency of rice (Oryza sativa) in systems of cultivation with varied n levels under 15N tracer technique Asian J. Agric. Res., 2: 37-40.
CrossRefDirect Link - De Lespinay, A., H. Lequeux, B, Lambillotte and S. Lutts, 2010. Protein synthesis is differentially required for germination in Poa pratensis and Trifolium repens in the absence or in the presence of cadmium. Plant Growth Regul., 61: 205-214.
CrossRef - Logan, D.C., A.H. Millar and L.J. Sweetlove, S.A. Hill and C.J. Leaver, 2001. Mitochondrial biogenesis during germination in maize embryos. Plant Physiol., 125: 662-672.
Direct Link - Omidi, H., 2010. Changes of proline content and activity of antioxidative enzymes in two canola genotype under drought stress. Am. J. Plant Physiol., 5: 338-349.
CrossRefDirect Link - Shah, K. and R.S. Dubey, 1997. Effect of cadmium on proline accumulation and ribonuclease activity in rice seedlings: Role of proline as a possible enzyme protectant. Biol. Plant., 40: 121-130.
CrossRef - Sharma, M.K. and B. Bose, 2006. Effect of seed hardening with nitrate salts on seedling emergence, plant growth and nitrate assimilation of wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants, 12: 173-176.
Direct Link - Srivastava, H.S., 1974. In vivo activity of nitrate reductase in maize seedling. Indian J. Biochem. Biophys., 11: 230-232.
Direct Link - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Lang, C.A., 1958. Simple micro determination of Kjeldahl nitrogen in biological materials. Annal. Chem., 30: 1692-1694.
CrossRef