
M. Otroshy
Agricultural Biotechnology Research Institute of Iran (ABRII), P.O. Box 85135-487, Isfahan, Najafabad, Iran
P.C. Struik
Crop and Weed Ecology, Department of Plant Sciences, Wageningen University, Haarweg 333, 6709 RZ Wageningen, Netherlands
Research Journal of Seed Science
Year: 2008 | Volume: 1 | Issue: 1 | Page No.: 41-50







ABSTRACT
A greenhouse experiment was carried out to investigate the effects of different sizes of normal seed tubers (35-45, 45-55 and 55-65 mm) and various growth regulators (gibberellic acid: 2.5 and 5 mg L-1; chlormequat: 10 and 100 mg L-1; included two controls: cut seed tubers and uncut seed tubers) on the dormancy, sprout behaviour, growth vigour and quality of tubers of different potato cultivars Frieslander, Marfona and Sante. Size of tuber, growth regulator and cultivar showed a highly significant effect on the sprouting behaviour. Highest number of sprouts with longest length and highest weight were produced by the size 55-65 mm. Highest dose of GA resulted highest number of sprouts and uncut control produced lowest number of sprouts. Frieslander produced higher number of sprouts and also more weight of sprouts than other cultivars. All parameters of vigour were highly significantly affected by cultivars and growth regulators while seed tuber size only affected number of stems. There were many significant two-way interactions between seed tuber size, cultivar or growth regulator application.
PDF Abstract XML References Citation
How to cite this article
M. Otroshy and P.C. Struik, 2008. Effects of Size of Normal Seed Tubers and Growth Regulator Application on Dormancy, Sprout Behaviour, Growth Vigour and Quality of Normal Seed Tubers of Different Potato Cultivars. Research Journal of Seed Science, 1: 41-50.
URL: https://scialert.net/abstract/?doi=rjss.2008.41.50
URL: https://scialert.net/abstract/?doi=rjss.2008.41.50
INTRODUCTION
Generally, the propagation material used to grow the potato crop is the seed tuber. A tuber is a greatly shortened and thickened stem that bears scale leaves with axillary buds (Van Ittersum, 1992a). The vegetative propagation is the cheapest and easiest way of potato propagation yet, but it has some disadvantages. One of these disadvantages is the low multiplication rate that can be achieved in this way.
The physiological age of the seed tuber is crucial for its quality as planting material. The physiological age influences the productive capacity of the seed tuber (Reust, 1986), but also changes the quality of the produce (e.g., by influencing the tuber size distribution; Struik and Wiersema, 1999).
In temperate regions, potatoes can only be grown once a year, in contrast to tropical and sub-tropical regions where potatoes can be grown year round. In the temperate regions, there are almost no problems with the physiological age of seed tubers, because of the natural dormancy period during the winter when the harvested seed potatoes are stored in good cold storage facilities. However, in tropical and sub-tropical regions large problems with physiological age of seed tubers can occur. This problem is caused by the short period between two potato crop cultivations and by the high storage temperatures when no cold stores are available. The seed tubers can be physiologically too young (when taken from the previous crop: short storage) or too old (when taken from last year`s crop: long storage at high temperatures) for planting (Van Ittersum, 1992a; Struik and Wiersema, 1999).
Crops from physiologically young seed tubers grow longer and are more yielding than those from physiologically old seed tubers. Wiersema (1985) found that crops from physiologically young seed tubers produce fewer stems, have later tuberization, more foliage and more tubers per stem than crops from physiologically old seed tubers.
Not only physiological age, but also other factors affect the vigour of the potato plant. One of these factors is the seed tuber size. According to Wiersema (1989), seed tubers below 20 g are less yielding per stem and have less foliage production per stem than seed tubers heavier than 20 g. Seed tubers heavier than 100 g showed more sprouts with longer length compared to small potato seed tubers of 2.5 g and gave a steady ground cover and yield. Van Ittersum (1992a) found a relation between the size of the seed tuber and its physiological age. Small seed tubers had a longer dormancy, so they were physiologically younger than bigger seed tubers. However this effect depended on cultivar, within some cultivars hardly an effect of size over a wide size range, but in other cultivars a significant response over a wide range of sizes. Lommen and Struik (1993), Lommen (1994) and Ranalli (1997) showed that size effects on dormancy and vigour are even more important for minitubers.
Physiological ageing can be controlled by storage conditions. With higher temperatures during storage, ageing goes faster than with lower temperatures (Struik and Wiersema, 1999). However, the way the temperature sum is built up during the storage season is also important, especially for cultivars with a high rate of natural ageing (Struik et al., 2006).
The trend of vigour after the dormancy period varies among cultivars (e.g., Van der Zaag and Van Loon, 1987; Van Ittersum, 1992b; Struik and Wiersema, 1999; Struik et al., 2006; Bodlaender et al., 1985). The optimum growth vigour of a cultivar at a certain time can be reached by programming storage conditions properly. These optimal storage conditions are also different among cultivars (Struik and Wiersema, 1999).
Cutting of the seed tuber can influence the early growth vigour in a positive way, because cutting may break dormancy. Also apical dominance will be broken, the sprouts will grow faster and more stems will grow out from one seed tuber (Struik and Wiersema, 1999). De-sprouting of relatively young seed tubers may contribute to the development of new sprouts and more sprouts with a higher rate of sprout growth than without de-sprouting (Struik and Wiersema, 1999).
It was discovered that gibberellins play a very important role in the regulation of growth and development in higher plants (Mohr and Schopfer, 1995). In the potato plant, GA plays an important role in different developmental events of the plant, but especially in tuber formation (Struik et al., 1999).
Shoot growth is promoted by an exogenous GA3 treatment and plant height is increased but tuber weight is decreased (Menzel, 1980). A foliar spray of GA also results in long stolons (Hammes and Nel, 1975).
Suttle (2004), on the other hand, observed that continued exposure of developing tubers to inhibitors of GA biosynthesis did not extend tuber dormancy but rather hastened dormancy release. CCC promoted growth of root systems in tomato and bean plants (Tognoni et al., 1966). The basic, direct physiological effect of CCC is that of blocking the biosynthesis of GA in a very early stage (Rademacher, 1999).
Indeed, sprouting is inhibited by CCC (Majeed and Bano, 2004). Shoot growth is reduced by a foliar application of CCC: CCC reduces stem length, leaf area and internode length (Dyson, 1965; Digby and Dyson, 1973). A foliar application of CCC reduced tuber yield (Rex, 1992).
Information on the effects of CCC on sprouting and vigour of seed tubers when applied directly to the seed tubers is scarce.
In this study, the effects of two different doses of GA and CCC applied directly on the seed tuber, on the sprouting behaviour and growth vigour of seed tubers will be described, using different sizes of normal seed tubers of normal to relatively high physiological age and different cultivars. Moreover, the effect of cutting of the seed tubers (necessary to effectively apply the growth regulator) will be assessed.
MATERIALS AND METHODS
This study was conducted in 2005 at the Unifarm of the Crop Science Department of Wageningen University and Research Center.The experiment included three factors: size of normal seed tubers (35- 45, 45-55 and 55-65 mm), growth regulator (GA and CCC, including two controls) and potato cultivars (Frieslander (early), Marfona (mid-early) and Sante (late). The factor growth regulator consisted of six levels: two doses of GA3 (2.5 and 5 mg L-1), two doses of CCC (10 and 100 mg L-1) and two controls (seed tubers cut and seed tubers uncut). The experiment consisted of two parts. The first part was a sprout behaviour test and the second part was a vigour experiment. The seed tubers were delivered by Unifarm of Wageningen University. First, seed tubers were accurately sorted out by a grader into the three sizes (35-45, 45-55 and 55-65 mm). The seed tubers were stored in a cold store at 4°C and a relative humidity of 80%. For the sprout behaviour test, 10 seed tubers per treatment combination were selected. All seed tubers (except those for the control treatment with uncut seed tubers) were hand cut and an incision was made to two thirds of the longest axis of the seed tubers, to allow growth regulators to penetrate. To prevent the incision from closing a piece of paper was inserted in the cut. After all incisions were made, growth regulators were applied by soaking the seed tubers in the appropriate solution for 10 min. Treatments included 2.5 or 5 mg L-1 gibberellic acid, 10 or 100 mg L-1 CCC, or demi-water. Seed tubers were then stored for a period of 11 days at 18°C, full darkness and a very high humidity to allow wound healing and sprouting. After 11 days, the seed tubers had produced sprouts and were transferred to the cold store (4°C; 80% relative humidity) to allow recording of data on sprouting at the same stage of development. Data were collected on number of sprouts, length of longest sprout and weight of sprouts to assess the effects of seed tuber size and growth regulators on sprouting capacity. As also the weight of the sprouts was recorded, the seed tubers were de-sprouted and the second part of the experiment was therefore carried out with de-sprouted seed tubers. De-sprouting enhances the expression of the effects of storage temperatures on growth vigour (Struik and Wiersema, 1999) and may also enhance the effects of the growth regulators.
When measurements on sprout behaviour were completed, the second part of experiment was started. This part was designed to assess the effects of the treatments on the growth vigour of the seed potato tubers. Seed tubers were planted into 5 L pots (filled with potting soil without fertilizer) at a depth of 8 cm. Pots were randomly placed in a greenhouse under the following conditions: day temperature 18°C (12 h), night temperature 12°C (12 h), relative humidity 80%. Daylight (12 h) was supplemented with 0.8 Philips SON-T-Agro lamps 400 W m-2 and with incandescent illumination.
Observations
For part one of the experiment (the sprout behaviour test) records were taken on number of sprouts, length of longest sprout and weight of sprouts. This set of parameters provides a good characterization of the sprouting capacity of the seed tubers (Struik and Wiersema, 1999). The criterion for a sprout was a structure of at least 2 mm length (Van Ittersum, 1992a). Length of the longest sprout was measured by taking the distance from the tip of the longest sprout to the base of sprout.
For part two (the vigour test) records taken included date of emergence [(Days After Planting (DAP)], number of stems, length of the longest stem, date of leaf death (or date of senescence; days after planting), duration of growth (date of senescence minus date of emergence; days), number of progeny tubers, dry weight of tubers and dry weight of aerial parts of plant. This set of parameters provides a characterization of both the above ground shoot development and the rate of multiplication and yield of tubers. The date of emergence was assessed as the day when the top of the first stem broke the soil surface. Not all seed tubers produced an emerged plant. When no plant was produced, the experimental unit was either recorded as one with a missing value or the value of 126 DAP (the end of the experiment) was assigned. The latter procedure not only allowed for a more reliable statistical analysis, it also gave a better reflection of the actual effect of the treatments.
The number of stems was counted at 72 DAP. The length of the longest stem was measured at 73 DAP. The date of senescence (date of leaf death) was defined as the day when all leaves were yellow. Because some seed tubers did not produce an emerging plant, the date of senescence for these cases was either recorded as a missing value or set at 126 days (the end of the experiment). The days of growth were calculated as the difference between day of senescence and day of emergence. For seed tubers that did not produce an emerging plant, the duration of growth was recorded as a missing value or set at 0 (126 DAP minus 126 DAP).
To assess above ground dry matter, the aerial part of the plants were cut and dried in a forced ventilated oven at 105°C for at least 24 h. Progeny tubers were harvested and number and weight of progeny tubers were recorded. Tubers were chopped and dried in an oven at 105°C for at least 24 h to assess the dry weight of the progeny tubers.
Statistical Analysis
The entire experiment was executed in 10 replications and included 540 experimental units. The experimental design was a completely randomized factorial experiment with three factors. Analysis of variance was applied on the variables using the MSTATC statistical package. Duncan`s Multiple Range Test was used to compare the means.
RESULTS
Sprouting Behaviour
Tuber size, cultivar and growth regulator all had a highly significant effect on the sprouting behaviour, as illustrated by the length, number and weight of sprouts (Table 1). There was a significant effect of seed tuber size on the length of the longest sprout. The longest sprouts were produced by the size 55-65 mm.
| Table 1: | The effect of tuber size and growth regulators on the sprout behaviour after growth regulators application in different cultivars of potato |
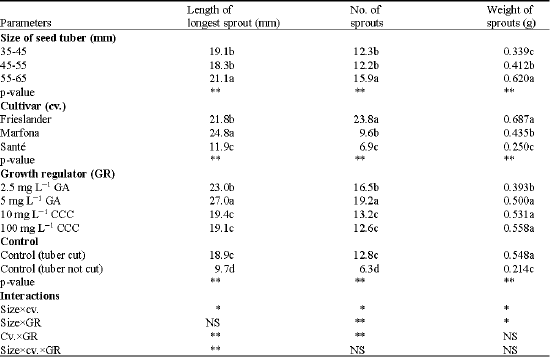 | |
| **Significant at p≤0.01, *Significant at p≤0.05. Values followed by a similar letter(s) are not statistically significantly different. GA: Gibberellic Acid, CCC: Chlorocholine Chloride, cv.: Cultivar, GR: Growth Regulator. NS: Not Significant, p≤0.05 | |
The effect of cultivar on the length of the longest sprout was highly significant. The effect of growth regulator on the length of the longest sprout was also highly significant (p < 0.0001). The seed tubers treated with GA produced the longest sprouts.
The main effects of the three factors were all significant for the parameter number of sprouts. For the factor seed tuber size, the biggest size (55-65 mm) had significantly more sprouts than the other sizes, which were the same. For the factor growth regulator (p < 0.0001), the highest dose of GA (5 mg L-1) produced the highest number of sprouts. The uncut control produced the lowest number of sprouts (Table 1).
All main factors showed a highly significant effect on the weight of sprouts (p < 0.0001). The bigger the seed tuber size, the higher the sprouts weight. Frieslander produced more sprout weight, especially than Sante. CCC (100 mg L-1) produced the highest sprout weight, but showed no significant difference with GA 5 mg L-1, both doses of CCC 10 mg L-1 and the cut control.
Vigour
Of all vigour parameters measured, size of seed tubers only affected the number of stems while highly significant effects of cultivar and growth regulators on all vigour parameters assessed were shown (Table 2, 3).
The main effect of cultivar on the date of emergence was highly significant (p < 0.0001). Sante had the latest emergence. The main effect of growth regulator was also highly significant (p < 0.0001). The uncut control gave the latest emergence.
All main effects on the number of stems were significant (Table 3). The seed tuber size effect (p = 0.0017) showed that the biggest seed tuber size produced significantly more stems than the lower two seed tuber sizes. Frieslander produced most stems. Sante produced the lowest number of stems. Seed tubers treated with GA 2.5 mg L-1 produced the highest number of stems. The uncut control produced the lowest number.
| Table 2: | The effect of seed tuber size and growth regulators on the vigour in different cultivars of potato |
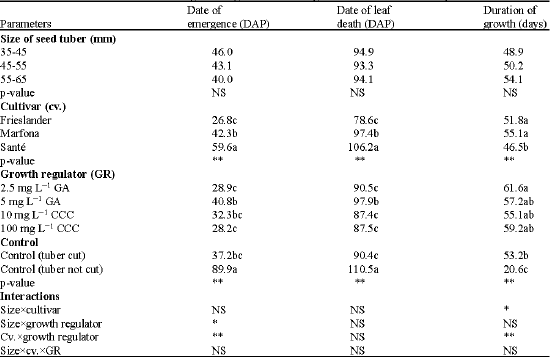 | |
| **Significant at p≤0.01, *Significant at p≤0.05. Values followed by a similar letter(s) are not statistically significantly different. DAP: Day After Planting, DAE: Dat After Emergence, GA: Gibberellic Acid, CCC: Chlorocholine Chloride, cv.: Cultivar, GR: Growth Regulator. NS: Not Significant, p≤0.05 | |
| Table 3: | The effect of seed tuber size and growth regulators on the vigour in different cultivars of potato |
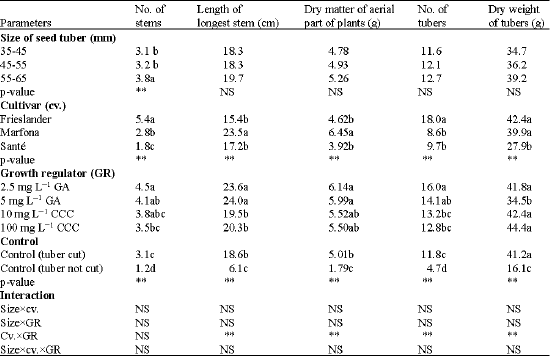 | |
| **Significant at p≤0.01, *Significant at p≤0.05. Values followed by a similar letter(s) are not statistically significantly different.DAP = Day After Planting, GA: Gibberellic Acid, CCC: Chlorocholine Chloride, cv. = Cultivar, GR: Growth Regulator, NS: Not Significant, p≤0.05 | |
The main effects of cultivar and growth regulator were both highly significant for the length of the longest stem (p < 0.0001). Marfona produced longer stems than Frieslander and Sante. The growth regulator effect showed that the seed tubers treated with GA (either 2.5 or 5 mg L-1) produced the longest stems.The uncut control produced the shortest stems.
Main effects of cultivar and growth regulator were significant for the dry weight of aerial parts (P < 0.0001; Table 3). The growth regulator effect showed that seed tubers treated with GA significantly produced more foliage than the controls.
The main effects of cultivar and growth regulator were significant for number of tubers (p < 0.0001) (Table 3). Frieslander produced the highest number of tubers, probably associated with the highest number of stems present in this cultivar. The main effect of growth regulator showed that seed tubers treated with GA 2.5 mg L-1 produced more tubers than seed tubers treated with CCC or not treated. The uncut control produced the lowest number of tubers, significantly fewer even than the cut control. There was a significant cultivarxgrowth regulator interaction (p < 0.0001) (Table 3).
The main effects on the dry weight of the tubers were significant for cultivar and for growth regulator (Table 3). The cultivar effect (p < 0.0001) showed that the cultivars Frieslander and Marfona produced higher tuber yields than the cultivar Sante. Seed tubers treated with GA 5 mg L-1 had significantly more yield than the uncut seed tubers, but a lower yield than the other treatments. The other treatments showed no significant differences among each other.
DISCUSSION
Length of the Longest Sprout
Seed tuber size had an effect on sprout elongation. The largest seed tuber size had longer sprouts than the other seed tuber sizes. The effect, though not large, is consistent with the generally observed effect of seed tuber size on vigour of each individual bud on the seed tuber (Struik and Wiersema, 1999). The size of the effect depends on the range of seed tuber sizes investigated. There was also a significant interaction between seed tuber size and cultivar for this parameter. Such interactions can occur when there are differences between cultivars in seed tuber shape or when the effect of size on the physiological behaviour (such as reflected by duration of dormancy or vigour, Van Ittersum, 1992a, b) is not the same for each cultivar.
Suttle (2004) showed that sprout length on seed tubers increases by exogenously applied GA. This was only investigated for the cultivar Russet Burbank. Our experiment confirms this. He also found that the endogenous GA levels became higher when the potato tuber reached dormancy release. After dormancy release the GA levels were still increasing. The seed tubers in this experiment were past their dormancy release point for a long time. This means that the endogenous GA levels were high. The different cultivars included in the experiment, however, may have differed in the temperature sum required to break dormancy and in the rate of physiological ageing after dormancy. For example, Marfona is known to have a long period of dormancy (Van Ittersum, 1992a, b).
The effect of GA on the length of the longest sprout was indeed different among cultivars. Frieslander did not react on GA in contrast to cultivar Marfona, which showed a strong response to both doses of GA by producing longer sprouts. Sante on the other hand only responded to the high dose of GA. These differences among cultivars could be explained by different sensitivities to GA. Since Frieslander is an early cultivar, which reached its dormancy release point sooner than the other cultivars, it is likely that the GA level in this tuber is so high that exogenous GA does not work anymore. Cultivar Marfona has a long dormancy and thus the endogenous levels of GA may have been much lower, which will enhance the effect of exogenous GA. Sante only responded to the highest concentration applied, suggesting that it has the right stage to be responsive but requires a higher dose as it is not sensitive to lower doses.
El-Sayed et al. (1988) investigated the effect of CCC on sprout length in potato. They observed that CCC reduced sprout length. This is consistent with the positive effect of GA on sprout length as we observed in the current experiment. However, it is not consistent with the lack of effect of CCC itself in our experiment. We made the observations on sprout length 11 days after seed tuber treatment, whereas El-Sayed et al. (1988) measured their sprouts after 4 months. It is possible that the CCC effect required more time than the 11 days given by us to come to expression.
Cutting stimulated sprout elongation for all seed tuber sizes. The effect of size of the seed tuber on the number of sprouts showed that larger seed tubers produce more sprouts than small seed tubers. This is caused by the increase in number of eyes in the seed tubers, which is higher with large seed tubers, but also by the amount of reserves available per eye (Struik and Wiersema, 1999).
Among the seed tuber treatments, the uncut seed tubers produced by far the lowest number of sprouts in all cultivars. Despite the energy required to heal the wounds caused by the cut, the cut seed tubers performed better, as the cutting procedure seemed to rejuvenate the seed tuber.
The weight of the sprouts depends upon the number of sprouts, the length of the sprouts and the thickness of the sprouts. The thickness of the sprouts was not measured. The results showed that the larger the seed tubers the higher the sprout weight, despite the fact that for length of the longest sprout and the number of sprouts only the difference between the largest seed tuber size and the two smaller seed tuber sizes were significantly different.
The effects of cultivar on sprout weight partly reflected the cultivar effects on length of longest sprout and partly the cultivar effect on number of sprouts.
Cutting had a strong effect on the weight of sprouts, which is consistent with the effects on length of the longest sprout and number of sprouts. GA in a low dose had a large negative effect on the sprout weight. During observations, it was obvious that GA treated seed tubers had thinner sprouts and this could explain a greater portion of the GA effect on weight. This effect was only visible in the lower concentration as for the higher concentration the lower sprout thickness was better compensated by longer sprouts and more sprouts per individual seed tuber. Different aspects of sprout behaviour show different sensitivities for GA.
In the results, the presented data includes not emerged seed tubers, for which the values for date of senescence were also set at 126 days. Since most not emerged seed tubers were with the uncut control, it seems logical that this treatment had the highest values for date of senescence. If the not emerged seed tubers were analysed as missing values, GA delayed senescence. Frieslander and Marfona showed no effects of size of the seed tubers on date of senescence, but Sante showed that the biggest seed tubers gave plants that senesced later, probably because of a more vigorous shoot, resulting in more sympodial layers.
CCC showed no retardant effect on the length of the longest stem. In wheat, CCC is used to retard stem elongation and Sharma et al. (1999) and Digby and Dyson (1973) showed that CCC applied to a potato crop in a foliar application did reduce shoot growth. This could mean that CCC applied to a seed tuber which is subsequently de-sprouted is not as effective as when sprayed on the foliage. The transport from the tuber to the new sprouts might not be efficient enough to establish a large effect.
Marfona produced the longest stems in all treatments. This should be a cultivar characteristic.
GA inhibits tuberization (Struik et al., 1999; Vreugdenhil and Sergeeva, 1999). In the experiment of Sharma et al. (1999), there were also fewer tubers per plant when treated with GA. However, in our experiments GA either had a positive effect on the number of tubers (Frieslander) or no effect at all (Marfona and Sante) compared to the cut control. In the cultivar Frieslander the positive effect was very large: many more tubers per plant were produced when the seed tubers had been treated with GA. The number of tubers almost doubled when treated with GA. Although GA might impede or delay tuberization, it can enhance the production of tuber sites, by increasing the number of stems, stimulating stolonization and enhancing stolon branching. Although probably all these effects occurred, the increased stolonization is most likely the dominant effect as appeared during the harvesting of the plants (Personal Communication).
CCC did not increase the number of tubers per plant compared to the cut control. This was true for all cultivars. This is not in line with the results of Radwan et al. (1971) and Sharma et al. (1999), who found that CCC increases the number of tubers. However, this was a foliar application and it differs with this experiment where the CCC was applied on seed tubers which were subsequently de-sprouted before planting .
CONCLUSIONS
The most striking findings are the following:
| • | Cutting of relatively old seed tubers seemed to improve their performance. |
| • | GA applied to seed tubers affected sprout behaviour and growth vigour after de-sprouting in various ways. The effects strongly depended on cultivar. In Frieslander, GA treated seed tubers produced more sprouts with GA, but of the same length. In Marfona GA treated seed tubers produced similar numbers but longer sprouts compared to the control. |
| • | CCC did not affect sprouting behaviour. |
| • | Although many significant effects on various parameters reflecting part of the growth vigour of seed tubers were recorded, none of the seed tuber treatments was able to increase the dry weight of the tubers, the ultimate test for the efficacy of a growth regulator. The only positive response relevant for a seed production scheme was observed in the cultivar Frieslander: it produced more tubers in response to GA, but this effect was mainly brought about by an increase in the number of stems per seed tuber. |
ACKNOWLEDGMENTS
My study in Wageningen University and Research Centre was supported by a grant from the Ministry of Jihad-e-Agriculture of Iran and the International Center for Agricultural Research in the Dry Areas (ICARDA). Hereby I would sincerely like to thank them for their encouragement, for giving me this opportunity and also for the funding of my study. Special thanks to Wageningen University and Research Centre and the Plant Sciences Group for their help, the management and the facilitating of my research project.
REFERENCES
- Bodlaender, K.B.A., H.M. Dekhuijzen, J. Marinus, A. Van-Es and K.J. Hartmans et al., 1985. Effect of physiological age on growth vigour of seed potatoes. A study with seed tubers of two cultivars stored at two different temperatures. Working Group Growth Vigour of Seed Potatoes. Wageningen, The Netherlands, pp: 142.
- Digby, J. and P.W. Dyson, 1973. A comparison of the effects of photoperiod and of a growth retardant (CCC) on the control of stem extension in potato. Potato Res., 16: 159-167.
CrossRef - Dyson, P.W., 1965. Effects of gibberellic acid and (2-chloroethyl)-trimethylammonium chloride on potato growth and development. J. Sci. Food Agric., 16: 542-549.
CrossRef - Hammes, P.S. and P.C. Nel, 1975. Control mechanisms in the tuberization process. Potato Res., 18: 262-272.
CrossRef - Lommen, W.M.J. and P.C. Struik, 1993. Performance of potato minitubers in a controlled environment after different storage periods. Potato Res., 36: 283-292.
CrossRef - Lommen, 1994. Effect of weight of potato minitubers on sprout growth, emergence and plant characteristics at emergence. Potato Res., 37: 315-322.
CrossRef - Menzel, C.M., 1980. Tuberization in potato at high temperatures: Responses to gibberellin and growth inhibitors. Ann. Bot., 46: 259-265.
Direct Link - Ranalli, P., 1997. Innovative propagation methods in seed tuber multiplication programmes. Potato Res., 40: 439-453.
CrossRef - Rex, B.L., 1992. Effect of two plant growth regulators on the yield and quality of Russet Burbank potatoes. Potato Res., 35: 227-233.
CrossRef - Sharma, N., N. Kaur and A.K. Gupta, 1999. Effects of gibberellic acid and chlorocholine chloride on tuberisation and growth of potato (Solanum tuberosum L.). J. Sci. Food Agric., 78: 466-470.
CrossRef - Struik, P.C., D. Vreugdenhil, H.J. Van-Eck, C.W. Bachem and R.G.F. Visser, 1999. Physiological and genetic control of tuber formation. Potato Res., 42: 313-331.
CrossRef - Struik, P.C., P.E.L. Van-der-Putten, D.O. Caldiz and K. Scholte, 2006. Response of stored potato seed tubers from contrasting cultivars to accumulated day-degrees. Crop Sci., 46: 1156-1168.
Direct Link - Suttle, J.C., 2004. Involvement of endogenous gibberellins in potato tuber dormancy and early sprout growth: A critical assessment. J. Plant Physiol., 161: 157-164.
Direct Link - Tognoni, F., A.H. Halevy and S.H. Wittwer, 1966. Growth of bean and tomato plants as affected by root absorbed growth substances and atmospheric carbon dioxide. Planta, 72: 43-52.
CrossRef - Van der Zaag, D.E. and C.D. van Loon, 1987. Effect of physiological age on growth vigour of seed potatoes of two cultivars. 5. Review of literature and integration of some experimental results. Potato Res., 30: 451-472.
CrossRefDirect Link - Van Ittersum, M.K., 1992. Variation in the duration of tuber dormancy within a seed potato lot. Potato Res., 35: 261-269.
CrossRef - Vreugdenhil, D. and L.I. Sergeeva, 1999. Gibberellins and tuberization in potato. Potato Res., 42: 471-481.
CrossRefDirect Link - Wiersema, S.G., 1989. Comparative performance of three small seed tuber sizes and standard size seed tubers planted at similar stem densities. Potato Res., 32: 81-89.
CrossRef