
Patade Vikas Yadav
Molecular Biology and Genetic Engineering Division, Defence Institute of Bio-Energy Research (DIBER), Goraparao, P.O. Arjunpur, Haldwani, Nainital (Uttarakhand)-263 139, India
Maya Kumari
Molecular Biology and Genetic Engineering Division, Defence Institute of Bio-Energy Research (DIBER), Goraparao, P.O. Arjunpur, Haldwani, Nainital (Uttarakhand)-263 139, India
Zakwan Ahmed
Molecular Biology and Genetic Engineering Division, Defence Institute of Bio-Energy Research (DIBER), Goraparao, P.O. Arjunpur, Haldwani, Nainital (Uttarakhand)-263 139, India
Research Journal of Seed Science
Year: 2011 | Volume: 4 | Issue: 3 | Page No.: 125-136







ABSTRACT
Seed priming is a simple pre-germination strategy to improve seed performance and alleviate the negative effects associated with stress exposure. Seed priming technique was used in capsicum (Capsicum annuum L. cv. California Wonder) to improve seed germination and tolerance on subsequent stress exposure. Capsicum seeds were treated with various priming agents for 24 h with moderate shaking. The effect of priming was assessed on germination and survival on subsequent exposure to salt (NaCl, 200 mM) or cold (4°C) stress for 10 days. Seed priming treatments significantly (p≤0.05) affected rate as well as the germination percentage under control, non-stressed condition. Seed priming with normal water (NW), warm water (40°C; WW) or NaCl (50 mM; NA) significantly (p≤0.05) increased the germination percentage by 7 to 17% as compared to the control. On cold exposure, the highest rate of seed germination and opening of cotyledonary leaves recorded in terms of Timson’s index (94 and 56.9 respectively) was observed in thiourea (TU, 1.3 mM) primed seeds. The germination percentage was also significantly (p≤0.05) higher in primed seeds (97.5 to 100%) as compared to the control (85%). Only the seedlings grown from TU, hydrogen peroxide (HP, 1.5 mM) or ABA (AB, 100 μM) primed seeds tolerated the cold or salt stress exposure of 10 days with 100% survival. However, the control seedlings could not survive the salt or cold stress exposure. The plants grown from primed seeds under controlled conditions flowered earlier than the control and produced more number of fruits than the control. Thus, the results suggest chemical seed priming as an efficient approach for cold or salt stress tolerance in capsicum.
PDF Abstract XML References Citation
Received: April 29, 2011;
Accepted: May 31, 2011;
Published: July 09, 2011
How to cite this article
Patade Vikas Yadav, Maya Kumari and Zakwan Ahmed, 2011. Seed Priming Mediated Germination Improvement and Tolerance to Subsequent Exposure to Cold and Salt Stress in Capsicum. Research Journal of Seed Science, 4: 125-136.
URL: https://scialert.net/abstract/?doi=rjss.2011.125.136
URL: https://scialert.net/abstract/?doi=rjss.2011.125.136
INTRODUCTION
Capsicum also known as bell or sweet pepper (Capsicum annum L.) originated in Central and South America is an important solanaceous vegetable crop worldwide. Capsicum grows better in warm climate and can be grown year round in frost-free conditions. Cold condition (below 15°C) lowers fruit setting due to poor pollination and may leads to fruit splitting (Eshbaugh, 1993). It has also been reported that capsicum seed germination is slow and non-uniform under normal as well as abiotic stress conditions including salinity (Chartzoulakis and Klapaki, 2000; Demir and Okcu, 2004; Khan et al., 2009).
Improvement in seed germination is an active area of research and efforts are being made to improve seed germination in various economically important plants as evident by earlier reports (Demir and Mavi, 2008; Kaur et al., 2009; Gebreselassie et al., 2010; Alderete-Chavez et al., 2011; Asomaning et al., 2011; Suleman et al., 2011). Priming, a prior encounter with a particular type of stress is known to endow plants with greater tolerance to subsequent stress exposure of the same or different kind (Parra et al., 2007; Wahid et al., 2008; Patanea et al., 2009). Seed priming is a pre-sowing strategy for influencing the seedling development at a later stage by modulating pre-germination metabolic activity prior to protrusion of the radicle (Taylor and Harman, 1990). During priming, the seeds are partially hydrated with water (hydro priming), or various chemical solutions like polyethylene glycol (osmopriming) or salts like CaCl2, CaSO4, NaCl (halopriming), in such a way that the pre-germination metabolic activities starts however, radicle protrusion is prevented, followed by drying of seeds to the original moisture level (McDonald, 2000). Although the phenomenon has been known for decades, the causes underlying better performance of the primed plants are not well understood. Over the past decades, there has been increasing evidence that primed plants exhibit activation of cellular defense responses, due to which they can better tolerate subsequent biotic or abiotic stresses in the field (Beckers and Conrath, 2007).
Seed priming techniques have been used to accelerate synchronized seed germination, improve seedling establishment, stimulate vegetative growth and crop yield in many field crops like wheat (Iqbal and Ashraf, 2007), chickpea (Kaur et al., 2002) and cotton (Casenave and Toselli, 2007). It has been reported that the plantlets from primed seeds emerge faster, grow more vigorously and perform better under sub-optimal conditions (Khalil et al., 2001; Sivritepe et al., 2003). Earlier reports on seed priming in field crops like wheat (Ghiyasi et al., 2008), lentil (Ghassemi-Golezani et al., 2008a), maize (Foti et al., 2008), sugarcane (Patade et al., 2009) and cucumber (Ghassemi-Golezani and Esmaeilpour, 2008) also indicated improved growth and stress tolerance of the primed seedlings. Although effect of seed priming in other crops is documented, no reports are available on potential of various seed priming treatments and responses of the capsicum plants on subsequent exposure to cold or salt stress. Therefore, the present investigation was carried out to assess the ameliorative potential of various chemical priming treatments on germination in capsicum cv. California Wonder as well the performance of seedlings grown from primed seeds on subsequent salt or cold stress exposure.
MATERIALS AND METHODS
Seeds of capsicum cv. California Wonder were obtained from Indian Agriculture Research Institute, New Delhi, Field Station, Katrain, India.
Chemical seed priming: The laboratory experiments on seed priming were conducted from July 2010 to January 2011. The seeds of capsicum cv. California Wonder, based on the earlier reports in other crops, were primed with various chemical-priming agents viz. NaCl (NA, 50 mM); CaCl2 (CA, 50 mM); thiourea (TU, 1.3 mM); hydrogen peroxide (HP, 1.5 mM) and abscisic acid, ABA (AB, 100 μM) along with soaked in water (NW) and unsoaked (US) controls. In addition, seeds were primed by soaking in warm water (WW; 40°C). The seeds were treated by soaking with shaking (60 rpm) in the priming solutions at 25±2°C for 24 h. At the end of the treatment, the solutions were decanted and seeds were rinsed twice with double distilled water to remove the traces of chemicals. Then the seeds were blot dried with tissue paper and allowed to dry in shed. The seeds (10 per plate) were then sown on filter paper placed on absorbent cotton in a glass petri plate. The absorbent cotton was moistened and kept wet with double distilled water. Data on seed germination as well as opening of cotyledonary leaves was recorded twice a day. The rate of germination was estimated in terms of Timson’s index of germination velocity as, Timson’s index = Σ(G/t); where G is germination percentage; t = total period of germination (days) (Patade et al., 2009). Similarly, the rate of opening of cotyledonary leaves was calculated.
Effect of chemical seed priming on germination and survival of seedlings in response to cold stress: The potential of primed seeds to tolerate cold stress was studied at germination stage. The primed seeds (10 per plate) were sown as explained earlier and then exposed to cold stress (15°C) until completion of germination. The rate of germination and opening of cotyledonary leaves was estimated in terms of Timson’s index (Patade et al., 2009). Also the seedlings grown from primed seeds (eight per treatment) at 60 days after sowing-DAS were exposed to cold (4°C) stress for 10 days in a cold chamber equipped with diffused cool florescent light, manually adjusted for 16 h light and 8 h dark period. The ability of primed plants to tolerate cold stress was recorded in terms of survival rate.
Salt tolerance in capsicum at germination stage: Capsicum seeds (10 per plate) were sown on filter paper placed on absorbent cotton in a glass petri plate. The salt concentrations chosen were 0, 25, 50, 100, 150, 200, 250 or 300 mM of NaCl. The cotton was kept wet with respective salt solution. Data on seed germination in terms of radicle protrusion as well as opening of cotyledonary leaves was recorded twice a day. The rate of germination and opening of cotyledonary leaves was estimated in terms of Timson’s index of germination velocity as explained earlier.
As the seeds kept for germination at higher salt concentrations viz. 250 and 300 mM NaCl failed to germinate even after long period of exposure, the seed viability was tested by removing the salt stress and allowing the seeds to germinate as in control condition.
Effect of chemical seed priming on survival of seedlings in response to salt stress: The seedlings grown from the earlier primed seeds (eight per treatment) at 60 DAS were exposed to salt, NaCl (200 mM) stress for 10 days at 16 h light and 8 h dark period. The ability of primed plants to tolerate salt stress was recorded in terms of seedling survival.
Optimization of seed priming treatment with TU: Capsicum seed priming with TU for 24 h improved seed germination in cold stress regime as well as imparted tolerance on exposure to cold stress by improving seedling survival. However, in absence of stress conditions, TU priming treatment inhibited seed germination rate as well as germination percentage. Therefore, in order to optimize the seed priming treatment, priming duration was reduced to 16 h and various TU concentrations (0, 0.33, 0.65, 1.3, 2.6 or 5.2 mM) were tried. The primed seeds were sown in glass petri plates as explained earlier. The rate and percentage of seed germination as well as the opening of cotyledonary leaves was calculated.
Performance of the seedlings from primed seeds under controlled condition: The seedlings from primed and non-primed seeds were transplanted in partially controlled (temperature 25±2°C; 16 h photoperiod and 8 h dark period) green house in pots filled with potting mixture (soil: FYM: sand, 1:1:1). Observations on plant height (cm) were recorded. The repeatability of the results on seed priming and additional data on economic traits (fruits per plant, fruit weight etc.) under controlled conditions, is being studied in the current season (From March 2011).
Statistical analysis: The treatments and controls of the experiments were replicated six times. The program CropStat for Windows (7.2.2007.2 module), developed by the Biometrics Unit, IRRI, Philippines was used for Analysis of Variance (ANOVA) of experiments laid out in Completely Randomized Design (CRD). The treatment means were compared by using the Duncan’s Multiple Range Test (DMRT) at a significance level of p≤0.05.
RESULTS
Effect of chemical seed priming on seed germination: Effect of chemical seed priming was assessed on germination and opening of cotyledonary leaves. Hydro priming with normal water (NW) or warm water (WW) improved the radicle protrusion significantly (p = 0.05) as compared to the control treatment (Fig. 1). Halo priming with salt (NA) also improved the rate of seed germination. The rest of the treatments inhibited rate of radicle protrusion significantly. The seed treatments with warm water (WW) significantly increased the germination percentage, whereas the calcium chloride (CA), thiourea (TU), or hydrogen peroxide (HP) treatments significantly reduced the germination. Opening of cotyledonary leaves was also greatly affected by various seed priming treatments (Fig. 2). Rate of opening of cotyledonary leaves was significantly higher in hydropriming with normal water (NW) or warm water (WW) as compared to the control (US). Whereas, seed priming with calcium chloride (CA), thiourea (TU), hydrogen peroxide (HP) or abscisic acid (ABA) significantly reduced the rate of cotyledonary leaves opening . The percentage of opening of cotyledonary leaves was significantly higher in NW, WW or NA treatments however the priming treatment with CA, HP, TU or AB significantly inhibited the opening of cotyledonary leaves.
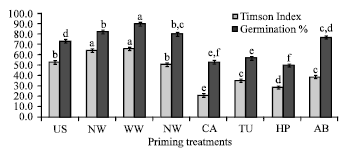 | |
| Fig. 1: | Effect of various chemical priming treatments on germination in terms of radicle emergence in capsicum cv. California Wonder. The error bars indicate SE. Means indicated by different alphabets in each column were significantly different at p≤0.05, according to DMRT. |
 | |
| Fig. 2: | Effect of various chemical priming treatments on opening of cotyledonary leaves in capsicum cv. California Wonder. The error bars indicate SE. Means indicated by different alphabets in each column were significantly different at p≤0.05, according to DMRT. |
Effect of chemical seed priming on germination and survival of seedlings in response to cold stress: Seed priming treatments affected radicle protrusion in response to cold stress treatment. The highest rate of seed germination in term of radicle protrusion was observed in TU or HP primed seeds followed by NA or CA treatments (Table 1). The rate of germination was higher in other treatments, than the control. The germination percentage was also significantly (p≤0.05) higher in primed seeds as compared to the control.
Similar to the radicle protrusion, opening of cotyledonary leaves was also induced by various seed priming treatments (Table 1). Significantly the highest rate of opening of cotyledonary leaves was observed in TU treatment. The WW or HP primed seeds exhibited significantly faster opening of cotyledonary leaves than the control. Similarly, the percentage of opening of cotyledonary leaves was the highest in TU treatment. NW and WW treatments also exhibited significantly higher percentage of opening of cotyledonary leaves as compared to the control.
The seedlings grown from primed and control seeds on exposure to 10 days cold stress exhibited differential survival rate. The seedlings grown from control (US), NW or WW treated seeds could not survive the cold stress and all the seedlings were observed drooped. Whereas, 66% of the seedlings grown from NA or CA primed seeds could survive the cold exposure of 10 days. The seedlings grown from TU, HP or AB primed seeds tolerated the cold stress exposure and exhibited 100% survival.
Salt tolerance limit of capsicum cv. California wonder: Salt concentrations affected rate as well as germination percentage of capsicum cv. California Wonder seeds (Table 2). The salt concentration of 100 mM and higher, reduced the rate of seed germination in terms of radicle protrusion significantly (p≤0.05). However, the lower NaCl concentration (25 mM) significantly stimulated rate of radicle protrusion. The rate of seed germination in terms of radicle protrusion was similar to that of control in 50 mM NaCl treatment however, the higher salt concentrations reduced not only the rate of germination but also the germination percentage. The salt concentration of 300 mM NaCl completely inhibited the radicle protrusion (Table 2).
The opening of cotyledonary leaves was also affected by various salt treatments. Radicle protrusion was observed in response to 250 mM NaCl or lower salt concentrations however, the cotyledonary leaves failed to open in salt treatment of more than 150 mM NaCl (Table 2).
| Table 1: | Effect of various chemical priming treatments on seed germination and opening of cotyledonary leaves on exposure to cold (4°C) stress at germination stage in capsicum cv. California Wonder |
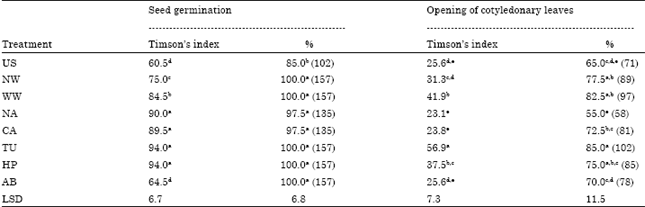 | |
| The values are means of six observations. Means superscribed by different alphabets in each column were significantly different at p≤0.05, according to DMRT. Values in parenthesis are arcsin angular transformed values | |
| Table 2: | Effect of salt-NaCl concentrations on capsicum cv. California Wonder seed germination and opening of cotyledonary leaves |
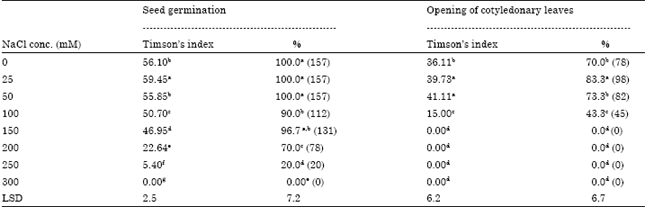 | |
| The values are means of six observations. Means superscribed by different alphabets in each column were significantly different at p≤0.05, according to DMRT. Values in parenthesis are arcsin angular transformed values | |
| Table 3: | Effect of salt-NaCl stress removal on capsicum cv. California Wonder seed germination and opening of cotyledonary leaves |
 | |
| The values are means of six observations. Means superscribed by different alphabets in each column were significantly different at p≤0.05, according to DMRT. Values in parenthesis are arcsin angular transformed values | |
As observed in case of radicle protrusion, the opening of cotyledonary leaves was also induced by the lower (25 mM NaCl) salt concentration. As the NaCl concentration greater than 150 mM inhibited completely the opening of cotyledonary leaves hence, may be used for screening of salt tolerance in capsicum cultivars/genotypes.
In order to check whether seeds have lost the viability after a long exposure to higher salt concentrations of 250 or 300 mM NaCl, the salt stress was removed and seeds were allowed to germinate as in control condition. After removal of salt stress, radicle protrusion as well as opening of cotyledonary leaves was greatly increased (Table 3) as compared to that observed during stress exposure (Table 2). Even after removal of the stress, the germination was higher in seeds earlier exposed to the lower salt concentration of 150 mM NaCl, indicating the inhibition of germination due to long-term salt stress exposure. Thus, though the higher salt concentrations inhibited seed germination, seed viability was not affected.
Effect of chemical seed priming on survival of seedlings in response to salt stress: The seedlings grown from primed and control seeds at 60 DAS on exposure to 10 days salt (NaCl, 200 mM) stress exhibited differential survival rate. The seedlings grown from US or NW treated seeds did not survive the salt stress. Whereas, the 25% of seedlings from WW or NA primed seeds survived the salt stress exposure of 10 days. The seedlings grown from the rest of the CA, TU, HP or AB priming treatments tolerated the salt stress exposure and exhibited 100% survival.
Optimization of seed priming treatment with TU: Germination percentage recorded in terms of radicle protrusion was significantly improved in capsicum seeds primed with various concentrations of TU or hydropriming for 16 h (Table 4) as compared to the unsoaked control (US). In addition the rate of seed germination in terms of Timson’s index was significantly higher in hydro and TU (1.3, 2.6 or 5.2 mM) primed seeds than the unsoaked control. Besides radicle protrusion, the seed priming with various TU concentrations for 16h also significantly improved the speed of opening of cotyledonary leaves and total number of cotyledonary leaves opened (Table 4). The seed germination inhibition which was observed in 1.3 mM TU primed seeds for 24 h (Fig. 1, 2), was not evident even with the higher TU concentrations (2.6, 5.2 mM), when treated for 16 h. Thus, seed priming with TU for 16 h may be considered as optimal treatment period.
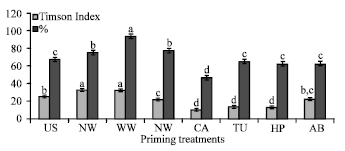
Performance of the primed seedlings under controlled conditions: The seedlings grown from primed seeds after transplantation in controlled green house condition established normally. The vegetative growth in terms of plant height of the plants grown from primed seeds was superior to the control. Plant height (cm) recorded 60 Days After Transplanting (DAT) was significantly (p≤0.05) higher in priming treatment and the lowest in control treatment (Fig. 3). Among the priming treatments, plants grown from NA, HP and AB primed seeds were significantly taller than all the other treatments. In addition, the plants from primed seeds showed earlier initiation of flowering and fruiting than the control. Further, the performance of plants grown from primed seeds in terms of economic yield and quality as compared to the control is being studied.
| Table 4: | Effect of TU concentrations on capsicum cv. California Wonder seed germination and opening of cotyledonary leaves the values are means of six observations |
 | |
| Means super scribed by different alphabets in each column were significantly different at p≤0.05, according to DMRT. values in parenthesis are arcsin angular transformed values | |
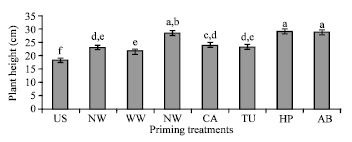 | |
| Fig. 3: | Effect of priming on plant height (cm) in capsicum cv California Wonder recorded 60 days after transplanting in partially controlled conditions. The column bars are means of six observations. The error bars indicate SE. The means indicated by different alphabets in each column bar were significantly different at p≤0.05, according to DMRT |
DISCUSSION
In the present investigation, salt concentrations affected percentage and rate of germination as well as the opening of cotyledonary leaves in capsicum cv. California Wonder. The 100 mM or higher salt concentrations reduced the rate of radicle protrusion significantly. However, the lower NaCl concentration of 25 mM significantly stimulated rate of radicle protrusion as well as opening of cotyledonary leaves. The results are in agreement with the earlier study by Chartzoulakis and Klapaki (2000) who reported the significant reduction in the germination as well as growth of two bell-pepper hybrids on exposure to 100 and 150 mM NaCl. As the salt concentration of 150 mM and higher inhibited completely the opening of cotyledonary leaves therefore, 200 mM NaCl was used to study the potential of priming to impart salt tolerance in capsicum. Bell pepper being sensitive, as revealed in the present and earlier studies, salinity affect rapid, synchronous and complete germination which is prerequisite for successful transplant production and stand establishment.
Seed germination and seedling establishment are critical stages in the life cycle of a plant, especially under adverse environmental conditions. Chemical seed priming is one of the pre-germination strategy to overcome the negative effects associated with the stresses (Patade et al., 2009). In the present study, effect of various chemical seed priming agents was assessed on germination and opening of cotyledonary leaves. Under control conditions, chemical seed priming with NA significantly increased rate of radicle protrusion, whereas hydro priming (with normal or warm water) improved the radicle protrusion significantly as compared to the control. In agreement with the results, earlier reports (Parra et al., 2007; Ghassemi-Golezani et al., 2008b; Wahid et al., 2008; Patanea et al., 2009) have shown positive effect of priming in relation to enhanced seed performance, faster and synchronized seed germination and ability to tolerate subsequent stress episodes, resulting in greater survival. Early initiation of metabolic activities and thus reserve breakdown and mobilization, might be the reason for faster germination in primed seeds. Contrary to the results obtained in the present study, Abebe and Modi (2009) reported negative effects of hydropriming on seed germination in dry bean wherein, better performance was observed in control over the priming. However, the priming treatments in the present investigation like thiourea (TU) under the control condition, inhibited rate of radicle protrusion significantly as well as opening of cotyledonary leaves. This could be due to the higher inhibitory concentrations of these priming agents or longer priming treatment. Therefore, in order to optimize the seed priming treatment, short duration seed priming for 16h with lower and higher TU concentrations was performed. Interestingly, even higher TU concentrations of 2.6 and 5.2 mM induced the rate of seed germination and germination percentage. Moradi and Younesi (2009) reported positive effect of 12 and 24 h osmo or hydropriming on the percentage and mean emergence time under sub-optimal temperature of 15°C in sorghum. However, the positive priming effect was not observed in 36 h primed seed and failed to improve emergence percentage and time. Under cold stress condition, seed priming treatments in the present study, improved radicle protrusion as well as opening of cotyledonary leaves. The highest rate of seed germination in term of radicle protrusion was observed in TU or HP treated seeds, followed by NA or CA treatments. The germination percentage was also significantly higher in primed seeds as compared to the control. In addition, in response to the lower salt concentration, the various seed priming treatments enhanced the rate of radicle protrusion in terms of Timson’s index. The AB primed seeds exhibited faster rate of radicle protrusion followed by TU. The opening of cotyledonary leaves was the highest in TU primed seeds followed by NA primed seeds. TU has been shown to be ameliorative and highly effective in promoting germination under abiotic stress condition (Khan and Ungar, 2001; Siddiqui et al., 2006; Srivastava et al., 2008). The results of the present investigations indicated that seed priming could be an effective pre-germination practice for uniform and faster germination of capsicum. The results are in agreement with the earlier reports on seed priming in field crops like sugarcane (Patade et al., 2009), wheat (Ghiyasi et al., 2008), lentil (Ghassemi-Golezani et al., 2008a), maize (Foti et al., 2008) and cucumber (Ghassemi-Golezani and Esmaeilpour, 2008).
After establishing the positive effect of some of the seed priming treatments on synchronous and faster germination, the potential of priming on imparting tolerance to cold or salt stress was assessed in comparison to non-primed seeds and seedling. The seedlings grown from primed and control seeds on exposure to 10 days cold or salt stress exhibited differential survival rate. The seedlings from US or NW treatments did not survive either salt or cold stress. Only seedlings grown from TU, HP or ABA (AB, 100 μM) primed seeds tolerated the cold or salt stress with 100% survival. Whereas, seedlings grown from CA primed seeds survived salt, but not the cold stress exposure. Priming mediated abiotic stress tolerance is earlier reported in hot pepper and other crops. Khan et al. (2009) reported salt priming induced salinity tolerance of hot pepper at seedling stage, wherein seed priming improved significantly the germination percentage and index, vigour index, plumule and radicle length and dry weight of seedlings as compared to the non-primed control. In another study in sugarcane, Patade et al. (2009) reported improvement in the percent and rate of sets germination and growth performance and, lower salt- and dehydration-induced leaf senescence when subjected to 15 days iso-osmotic salt or polyethylene glycol induced osmotic stress.
Although the phenomenon has been known for decades, the causes underlying better performance of the primed plants are not well understood. Over the past decades, there has been increasing evidence that primed plants exhibit activation of cellular defense responses, due to which they can better tolerate subsequent biotic or abiotic stresses in the field. Improved stress tolerance of primed plants is thought to arise from the activation of cellular defense responses (Beckers and Conrath, 2007). This has been substantiated by reports on better osmotic adjustment in primed plants of canola (Rozbeh and Farzad, 2006) and a better antioxidant system in primed plants of wheat (Afzal et al., 2006) on exposure to stress. Conrath et al. (2006) proposed that priming could involve accumulation of signaling proteins or transcription factors. The model would mediate more transient or short-term effects but our data revealed that the priming effects could be observed for a longer period (more than two months). The results of the present study support the earlier finding that epigenetic changes would enable stress imprints to be left in the plant for long term (Bruce et al., 2007).
Thus, based on the finding of the present investigation, we propose seed priming as a tool to impart abiotic stress tolerance in capsicum. Our further studies are underway to investigate the mechanisms underlying the priming-induced stress tolerance and whether the beneficial effects of priming persist beyond the seedling stage and, its nature (epigenetic and hereditary). The seed priming experiment is being repeated and the seedlings raised from primed seeds are being grown in controlled conditions to confirm the results and study their performance in terms of economic yield.
CONCLUSIONS
The chemical priming treatments improved germination percentage as well as rate of seed germination in capsicum under normal as well as the stressed conditions. Further, chemical seed priming treatments imparted tolerance to subsequent cold or salt stress exposure at later growth stages. Among the treatments, seed priming with TU was more effective in imparting cold as well as salt stress tolerance. Preliminary analysis indicated better performance of plants grown from primed seeds than the control. Thus, the results of the present study suggest priming as a simple and environment friendly approach for imparting the stress tolerance in capsicum. However, it needs further exploration under large-scale trial under natural field conditions to gain more insights towards the yield components.
REFERENCES
- Abebe, A.T. and A.T. Modi, 2009. Hydro-priming in dry bean (Phaseolus vulgaris L.). Res. J. Seed Sci., 2: 23-31.
CrossRefDirect Link - Afzal, I., S.M.A. Basra, A. Hameed and M. Farooq, 2006. Physiological enhancements for alleviation of salt stress in wheat. Pak. J. Bot., 385: 1649-1659.
Direct Link - Alderete-Chavez, A., N. de la Cruz-Landero, J.J. Guerra-Santos, E. Guevara and R. Gelabert et al., 2011. Promotion of germination of Bauhinia divaricata L. seeds by effects of chemical scarification. Res. J. Seed Sci., 4: 51-57.
CrossRefDirect Link - Beckers, G.J.M. and U. Conrath, 2007. Priming for stress resistance: From the lab to the field. Curr. Opin. Plant Biol., 10: 425-431.
CrossRefPubMedDirect Link - Bruce, T.J.A., M.C. Matthes, J.A. Napier and J.A. Pickett, 2007. Stressful memories of plants: Evidence and possible mechanisms. Plant Sci., 173: 603-608.
CrossRefDirect Link - Casenave, E.C. and M.E. Toselli, 2007. Hydropriming as a pre-treatment for cotton germination under thermal and water stress conditions. Seed Sci. Technol., 35: 88-98.
Direct Link - Chartzoulakis, K. and G. Klapaki, 2000. Response of two greenhouse pepper hybrids to NaCl salinity during different growth stages. Sci. Hortic., 86: 247-260.
CrossRefDirect Link - Conrath, U., G.J.M. Beckers, V. Flors, P. García-Agustín and G. Jakab et al., 2006. Priming: Getting ready for battle. Mol. Plant Microbe Interact., 19: 1062-1071.
CrossRefPubMedDirect Link - Demir, I. and G. Okcu, 2004. Aerated hydration treatment for improved germination and seedling growth in aubergine (Solarium melongena) and pepper (Capsicum annuum). Ann. Applied Biol., 144: 121-123.
CrossRefDirect Link - Demir, I. and K. Mavi, 2008. Seed vigour evaluation of cucumber (Cucumis sativus L.) seeds in relation to seedling emergence. Res. J. Seed Sci., 1: 19-25.
CrossRefDirect Link - Foti, R., K. Abureni, A. Tigere, J. Gotosa and J. Gere, 2008. The efficacy of different seed priming osmotica on the establishment of maize (Zea mays L.) caryopses. J. Arid Environ., 72: 1127-1130.
CrossRefDirect Link - Gebreselassie, W., A. Mohammed and A. Netsere, 2010. Pre-sowing treatment of coffee (Coffea arabica L.) seeds to enhance emergence and subsequent growth of seedlings. Res. J. Seed Sci., 3: 218-226.
CrossRefDirect Link - Ghassemi-Golezani, K., AA. Aliloo, M. Valizadeh and M. Moghaddam, 2008. Effects of hydro and osmo-priming on seed germination and field emergence of lentil (Lens culinaris Medik.). Not. Bot. Hort. Agrobot. Cluj-Napoca, 36: 29-33.
Direct Link - Ghassemi-Golezani, K. and B. Esmaeilpour, 2008. The effect of salt priming on the performance of differentially matured cucumber Cucumis sativus seeds. Not. Bot. Hort. Agrobot. Cluj-Napoca, 36: 67-70.
Direct Link - Ghassemi-Golezani, K., P. Sheikhzadeh-Mosaddegh and M. Valizadeh, 2008. Effects of hydropriming duration and limited irrigation on field performance of chickpea. Res. J. Seed Sci., 1: 34-40.
CrossRefDirect Link - Ghiyasi, M., A. Abbasi Seyahjani, T. Mehdi, A. Reza and S. Hojat, 2008. Effect of osmopriming with polyethylene glycol (8000) on germination and seedling growth of wheat (Triticum aestivum L.) seeds under salt stress. Res. J. Biol. Sci., 3: 1249-1251.
Direct Link - Iqbal, M. and M. Ashraf, 2007. Seed treatment with auxins modulates growth and ion partitioning in salt-stressed wheat plants. J. Integr. Plant Biol., 49: 1003-1015.
CrossRefDirect Link - Kaur, R., S. Arora and A.K. Thukral, 2009. Enhancing seed germination of Chlorophytyum borivilianum Sant. Et fernand. with pgrs, steroidal hormones and zinc. Res. J. Seed Sci., 2: 32-39.
CrossRefDirect Link - Khan, H.A., C.M. Ayub, M.A. Pervez, R.M. Bilal, M.A. Shahid and K. Ziaf, 2009. Effect of seed priming with NaCl on salinity tolerance of hot pepper (Capsicum annuum L.) at seedling stage. Soil Environ., 28: 81-87.
Direct Link - Moradi, A. and O. Younesi, 2009. Effects of osmo and hydro-priming on seed parameters of grain sorghum. Aust. J. Basic Applied Sci., 3: 1696-1700.
Direct Link - Parra, M., A. Albacete, C. Martinez-Andujar and F. Perez-Alfocea, 2007. Increasing plant vigour and tomato fruit yield under salinity by inducing plant adaptation at the earliest seedling stage. Environ. Exp. Bot., 60: 77-85.
CrossRef - Patade, V.Y., S. Bhargava and P. Suprasanna, 2009. Halopriming imparts tolerance to salt and PEG induced drought stress in sugarcane. Agric. Ecosyst. Environ., 134: 24-28.
CrossRefDirect Link - Patanea, C., V. Cavallaro and S.L. Cosentino, 2009. Germination and radicle growth in unprimed and primed seeds of sweet sorghum as affected by reduced water potential in NaCl at different temperatures. Indus. Crops Prod., 30: 1-8.
CrossRef - Khalil, S.K., J.G. Mexal and L.W. Murray, 2001. Germination of soybean seed primed in aerated solution of polyethylene glycol (8000). J. Biol. Sci., 1: 105-107.
CrossRefDirect Link - Sivritepe, N., H.O. Sivritepe and A. Eris, 2003. The effects of NaCl priming on salt tolerance in melon seedlings grown under saline conditions. Sci. Hortic., 97: 229-237.
CrossRefDirect Link - Suleman, M.K., N.R. Bhatt, S. Jacob and R.R. Thomas, 2011. Germination studies in Ochradenus baccatus delile, Peganum harmala L. and Gynandris sisrinchium parl. Res. J. Seed Sci., 4: 58-63.
CrossRefDirect Link - Taylor, A.G. and G.E. Harman, 1990. Concepts and technologies of selected seed treatments. Annu. Rev. Phytopathol., 28: 321-339.
CrossRefDirect Link - Wahid, A., A. Noreen, S.M.A. Basra, S. Gelani and M. Farooq, 2008. Priming-induced metabolic changes in sunflower (Helianthus annuus) achenes improve germination and seedling growth. Bot. Stud., 49: 343-350.
Direct Link - Kaur, S., A.K. Gupta and N. Kaur, 2002. Effect of osmo- and hydropriming of chickpea seeds on seedling growth and carbohydrate metabolism under water deficit stress. Plant Growth Regul., 37: 17-22.
CrossRefDirect Link
RAJAT AWASTHI Reply
thank you sir