
M.A. Baset Mia
Department of Land Management, Faculty of Agriculture, University Putra Malaysia, 43400 UPM, Serdang, Selangor DE, Malaysia
Z.H. Shamsuddin
Department of Land Management, Faculty of Agriculture, University Putra Malaysia, 43400 UPM, Serdang, Selangor DE, Malaysia
Research Journal of Seed Science
Year: 2009 | Volume: 2 | Issue: 4 | Page No.: 96-104







ABSTRACT
An experiment was conducted at the Soil Microbiology Laboratory, Department of Land Management, University Putra Malaysia, Malaysia to observe the effect of plant growth promoting bacterial inoculation on the enhancement of germination and vigor seedling production of rice. The design of the experiment was completely randomized with five replicates. Seeds were either inoculated by Bradyrhizobium strains UPMR48, UPMR29, Burkhloderia sp. strain SB13, Corynebacterium sp. strains SB35 and SB41 or un-inoculated control or cultural media only. The results pointed out that inoculation significantly influenced the seedling emergence and vigor and seeds inoculated with strain UPMR29 and UPMR48 showed better performance in respect of vigor index. Inoculation of seeds with UPMR29 showed the longest plumule (shoot length). Bacterial inoculation also influenced on the root hair formation where UPMR 48 showed earlier root hair initiation than others. The overall root growth and development were positively influenced by the bacterial inoculation. The results concluded that plant growth promoting bacteria UPMR29 and UPMR48 could be used for vigor seedling production of rice.
PDF Abstract XML References Citation
How to cite this article
M.A. Baset Mia and Z.H. Shamsuddin, 2009. Enhanced Emergence and Vigor Seedling Production of Rice through Growth Promoting
Bacterial Inoculation. Research Journal of Seed Science, 2: 96-104.
URL: https://scialert.net/abstract/?doi=rjss.2009.96.104
URL: https://scialert.net/abstract/?doi=rjss.2009.96.104
INTRODUCTION
Rice (Oryza sativa L.) is the major food crop of nearly half of the world’s population. For over half a century, the world has relied on increasing crop yields to supply an ever increasing demand for food. World rice production increased significantly during last two decades. This dramatic increase in world grain production was the result of a 122% increase in crop yields. However, this trend of grain production cannot be maintained due to decreasing the cultivable land for rapid urbanization. Therefore, vertical expansion for food production is essential. In order to increase world food production in a sustainable manner, farmers have to use quality seeds. Good quality and viable seed is required for rapid and synchronous seedling which is prerequisite for successful crop establishment and uniform crop growth and development and finally the yield. Vigor test provides additional information on the rate and uniformity of seedling growth under wide range of environmental conditions (Powell and Matthews, 2005). Seedling vigor is critical when competition for light, air and water becomes strong. Seedlings with vigorous growth pattern can compete successfully under stress, influencing stand establishment and ultimately grain yield. Germination is a critical stage in the development and life cycle of many plants; it ensures the production and consequently controls the dynamics of the population. Seed germination can be divided into three phases: imbibition, increased metabolic activity and initiation of growth, which loosely parallel the triphasic water uptake of mature dry seeds (Finch-Savage and Leubner Metzger, 2006; Ghars et al., 2009).
Plant Growth Promoting Bacteria (PGPB) are bacteria that improve plant growth when introduced onto seeds, seed pieces, roots, or into soil. The PGPB improve plant growth by one or more mechanisms: direct stimulation of plant growth by the production of phytohormones, vitamins and siderphores; enhancement of nutrient uptake; suppression of plant pathogens and/or induction of resistance in plant hosts against pathogens (Mayak et al., 2004; Alikhani et al., 2006; Dakora, 2003; Dobbelaere et al., 2003). The inoculation of seeds or seedlings with microbial inocula has been adopted as a method of modifying microbial populations around crop plants to promote both development and yield. The stimulation of seedling development by bacteria has also been attributed to the production of biologically active compounds. A significant increase in grain yield was also recorded in rice plants inoculated with plant growth promoting bacteria (Singh et al., 2006; Francine et al., 2007). Similarly, inoculation of rhizobia to rice produced significantly higher root and shoot biomass; increased their photosynthetic rate and accumulated higher levels of indoleacetic acid and gibberellin growth-regulating phytohormones (Feng et al., 2005).
It has been shown that Rhizobium sp. can develop on the surface of monocots similarly to dicotyledons and that Bradyrhizobium sp. can grow efficiently on grass or legumes seeds during germination, stimulating root development in a similar manner to free-living bacteria (Pena-Cabrales and Alexander, 1983). The present investigation was designed to assess the range of growth promoting activities of various plant growth promoting bacteria on seedling emergence, vigor, growth and development of rice.
MATERIALS AND METHODS
The experiment was conducted at the Soil Microbiology Laboratory, Department of Land Management, Faculty of Agriculture, University Putra Malaysia during July to September 2009. The design of the experiment was completely randomized with five replicates. The rice cultivar MR219 (Oryza sativa L.), collected from the Malaysian Agricultural Research and Development Institute (MARDI), Malaysia were used in the experiment. Following PGPB strains were used as test materials:
 |
Strains were grown in yeast manitol broth (YMB) (Somasegaran and Hoben, 1994) and nutrient broth. Exponentially growing cells in shaken broth culture were inoculated. Rice seeds was surface sterilized by 95% ethanol in an Erlenmeyer flask and was treated with 1% sodium hypochlorite (Clorox) for 2 min followed by six times washing with sterile water. Thereafter the seeds were soaked in various Bradyrhizobial or PGPB grown broths. Seeds soaking in yeast manitol broth and nutrient broth were treated as control. Twenty five seeds either inoculated or control were put in sterilized Petri dish containing filter paper (Whatman No. 102) and the Petri dish were kept an incubator at 30°C for 120 h.
Following parameters were made for seed germination:
Seedling Emergence Test
After soaking, the air-dried seeds were used for germination and the seedling percent emergence was calculated with the following formula:
Co-efficient of germination and germination index were calculated using following formula (Copeland, 1976):
Where:
| A | = | No. of seed germinated |
| T | = | Time corresponding to A |
| x | = | Counting number (1, 2, 3 ...nth) Ti Ni |
Where:
| Ti | = | The ith No. of days after sowing |
| Ni | = | The ith No. of seeds emergence |
| S | = | The total No. of seed used |
Plumule and radical length were measured by a meter scale and root surface area, root volume and total root length were measured by a root image scanner (Model: WinRhizon-Analysis for Root, Regent Instruments Inc.). The roots and shoots were separated and keep in an electric oven at 48°C for 72 h for taking a constant dry weight. Seedling vigor index was measured according to the modification of Abdul Baki and Anderson (1973).
Statistical Analysis
Treatments were arranged in a completely randomized design, the mean, standard error analysis of variance and the LSD were calculated by using SAS package, Version 9.0 (SAS Institute Inc., 2004). Means were compared using the Least Significance Difference (LSD) or Duncan Multiple Range Test (DMRT) at 5% level.
RESULTS
Seedling Emergence Percentage
Seedling emergence was positively influenced by the application of different plant growth promoting bacterial inoculation (Fig. 1). Although, initially there were no pronounced
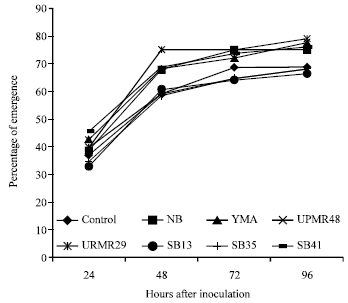 | |
| Fig. 1: | Effects of different plant growth promoting bacterial inoculation on the percent seedling emergence of rice seeds variety MR219 |
differences which accelerated with time and a clear differences were visible at 48 h after inoculation where strain UPMR29 showed higher percentage of germination. Finally, both control and inoculated seeds showed similar percent of emergence.
Emergence Index
The seedling emergence index increased with time (Fig. 2a, b). There were no significant differences between inoculated and un-inoculated seeds. The trend of emerging index was linear.
Vigor Index
Inoculation significantly influenced on the seedling vigor index and seeds inoculated with Bradyrhizobium spp. strain UPMR48 and UPMR29 showed the higher seedling vigor as compared to control (Table 1). However, seeds inoculated with SB35 showed the poorest vigor index. Strains SB 41 and SB13 showed lower but comparable vigor index. Application of nutrient broth also showed the higher vigor index.
Seedling Growth
Inoculation of PGPB significantly influenced the plumule (shoot) length (Table 1). The seeds inoculated with UPMR29 showed the longest shoot and application of SB35 showed the shortest shoot than others. Application of nutrient broth also showed the higher shoot length.
Root Growth
Application of PGPB on the seeds of rice significantly influenced on the root hair formation where seeds inoculated with SB35, SB41 and UPMR 48 initiated root hair at 48 h after inoculation whereas control and other strains initiated root hair after 72 h of inoculation (Table 2). Similarly, those two strains showed earlier initiation of lateral roots. Inoculation of SB13 and SB35 showed deformed or black tip of roots. All the strains showed root hair until
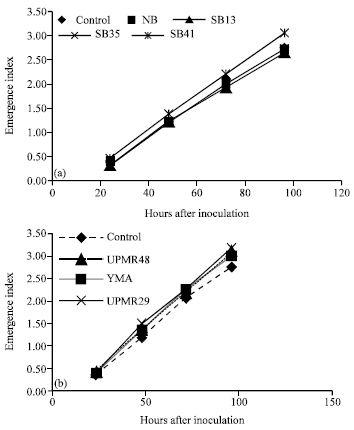 | |
| Fig. 2: | (a, b) Effects of different plant growth promoting bacterial inoculation on the seedling emergence index of rice seeds variety MR219 |
| Table 1: | Effects of different PGPB strains on the germination, vigor index, shoot length and total dry matter of rice seedlings variety MR 219 |
 | |
| In a column means having same letter (s) do not differ significantly at 5% level of significant | |
| Table 2: | Initiation of root hair in rice variety MR219 as influenced by plant growth promoting bacterial inoculation |
 | |
| +: Presence, -: Absence | |
96 h of inoculation except control and roots inoculated with SB13 and SB41. Inoculation of plant growth promoting bacteria significantly influenced the root length of rice (Table 3).
| Table 3: | Effects of different PGPB inoculation on the root growth and development of rice variety MR219 |
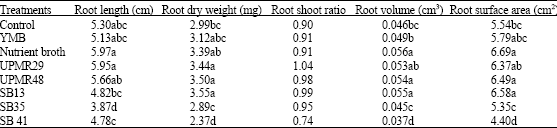 | |
| In a column means having same letter (s) do not differ significantly at 5% level of significant | |
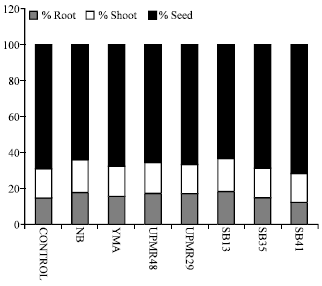 | |
| Fig. 3: | Effect of PGPB inoculation on the seed reserve mobilization of rice variety MR219 |
Seeds treated with UPMR 29 produced significantly the longest radical compared to others. Application of nutrient broth also increased the root length. The shortest root was also produced by the inoculation of SB35. The remaining treatments showed the intermediate status. Similarly, root dry weight also significantly influenced by the application of PGPB (Table 3). Inoculation with strain UPMR48, UPMR29 and SB13 significantly increased the root biomass as compared to control treatments. Inoculation with SB35, SB41 showed the lowest performance compared to others. Root to shoot ratio also influenced by the application of UPMR 29 indicating higher root growth than shoot.
Root Volume and Surface Area
Inoculation of PGPB significantly influenced on the production of root surface area and root volume (Table 3). The strain UPMR48 and SB13 produced the greatest root volume and similarly the root surface area. However, the strain SB41 recorded the lowest root volume and surface area.
Mobilization of Seed Reserve
The PGPB inoculation significantly influenced on the seed reserve mobilization of rice (Fig. 3). Inoculated seeds with SB13 and UPMR48 remained less amount of food in the endosperm. Seeds inoculated with SB13 and nutrient broth showed higher seedling shoot percentage than others. Similarly, SB13, UPMR29 and UPMR48 showed higher percentage of root than others.
DISCUSSION
Plant growth promotion along with N2 fixation by bacteria might be performed important factors for achieving a sustainable agriculture in the future by saving the environment. Since, various PGPB have been found diazotrophic bacteria have been found in association with graminaceous plants, which are possible candidates for beneficial interactions with agriculturally important crops. The experimental results clearly indicated that significant improvement was made on seedling emergence and seedling vigor due to PGPB inoculation. Vigor seeds are precondition for better seedling establishment consequently the higher yield (McDonald and Copeland, 1997). Induction of root hair, lateral roots formation, longer roots and consequently the root mass was the results due to PGPB inoculation. This root enhancing effect might be due to the production of phytohormone especially IAA where Noel et al. (1996) reported significance enhancement of early seedling root growth in non-legumes by seed inoculation with R. leguminosarum and attributed this effect to bacterial phytohormone production. It is well-known that inoculation of cereals with Plant Growth Promoting Bacteria (PGPB) resulted in increased crop growth and yield and act as biofertilizer and bioenhancer for different non-legumes (Andrews et al., 2003; Mia et al., 2009; Shamsuddin et al., 1999). However, there is general agreement that these growth responses were not due to N2 fixation by the bacterium, but were primarily related to the bacterial production of phytohormone, which caused changes in root morphology and physiology that resulted in increased nutrient and water uptake from the soil (Andrews et al., 2003; James, 2000; Mantellin and Touraine, 2004; Mia et al., 2009). Similar crop bio-enhancing activity in different non-legumes has been observed due to rhizobial inoculation. These growth enhancing phenomena are more pronounced in roots rather shoot.
In terrestrial plants, roots serve several functions namely conducts to supply both nutrients and water to the plants from the soil, a location for the synthesis and exchange of various plant hormones, storage organs of plant resources and the anchorage of the plant (Fitter, 1991; Schiefelbein and Benfey, 1991). Root development is clearly influenced by the activity of phytohormone especially auxins, which are known to influence a number of plant functions such as promotion of cell elongation and cell division apical dominance, root initiation, differentiation of vascular tissue, ethylene biosynthesis mediation of tropic responses and alteration of the expression of specific gene (Chasan, 1993; Key, 1989; Sachs, 1993; Warren-Wilson and Warren-Wilson, 1993). There are evidences that certain strains of Rhizobium can enhance rice growth and yield through the accompanying changes in growth physiology or root morphology rather than biological N2 fixation (Biswas et al., 2000). Rhizobium sp. are capable of synthesizing IAA in absence of tryptophan, but the exogenous application of tryptophan increases IAA production several fold (Kittell et al., 1989). Increased rice root systems with a significantly greater absorptive surface area and extra cellular bioactive metabolites that can promote rice root development resulting in expansive root architecture were also observed by the application of Rhizobium.
Inoculation of rhizobia can increase N uptake by rice plant which was reported by Yanni et al. (1997). These raises questions of whether this benefit of rhizobia to rice may be due to their associative N2 fixing activity and/or their ability to change the phytohormone balance, thereby influencing growth physiology in ways that affect major nutrient uptake in rice.
The greater total Dry Matter (DM) accumulation by rice seedling due to inoculation with Bradyrhizobia strain UPMR29 and UPMR48 on rice roots, their longevity is nevertheless adequate to trigger plant growth stimulation and vigor of young seedlings that carryover to produce more productive plants, resulting in higher yields at maturity (Biswas, 2000). Yanni et al. (2001) has found good response of phytohormone production in rice by inoculating Rhizobium leguminosarum bv. Trifolli. In present results, inoculation significantly increased the vigor index which determines the state of the healthy seedling and ultimately the condition of the productivity of the plant. Similarly higher the vigor index better will be the yield of crop plants.
CONCLUSIONS
The results have provided useful information about the responses of PGPB on rice seed emergence and seedling development. It is indicated that inoculation of Bradyrhizobium sp. strain UPMR48 and UPMR29 resulted in an increased seed emergence, seedling vigor and seedling root growth and development. Considerable improvements were obtained in root morphology namely root length, root surface area, volume and consequently the mass. Seeds inoculated with UPMR48 showed better performance in respect of seedling growth although, percent germination was not influenced. In conclusion, the results suggested that simultaneous screening of PGPB for seedling establishment under pot and field experiment is a good tool to select effective PGPB for biofertilizer development biotechnology.
REFERENCES
- Alikhani, H.A., N. Salesh-Rastin and H. Antoun, 2006. Phosphate solubilization activity of rhizobia native to Iranian soil. Plant Soil, 287: 35-41.
Direct Link - Abdul-Baki, A.A. and J.D. Anderson, 1973. Vigor determination in soybean seed by multiple criteria. Crop Sci., 13: 630-633.
CrossRefDirect Link - Biswas, J.C., J.K. Ladha, F.B. Dazzo, Y.G. Yanni and B.G. Rolfe, 2000. Rhizobial inoculation influences seedling vigor and yield of rice. Agron. J., 92: 880-886.
CrossRef - Dobbelaere, S., J. Vanderleyden and Y. Okon, 2003. Plant growth promoting effects of diazotrophs in the rhizosphere. Crit. Rev. Plant Sci., 22: 107-149.
Direct Link - Chi, F., S.H. Shen, H.P. Cheng, J.X. Jing, Y.G. Yanni and F.B. Dazzo, 2005. Ascending migration of endophytic rhizobia, from roots to leaves, inside rice plants and assessment of benefits to rice growth physiology. Applied Environ. Microbiol., 71: 7271-7278.
Direct Link - Francine, M., W. Perrine, E. Gartner, H.H. Charles, A. Becker and G.R. Barry, 2007. Rhizobium-initiated rice growth inhibition caused by nitric oxide accumulation. Mol. Plant Microbe Interac., 20: 283-292.
Direct Link - Finch-Savage, W.E. and G. Leubner-Metzger, 2006. Seed dormancy and the control of germination. New Phytol., 171: 501-523.
CrossRefPubMedDirect Link - Ghars, M.A., A. Debez and C. Abdelly, 2009. Interaction between salinity and original habitat during germination of the annual seashore halophyte Cakile maritime. Commun. Soil Sci. Plant Anal., 40: 3170-3180.
Direct Link - James, E.K., 2000. Nitrogen fixation in endophytic and associative symbiosis. Field Crops Res., 65: 197-209.
CrossRefDirect Link - Kittell, B.L., D.R. Helinski and G.S. Ditta, 1989. Aromatic aminotransferase activity and indoleacetic acid production in Rhizobium meliloti. J. Bacteriol., 171: 5458-5466.
Direct Link - Mantelin, S. and B. Touraine, 2004. Plant growth-promoting bacteria and nitrate availability: Impacts on root development and nitrate uptake. J. Exp. Bot., 55: 27-34.
CrossRefDirect Link - Mia, M.A.B., Z.H. Shamsuddin, W. Zakaria and M. Marziah, 2009. The effect of rhizobacterial inoculation on growth and nutrient accumulation of tissue-cultured banana plantlets under low N-fertilizer regime. Afr. J. Biotechnol., 8: 5855-5866.
Direct Link - Mayak, S., T. Tirosh and B.R. Glick, 2004. Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol. Biochem., 42: 565-572.
CrossRefDirect Link - Noel, T., C. Sheng, C. Yost, R. Pharis and M. Hynes, 1996. Rhizobium leguminosarum as a plant growth promoting rhizobacterium: Direct growth promotion of canola and lettuce. Can. J. Microbiol., 42: 279-283.
Direct Link - Pena-Cabrales, J. and M. Alexander, 1983. Growth of Rhizobium in unammended soil. Soil Sci. Am. J., 47: 81-84.
Direct Link - Sachs, T., 1993. The role of auxin in the polar organization of apical meristems. Aust. J. Plant Physiol., 20: 531-533.
CrossRefDirect Link - Schiefelbein, J.W. and P.N. Benfey, 1991. The development of plant roots: New approach to underground problems. Plant Cell., 3: 1147-1154.
Direct Link - Shamsuddin, Z.H., A.H. Ghazali and M.A.B. Mia, 1999. Azospirillum as a bioenhancer and biofertilizer for banana and oil palm seedlings. Biotechnol. Sustain. Util. Biol. Resour. Trop., 13: 326-338.
Direct Link - Singh, R.K., P.N.M. Ravi, K.J. Hemant, K.S.P.P. Vinod, B.R. Sasi and A. Kannepalli, 2006. Isolation and identification of natural endophytic rhizobia from rice (Oryza sativa L.) through rDNA PCR-RFLP and sequence analysis. Curr. Microbiol., 52: 345-349.
CrossRef - Warren-Wilson, J. and P.M. Warren-Wilson, 1993. Mechanisms of auxin regulation of structural and physiologic polarity in plants, tissues, cells and embryos. Aust. J. Plant Physiol., 20: 555-571.
Direct Link - Yanni, Y.G., R.Y. Rizk, F.K.A. El-Fattah, A. Squartini and V. Corich et al., 2001. The beneficial plant-growth promoting association of Rhizobium leguminosarum bv. Trifolii with rice roots. Aust. J. Plant Physiol., 28: 845-870.
Direct Link - Dakora, F.D., 2003. Defining new roles for plant and rhizobia molecules in sole and mixed plant cultures involving symbiotic legumes. New Phytol., 158: 39-49.
Direct Link - Yanni, Y.G., R.Y. Rizk, V. Corich, A. Squartini and K. Ninke et al., 1997. Natural endophytic association between Rhizobium leguminosarum bv. trifolii and rice roots and assessment of its potential to promote rice growth. Plant Soil, 194: 99-114.
CrossRefDirect Link