
G. Kanmegne
Department of Plant Biology, Faculty of Science, University of Dschang, P.O. Box 67 Dschang, Cameroon
H.D. Mbouobda
Department of Biology, Higher Teacher Training College, Annex Bambili (ENSAB) University of Yaounde I, P.O. Box 39, Bamenda, Cameroon
B. Temfack
Department of Plant Biology, Faculty of Science, University of Dschang, P.O. Box 67 Dschang, Cameroon
E.K. Koffi
National Center for Agronomic Research, Adiopodoume, KM 17, Route de Dabou 01 P.O. Box 1740 Abidjan, Cote d `Ivoire University of Yaounde I, P.O. Box 47 Yaounde, Cameroon
D.N. Omokolo
Laboratory of Plant Physiology and Biochemistry, Higher Teacher Training College, University of Yaounde I, P.O. Box 47 Yaounde, Cameroon
Research Journal of Seed Science
Year: 2010 | Volume: 3 | Issue: 2 | Page No.: 82-92







ABSTRACT
The aim of the present study was to describe the variation that may exist among G. kola seeds collections found in Cameroon and to determine possible established correlations between one or many morpho-biochemical parameters of seeds and germination capacity. Twenty seven accessions of G. kola seeds originating from eleven locations in Cameroon were analyzed based on the variation of their morphological (weight, length and diameter), biochemical (phenols, soluble carbohydrates, proteins and amino acids contents), germination index and germination velocity. Results revealed a significant variation in weight, size, metabolites contents, germination index and germination velocity among the different accessions of seeds. These parameters also varied within samples from the same ecological zone. Significant (p<0.01) and negative correlation were revealed between weight and amino acids content of seeds (r = -0.402) as well as between the length of the seeds and their carbohydrates content (r = -0.479). A significant (p<0.05) and negative correlation was also found between proteins content and germination index of seed (r = -0.442) in one hand and between proteins content and germination velocity (r = -0.364) in another hand. The dendogram generated by cluster classification put together in the same group accessions which performed best germination and members of this group were also characterized by low proteins content. These results indicated that proteins content could serve as a criterion of selection for germination capacity in G. kola seeds. The influence of seed proteins in dormancy and germination is discussed.
PDF Abstract XML References Citation
How to cite this article
G. Kanmegne, H.D. Mbouobda, B. Temfack, E.K. Koffi and D.N. Omokolo, 2010. Impact of Biochemical and Morphological Variations on Germination Traits in Garcinia kola Heckel Seeds Collected from Cameroon. Research Journal of Seed Science, 3: 82-92.
URL: https://scialert.net/abstract/?doi=rjss.2010.82.92
URL: https://scialert.net/abstract/?doi=rjss.2010.82.92
INTRODUCTION
Garcinia kola (Heckel) of Guttiferae family is a medium size tree growing up to 12 m high in 8 years in moist forest throughout West and Central Africa (Isawumi, 1993). In Cameroon, the species is found in lowland forests of the Southern part of the country, including Centre, East, Northwest, South, Southwest and West Regions. It is an economic and highly valued tree, used extensively in African traditional medicine. Extracts of various parts of the plant are used for the treatment of laryngitis, mouth infections, cough, heart burn, liver disorder, chest colds hoarseness and others inflammatory diseases (Iwu, 1982). The seed, commonly called bitter kola because of its bitter taste, is also used in the treatment of bronchitis, throat troubles, post partum haemorrhage, urinary tract infections and emesis (Irvine, 1961). The antimicrobial activity of this plant is attributed to the benzophenones and flavanones (Iwu et al., 1987). Garcinia kola seed is one of the many non-timber forest products that are of socio-economic importance. Its commercialisation in both domestic and national markets raises the standard of living of those involved in its trading activities, in rural and urban centres (Adebisi, 2004). In spite of great demand for G. kola seeds, its cultivation is not popular owing to the difficulty in seed germination (Adebisi, 2004) although G. kola is one of the useful indigenous trees prioritised by farmers in West and Central Africa (Anegbeh et al., 2006) and the species is now close to commercial extinction (IUCN, 2004). There have been some attempts to overcome seed dormancy and enhance germination in some Nigerian, Cameroonian, Ivorian and Ghanaian collections (Emmanuel and Roy, 2001; Agyili et al., 2007; Kanmegne and Omokolo, 2008), but the high diversity of results makes it difficult to prescribe a standard procedure for enhancing its germination. More, a significant tree to tree variation in G. kola seeds responses to dormancy-breaking treatments has been reported and suspected to be due to the existence within the species, of a variability which therefore needs to be described and measured if it is to be effectively incorporated into breeding strategies and management of plant genetic resources (Kanmegne and Omokolo, 2008). Although, some studies have shown the extent of tree to tree variation in morphological and biochemical fruit and seed traits of others indigenous fruit tree species as Irvingia gabonensis (Leakey et al., 2000; Atangana et al., 2002) Canarium schweinfurthii (Kapchie et al., 2002) and Dacryodes edulis (Leakey et al., 2003), the genetic variation in G. kola remains undocumented. The establishment of correlation between germination traits and morphological/biochemical traits would contribute not only to a better understanding of the phenomenon of seed dormancy in the species, but also to an efficient orientation of research aimed at seed germination improvement. Such studies have formerly been carried out with many other plant species as groundnuts (Carvalho et al., 1981), Irish potato (Bhatt et al., 1989), Triticum aestivum (Hampton, 1981), Prunella vulgaris (Win, 1985), Balanites aegyptiaca (Azo et al., 2005) and Cicer arietinum (Emenky and Khalaf, 2008) for which correlations between seed size and germination have been reported. Also, in many plant species, there are many reports attributing the wide variation in germination traits to difference in some metabolites contents of various seed lots (Bewley and Black, 1978; Malik and Shamet, 2009). The aim of the present study, therefore, was to (1) describe the variation in G. kola seeds based on morphological (size and weight), biochemical (phenols, proteins, amino acids and carbohydrates contents) and germination traits and (2) determine possible established correlations between one or many morpho-biochemical parameters of seeds and germination traits.
MATERIALS AND METHODS
Plant Material
Twenty seven accessions of seeds originating from eleven locations covering the distribution area of G. kola in Cameroon were used. The number of accessions from each location was as follows: five accessions from Santchou (San), four from Fontem (Fon), four from Essoh-attah (Ess), three from Mbanga (Mba), three from Djoungo (Djo), three from Mbokambo (Mbo), one from Ebolowa (Ebo), one from Yokadouma (Yok), one from Ombessa (Omb), one from Sangmelima (Sang) and one from Mamfe (Mam). For each accession, fully mature orange fruits (14-week old) were harvested from a single tree in July 2007 and processed by removing the pericarp to extract the oval-shape seeds (Agyili et al., 2007) whose initial characteristics (i.e., weight, length and diameter) were determined before they were immediately used for germination and biochemical tests. Three hundreds mature seeds were used for each accession. The length and the diameter of each individual seed were measured using a calliper-square graduated to 0.1 mm and its mass was weighted by balance (Sartorius Basic), as described by Nehemie et al. (2007). After the above mentioned morphological parameters measurement, each accession was divided into three lots of 100, 150 and 50 seeds for viability test, germination essay and biochemical parameters measurement, respectively.
Viability Test
Seed viability was determined using 2, 3, 5-triphenyl-tetrazolium chloride (TZ). The TZ staining procedure is a standard test prescribed by the Association of Official Seed Analysts (AOSA) to determine the percentage of viable seeds in a lot; it was as follows: one hundred seeds from each accession were hydrated for 24 h at room temperature, cut longitudinally, placed in 0.1% TZ solution and incubated at 30°C for 5 h and examined for color change (Anonymous, 2000).
Germination Essay
Seeds from each accession were surface-sterilised using 1% (w/v) sodium hypochlorite for 10 min, followed by thorough rinsing with distilled water. They were then placed to germinate at room temperature (25±2°C) with a 12 h day/12 h night photoperiod in 20 cm sterile glass Petri dishes (3 cm deep) lined with two sheets of paper towels. Three replicates of 50 seeds were used for each accession. Seeds were watered when necessary and considered to be germinated once the radical or the shoot tip had emerged.
Percent germination was recorded at 2-weeks intervals for 30 weeks and the Germination Index (GI) was calculated as follows:
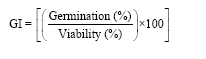 |
Rate of germination was estimated by using a modified Timson’s index of germination velocity:
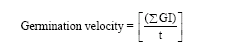 |
where, GI is the germination index at 2-weeks intervals and t is the total germination period (in weeks) (Khan and Ungar, 1997; Zia and Khan, 2004). The maximum value possible for germination velocity using this index was 50, i.e., [1500/30]. The higher the value, the more rapid the germination.
Carbohydrates and Amino Acids Analysis
Carbohydrates and amino acids were extracted according to Singh et al. (1990). Two grams of fresh seed were chopped and immediately ground to a fine powder and homogenized in 10 mL ethanol 70%. The mixture was placed under reflux for 20 min and the filter collected. The extraction was done twice and extracts from each sample were combined.
Carbohydrates contents were determined using the anthron method of Yemm and Willis (1954). One milliliter of extract was incubated with 5 mL anthron solution (0.12 g anthron in 100 mL 6.5 M H2SO4) at 90°C for 10 min. The absorbance of the green product was measured at 630 nm (Beckman DU-68 spectrophotometer). Glucose equivalents were calculated from a standard curve obtained with pure analytical grade glucose.
Amino acids contents were determined by the ninhydrin method of Yemm and Cocking (1955). The incubation mixture (4.1 mL final volume) containing 100 μL of the ethanolic extract, 1 mL of 70% ethanol, 1 mL of 0.2 M citrate buffer pH 5 and 2 mL of an acetonic ninhydrin solution (1% ninhydrin and 0.06% KCN in acetone), was incubated for 15 min at 100°C. The mixture was cooled for 5 min with tap water before 8 mL of distilled water was added. The absorbance was recorded at 570 nm. Glycine equivalents were calculated from a standard curve obtained with pure analytical grade glycine.
Proteins Analysis
Seed proteins were extracted according to the method described by Lanaud (1986). Seed tissues (2 g) were crushed in 5 mL 80% acetone in a mortar. Acetonic extract was removed by filtration using Whatman N° 3 paper. The residue was homogenized in 15 mL pre-chilled extraction buffer (0.1 M Tris-HCl pH 7.2 containing 10 mM cysteine-HCl and 10 mM Mg Cl2. 6H2O). The homogenate was centrifuged at 6000 g for 30 min at 4°C. The supernatant was collected and constituted our proteins extract. Proteins content was measured using the Coomassie Blue dye binding method of Bradford (1976), with pure analytical grade of bovine serum albumin as standard.
Phenols Analysis
Phenols were extracted at room temperature with 80% methanol as described by Bastide (1987). Phenols contents were measured spectrophotometrically at 725 nm using the Folin and Ciocalteu reagent (Marigo, 1973). The standard curve was established using pure analytical grade of chlorogenic acid.
Data Analysis
Data recorded for different variables were analysed using the statistical software package SPSS 12.0 for windows. Analysis of variances (ANOVA) was used to determine variability among means and the Student-Newman-Keuls test was used (p<0.05) to determine significant differences between means. The Spearman Rho coefficient of correlation was used to establish correlations between different variables surveyed. To establish relationship among individual accessions, a dendogram was generated by hierarchical cluster analysis based on quantitative biochemical, morphological and germination traits of seeds, using the statistical software package SPAD 4.1 for windows.
RESULTS
Biochemical Parameters of Seeds
Table 1 shows that phenols mean contents ranged from 35.78 μg g-1 FW for Fon1 to 1673.68 μg g-1 FW for San3. Analysis of variance showed significant variation among the accessions studied for this parameter.
| Table 1: | Phenols, proteins, carbohydrates and amino acids contents (μg g-1 FW) of different accessions of G. kola seeds |
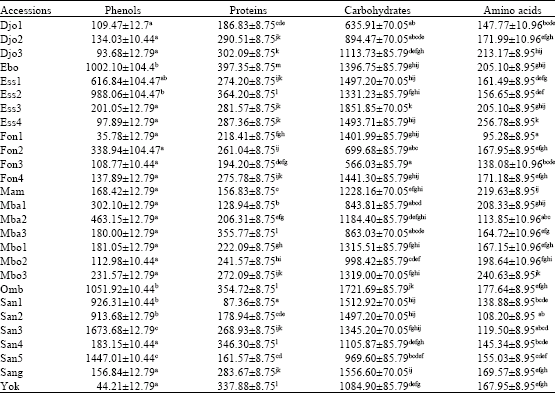 | |
| Each value represents Mean±SE (n = 6). Within the same column, means with same letter(s) attached are not significantly different at 5% level (Student-Newman-Keuls) | |
There were also significant differences among seed samples originating from same ecological zone, as Esso attah and Santchou for example. Phenols content was significantly higher in Ess2 than in Ess3 and Ess4, it was also higher in San3 and San5 than in San1, San2 and San4.
Proteins mean contents ranged from 87.36 μg g-1 FW for San1 to 397.35 μg g-1 FW for Ebo, with significant differences among the accessions studied. There were significant differences among seed accessions originating from the same location.
Carbohydrates mean contents of the seed accessions studied varied from 566.03 μg g-1 FW for Fon3 to 1851.85 μg g-1 FW for Ess3 and there were significant differences among the accessions studied as well as within accessions originating from the same location.
Amino acids mean contents ranged from 95.28 μg g-1 FW for Fon1 to 256.78 μg g-1 FW for Ess4, with significant differences among the accessions as well as within accessions originating from the same location (ecological zone).
Morphological and Germination Parameters of Seeds
Means ranged from 3.37 to 8.69 g for weight, from 2.39 to 3.42 cm for length, from 1.36 to 1.84 cm for diameter, from 0 to 89.7% for germination index and from 0 to 28.55 for germination velocity (Table 2). For each of these parameters, analysis of variance showed significant variation among the accessions studied and there were also differences within accessions originating from the same location.
Correlations Between Seeds Traits
Table 3 shows that there were significant (p<0.05) positive correlation between seed diameter and length (r = 0.413) and a highly significant (p<0.01) positive correlation between seed length and weight (r = 0.755) as well as between seed diameter and weight (r = 0.825).
| Table 2: | Quantitative morphological and germination traits of different accessions of G. kola seeds |
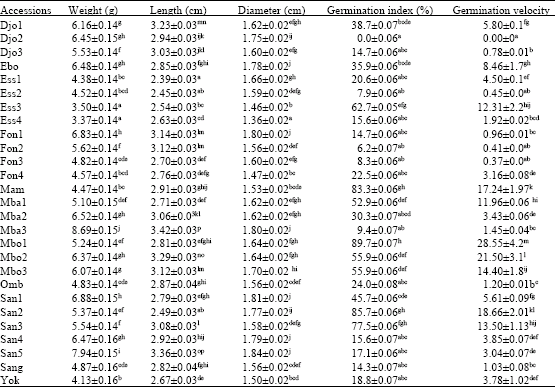 | |
| Each value represents Mean±SE of 300 measures for weight, length and diameter and Mean±SE of 3 replicates of 50 seeds each for germination index and germination velocity. Within the same column, means with same letter(s) attached are not significantly different at 5% level (Student-Newman-Keuls) | |
| Table 3: | Spearman’s Rho coefficients of correlation between different biochemical, morphological and germination parameters of G. kola seeds |
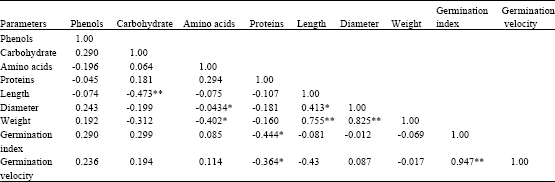 | |
| *Correlation is significant the 5% level (bilateral), **Correlation is significant at the 1% level (bilateral) | |
The germination parameters (germination index and germination velocity) were also significantly (p<0.01) correlated (r = 0.947). Amino acids contents were significantly (p<0.05) and negatively correlated to diameter (r = -0.434) and to weight (r = -0.402), while carbohydrates content w as significantly (p<0.01) and negatively correlated to length (r = -0.473). Proteins content of seeds was significantly (p<0.05) and negatively correlated to germination index (r = -0.444) as well as to germination velocity (r = -0.364).
Cluster Classification
The direct hierarchical classification of our accessions allowed obtaining 3 groups based on biochemical, morphological and germination parameters (Fig. 1). The group I, constituted of Ess3, Ess4, Ess1, Ess2, Omb, Fon4, San and Yok was characterised by high proteins and carbohydrates content, small size and small weight. Apart from Ess3 whose germination index and germination velocity were high, members of this group displayed poor seed germination.
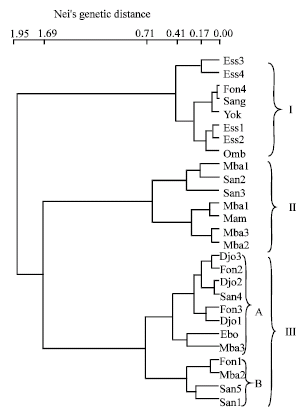 | |
| Fig. 1: | Direct hierarchical classification of G. kola seeds accessions based on biochemical, morphological and germination traits |
The group II which displayed high germination index and high germination velocity was also characterised by low proteins content. This group was constituted of Mbo1, San2, San3, Mba1, Mam, Mbo3 and Mbo2. Members of this group represented accessions displaying seven of the height top germination index. The group III was constituted of two sub-groups. The sub-group A constituted of Djo3, Fon2, Djo2, San4, Fon3, Djo1, Ebo and Mba3 was characterised by low carbohydrates content while sub-group B, characterised by low amino acids content and low proteins content associated with high weight was constituted of Fon1, Mba2, San5 and San1.
DISCUSSION
Information on intraspecific variation in G. kola is lacking. Results of the present study showed significant differences among G. kola seed accessions for their quantitative morphological, biochemical and germination traits. The above parameters also significantly varied within accessions originating from the same location, indicating that these variations may be attributed to genetic characteristics of trees rather than topographic or climatic characteristics of locations. Similar variations have been found in seeds of many others tropical species such as Dalbergia sissoo (Khurana and Singh, 2001), Irvingia gabonensis (Leakey et al., 2000; Atangana et al., 2002), Canarium schweinfurthii (Kapchie et al., 2002) and Dacryodes edulis (Leakey et al., 2003). The variation in germination traits may explain the diversity of results reported by former research on the improvement of G. kola seeds germination. These results contribute to knowledge on intraspecific variation in G. kola and could help in further selection and domestication processes of this species, which has suffered neglect in the areas of research and development, despite its socio-economic importance (Adebisi, 2004). Exploiting natural variation and using less dormant seed accessions as planting material could be promising tools for domestication and development of the cultivation of the species.
The highly significant negative correlation between carbohydrates content and diameter of seed is in agreement with the fact that people who use to eat bitter kola commonly agree that small size seeds are usually sweeter than large size seeds. Nevertheless, the physiological or biochemical explanation of this correlation, as well as the correlation between amino acid content and two quantitative morphological parameters of seeds (diameter and weight) remain to be elucidated. Results of the present study showed a significant negative correlation between proteins content of seeds and germination parameters (germination index and germination velocity). Moreover, dendogram generated by cluster classification put together in the same group accessions which performed best germination and members of this group were also characterized by low proteins content. These results indicated that seeds with low proteins contents germinated better than seed with high proteins contents. This result which contradicts that of Bhatt et al. (1989) on Irish potato seeds was rather surprising since it is widely known that a key factor in seed germination is the hydrolysis of storage proteins (Bewley and Black, 1978) which, during water uptake, may make them available as water soluble proteins or amino acids for embryo growth (Seal et al., 1972). Present result also contradict that of Lee et al. (2006), who reported that in Prunus campanulata, release from dormancy is accompanied by a significant increase in proteins content. At our knowledge, there is no former report indicating negative correlation between total soluble proteins content of seeds and germination capacity. Nevertheless, some proteins that play specific roles in seeds in the dormant state and in seeds during activation of germination have been identified: In Arabidopsis seeds, a repressor of germination have been identified and called RGL2 DELLA Protein, which is associated with dormant state (Lee et al., 2002). Disappearance of the RGL2 DELLA protein 5 h after dormancy-breaking treatment in Arabidopsis seeds (Tyler et al., 2004) suggested that a quick turnover of this repressor induces germination. Another protein has been identified in the same plant by Oh et al. (2005) as a negative regulator of seed germination. Moreover, a protein encoded by the pBS128 cDNA has been found associated to the maintenance of dormancy in the grass species Bromus secalinus (Goldmark et al., 1992) as well as in Hordeum vulgare (Reidunn et al., 1994). These former findings suggest that G. kola seed proteins may be dormancy related proteins.
CONCLUSION
Our ongoing research program which aims at identifying and characterising G. kola seeds proteins may shed light on the negative correlation between proteins content and germination traits reported in the present study.
REFERENCES
- Adebisi, A.A., 2004. A Case Study of Garcinia kola Nut Production-to-Consumption System in J4 area of Omo Forest Reserve, South-West Nigeria. In: Forest Products, Livelihoods and Conservation: Case Studies on Non-Timber Forest Systems, Sundenland, T. and O. Ndoye (Eds.). Vol. 2, CIFOR, Bogor, Indonesia, pp: 1-24.
- Atangana, A.R., V. Ukafor, P. Anegbeh, E. Asaah and Z. Tchoundjeu et al., 2002. Domestication of Irvingia gabonensis: 2. The selection of multiple traits for potential cultivars from Cameroon and Nigeria. Agrofor. Syst., 55: 221-229.
CrossRefDirect Link - Bhatt, A.K., T.C. Bhalla, H.O. Agrawal and M.D. Upadhya, 1989. Effect of seed size on protein and lipid contents, germination and imbibition in true potato seeds. Potato Res., 32: 477-481.
CrossRef - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Nehemie, D.T., F. Sanonne and O.N. Denis, 2007. Morpho-physical variation of fruits and impact on almond production of djansang (Ricinodendron heudelotii Baill.) in West and Centre of Cameroon. Pak. J. Biol. Sci., 10: 2838-2843.
CrossRefPubMedDirect Link - Iwu, M.M., O.A. Igboko and C.O. Okunji, 1987. Evaluation of the antihepatotoxic activity of the biflavonoids of Garcinia kola seed. J. Ethnopharmacol., 21: 127-142.
CrossRefDirect Link - Kapchie, N.V., C. Tchiegang, C.M. Mbofung and C. Kapseu, 2002. Variation of physical and chemical characteristics of fruits of Aiele (Canarium schweinfurthii Engl.) from different provinces in Cameroon. Proceedings of the 3rd International Workshop on Enhancing of Safou and Other Non-Conventional Oil Trees, (ES-NCOT'02), Yaounde, Cameroon, pp: 149-263.
- Khan, M.A. and I.A. Ungar, 1997. Effect of thermoperiod on recovery of seed germination of halophytes from saline conditions. Am. J. Bot., 84: 279-283.
Direct Link - Leakey, R.R.B., J.M. Fondoun, A. Atangana and Z. Tchoundjeu, 2000. Quantitative descriptors of variation in the fruits and seeds of Irvingia gabonensis. Agrofor. Syst., 50: 47-58.
CrossRefDirect Link - Leakey, R.R.B., K. Schrecheriberg and Z. Tchoundjeu, 2003. The participatory domestication of West African indigenous fruits. Int. For. Rev., 5: 338-347.
CrossRefDirect Link - Malik, A.R. and G.S. Shamet, 2009. Storage of Pinus gerardiana seeds: Biochemical changes and its applicability as vigour test. Res. J. Seed Sci., 2: 48-55.
CrossRefDirect Link - Emmanuel, N. and M. Roy, 2001. Effect of pre-sowing and incubation treatment on germination of Garcinia kola (Heckel) seeds. Fruits, 56: 437-442.
CrossRef - Yemm, E.W., E.C. Cocking and R.E. Ricketts, 1955. The determination of amino-acids with ninhydrin. Analyst, 80: 209-214.
CrossRefDirect Link - Yemm, E.W. and A.J. Willis, 1954. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J., 57: 508-514.
PubMedDirect Link - Emenky, F.A.O. and A.S. Khalaf, 2008. Effect of sowing depths and seed size of some winter cultivars of chickpea (Cicer arietinum L.) on field emergence and vegetative growth. Res. J. Seed Sci., 1: 11-18.
CrossRefDirect Link